







The following is the established format for referencing this article:
Cockle, K. L., O. Villalba, N. Fariña, A. Bodrati, L. G. Pagano, E. D. Stein, and A. R. Norris. 2023. Non-breeding ecology of a Neotropical-Nearctic migrant, the Common Nighthawk (Chordeiles minor): habitat, activity patterns, molt, and migration. Journal of Field Ornithology 94(3):8.ABSTRACT
Long-distance migratory aerial insectivores are among the most threatened groups of birds breeding in North America, yet little is published about the two-thirds of their annual cycle that unfolds in South America. To study non-breeding ecology and migration of the Common Nighthawk (Chordeiles minor), we observed, captured, and GPS-tagged individuals at Reserva Natural Rincón de Santa María, an Important Bird Area on the Paraná River in Corrientes, Argentina, from 2017 to 2022 (237 captures, 95 individuals, 556 GPS fixes from 8 females). Common Nighthawks arrived at the reserve (late November to mid-January) partway through flight feather molt; primaries 8, 9, and 10 molted consecutively until early February, followed by body feathers. Three tagged females spent December/January to March at the reserve, May to August in Florida (USA), and October/November to December/January in the Cerrado (Brazil), and at least one traveled 1800 km while molting P8. Recapture rates across seasons were 44% for females and 25% for males. Small body size and breeding locations in Florida are consistent with the chapmani subspecies previously recorded from Argentina. Direct observations and GPS tracking (8 individuals) revealed that nighthawks roosted during the day in mature exotic pines; foraged over pines and native grasslands for 26–41 min within ~40 min after sunset, nearly always in silence; and rested for the remainder of the night in native grasslands, on roads, or on rocky platforms. Common Nighthawks should be included among the growing number of migratory species that exhibit long-distance movements between stationary sites during their long non-breeding period. Our results also suggest site fidelity to non-breeding areas, a continuous molt-migration strategy, and some nuanced migratory connectivity. Programs aiming to conserve long-distance migratory aerial insectivores should support long-term monitoring led from the Global South, especially in the Cerrado and southern South America. Please see Appendix 1 for a Spanish translation of this article.
RESUMEN
Los insectívoros aéreos migratorios de larga distancia se encuentran entre los grupos de aves más amenazados que se reproducen en Norteamérica, sin embargo, poco se ha publicado sobre los dos tercios de su ciclo anual que se desarrolla en Sudamérica. Para estudiar la ecología no reproductiva y la migración del Añapero Boreal (Chordeiles minor), observamos, capturamos y marcamos con GPS individuos en la Reserva Natural Rincón de Santa María, un Área Importante para la Conservación de las Aves sobre el río Paraná en Corrientes, Argentina, desde 2017 hasta 2022 (237 capturas, 95 individuos, 556 GPS fijos de 8 hembras). Los Chordeiles minor llegaron a la reserva (finales de noviembre a mediados de enero) a mitad de la muda de las plumas de vuelo; las primarias 8, 9 y 10 mudaron consecutivamente hasta principios de febrero, seguidas de las plumas del cuerpo. Tres hembras marcadas pasaron de diciembre/enero a marzo en la reserva, de mayo a agosto en Florida (EE.UU.), y de octubre/noviembre a diciembre/enero en el Cerrado (Brasil), y al menos una viajó 1800 km mientras mudaba P8. Las tasas de recaptura entre estaciones fueron del 44% para las hembras y del 25% para los machos. El pequeño tamaño corporal y las localidades de cría en Florida concuerdan con la subespecie chapmani registrada previamente en Argentina. Las observaciones directas y el seguimiento por GPS (8 individuos) revelaron que los C. minor se posaban durante el día en pinos exóticos maduros; buscaban alimento sobre pinos y pastizales nativos durante 26-41 min en un plazo de ~40 min tras la puesta de sol, casi siempre en silencio; y descansaban el resto de la noche en pastizales nativos, en carreteras o en plataformas rocosas. Chordeiles minor debería incluirse entre el creciente número de especies migratorias que muestran movimientos de larga distancia entre lugares estacionarios durante su largo periodo no reproductor. Nuestros resultados sugieren también una fidelidad a las zonas donde no se reproduce, una estrategia continua de muda-migración y cierta conectividad migratoria matizada. Los programas destinados a conservar los insectívoros aéreos migratorios de larga distancia deberían apoyar un seguimiento a largo plazo dirigido desde el Sur Global, especialmente en el Cerrado y el sur de Sudamérica.
INTRODUCTION
Long-distance Neotropical-Nearctic migratory aerial insectivores inhabit South America for about two-thirds of their annual cycle, returning each year to North America for a brief four-month breeding period. Many of these species are known to be declining, but a single driver is unlikely (Michel et al. 2016). Efforts to identify stressors are hindered by large gaps in knowledge about the ecology, distribution, and natural history of these birds while they are in South America (Nebel et al. 2010, Spiller and Dettmers 2019, Albert et al. 2020). Researchers have identified migratory routes and linked some non-breeding and breeding areas by tracking individuals from breeding populations in North America (e.g., Fraser et al. 2017, Imlay et al. 2018, Knight et al. 2021a). However, knowledge of non-breeding habitat use, behavior, molt, group size, and movement patterns remains patchy at best (e.g., Somenzari et al. 2018), and many assumptions about non-breeding ecology are based on extrapolation from the short portion of their annual cycle that these birds spend in North America.
Filling gaps in the basic non-breeding ecology and natural history of Neotropical-Nearctic migratory aerial insectivores is critical for moving forward with in-depth studies and conservation planning. To identify areas for conservation, understand carry-over effects, assess migratory connectivity, and build conservation partnerships, it is important to connect non-breeding areas with breeding areas and stopover sites (Somenzari et al. 2018, Spiller and Dettmers 2019, Albert et al. 2020). Surveys are needed to determine seasonal distribution, occupancy, and abundance during the long non-breeding season, but first researchers need a basic understanding of group dynamics and behavior, including information about the conditions under which birds can be detected, identified, and counted, particularly for species that are difficult to distinguish visually or are active at night. Researchers also need knowledge of where and when birds molt their feathers, not only to understand the energetic trade-offs involved in molt and migration, but also to link non-breeding and breeding areas via isotope analysis of feathers (e.g., García-Pérez and Hobson 2014). To identify critical habitat and develop conservation plans, managers need information about home range size and how birds use different habitats and sites (e.g., foraging, roosting; Knight et al. 2021c). Researchers could program satellite tags to study use of space and habitat selection by aerial insectivores, but first we need an understanding of their daily activity patterns. It is important to assess whether availability of high-quality prey has long-term effects on survival and reproduction (Spiller and Dettmers 2019), but to survey prey in non-breeding areas researchers first need information about the timing, habitats, and heights at which aerial insectivores forage. Here, we offer a first step and a model to fill gaps in knowledge of non-breeding ecology for one species, the Common Nighthawk (Chordeiles minor), a long-distance migratory aerial insectivore that has suffered steady declines in abundance over recent decades (Nebel et al. 2010, Environment Canada 2016).
At a broad scale, conserving Common Nighthawks requires an understanding of where these birds occur throughout the non-breeding season. Nighthawks spend the non-breeding season (i.e., September to April; austral spring, summer, and fall) in South America, breed during May–August across Central and North America (Brigham et al. 2020), and migrate south through the Darien (Colombia) in September (Bayly et al. 2014, Knight et al. 2021a). Their distribution, and the timing of their movements within South America from October to April, remain unclear. Based on observational records, from October to March they are generally assumed to occupy a non-breeding range in eastern Ecuador, eastern Peru, and southern South America (i.e., southern Brazil, Paraguay, Uruguay, and northern Argentina; Stotz et al. 1992, Bayly et al. 2014, Brigham et al. 2020, https://ebird.org, https://www.wikiaves.com.br). However, 41 of 43 Common Nighthawks satellite-tagged at 12 locations across the United States and Canada (excluding the southeastern United States) spent the non-breeding season in the northern Cerrado and southern Amazon of central Brazil (Ng et al. 2018, Knight et al. 2021a), approximately 2000 km north of the concentration of e-bird records in southern South America. The other two tracked individuals spent part of their non-breeding season in southern South America: one traveled from south Texas (United States) to the Chaco of western Paraguay, and the other traveled from New Brunswick (eastern Canada, subspecies minor) to southeastern Brazil.
To understand carry-over effects, differential population declines, and stressors during the non-breeding season, it is important to assess migratory connectivity: the degree to which separate breeding populations use geographically distinct non-breeding areas (Kramer et al. 2018). Results from global positioning system (GPS) tracking of Common Nighthawks contrast with direct observations. Nighthawks tagged across North America mixed indiscriminately during the non-breeding season (i.e., low migratory connectivity; Knight et al. 2021a). However, of the nine described subspecies, only C. m. chapmani and, to a lesser extent, C. m. minor have been collected in southern South America (Argentina, Paraguay, Uruguay, and southern Brazil; Hartert and Venturi 1909, Oberholser 1914, Steullet and Deautier 1935–1946, Zotta 1937, Brodkorb 1938, Cuello 1966, Belton 1978, Olrog 1979, Navas and Bó 1988). The minor subspecies breeds across much of Canada and the northeastern United States, and the chapmani subspecies breeds along the Gulf slope of the United States from eastern Texas to Florida, and, on the Atlantic slope, throughout the Florida peninsula north to South Carolina (Brigham et al. 2020). Subspecies identifications in South America were based on measurements and plumage patterns, which overlap among subspecies (Pyle 1997) and are not always reliable. However, they offer a potential clue as to why only two of the 43 GPS-tagged birds migrated to southern South America: tagging in breeding areas did not include the subspecies chapmani, neotropicalis, and panamensis (i.e., populations in the southern part of the breeding distribution, Florida to Central America). In summary, (1) there are many records of Common Nighthawks in southern South America, (2) most of the birds collected in southern South America were identified as C. m. chapmani (breeding in Florida or southeastern United States), and (3) birds GPS-tracked from Canada and the United States (except Florida) rarely traveled as far as southern South America. This evidence suggests that, contrary to the hypothesis of indiscriminate mixing during the non-breeding season, Chordeiles minor might exhibit a chain migration pattern, whereby more southerly breeding populations spend the non-breeding season farther south (e.g., as shown for another aerial insectivore, the Common Swift, Apus apus; Åkesson et al. 2020).
Among the major stressors thought to affect Common Nighthawks are habitat loss and insect declines, which probably affect aerial insectivores across their annual cycle (Nebel et al. 2010, Environment Canada 2016). Conservation actions to address these stressors in South America would first require an understanding of habitat and foraging requirements. Common Nighthawks appear to require open areas for hawking insects on the wing, and they may or may not be associated with water bodies on their non-breeding grounds (Environment Canada 2016, Brigham et al. 2020). In South America they occur in open areas, savannas, and scrublands; and over large rivers, second-growth forests, tree plantations, and salt flats (Costa et al. in press). They were observed roosting by day, alone or in groups, among the branches of Ficus sp. trees in brushy areas among sandy grassland; along limbs of scattered trees in city parks, campuses, and ranch headquarters; on horizontal branches of native trees (Tipuana tipu, Prosopis sp.) on a roadside in disturbed habitat along a small river; and in exotic trees (Eucalyptus spp., Populus spp., Salix spp.) of cities (Belton 1978, Ingels et al. 1999, Bodrati 2005, Salvador and Bodrati 2017). Lillo (1905, as cited in Steullet and Deautier 1935–1946) reported that they were very common in mountain forests in Tucumán (Argentina), in natural hedgerows, and on the margins of the Calera River. Five of the seven individuals tracked by Ng et al. (2018) to the Cerrado and Amazon had roosting home ranges in patchy landscapes composed of forest, shrubland, cropland, and grassland. The remaining two roosted in continuous forest. It is not known where they foraged or spent nights. To our knowledge, nothing has been published on Common Nighthawk sounds or daily activity in South America (Brigham et al. 2020).
To understand energy trade-offs during the non-breeding season and to develop studies using feather isotopes to link breeding and non-breeding areas, we need to know where and when migratory birds molt. Molt is energetically costly and does not usually overlap with migration (Newton 2009, Renfrew et al. 2011). However, current ideas about where and when Common Nighthawks molt are mostly based on assumptions and speculation. Like most or all Caprimulgidae (Johnson and Wolfe 2017), Common Nighthawks exhibit a “complex basic molt strategy.” Brigham et al. (2020) indicate that definitive prebasic molt occurs “on the non-breeding grounds,” suggesting a stationary area where birds molt, which we will challenge in this paper. Oberholser (1914) indicated that Common Nighthawks leave North America with flight feathers intact, molting their flight feathers distally (P1 to P10) in January and February. Brigham et al. (2020) indicated molt of flight feathers primarily from November to January, and body feathers primarily from September to December. However, Pyle (1997:105) noted that further study is needed to understand the timing of flight feather replacement, and thus the timing of age-code assignments.
Here, we contribute information on the non-breeding ecology, migration, and natural history of Common Nighthawks based on fieldwork during the austral summer from 2017 to 2022 in grasslands of northeastern Argentina. Specifically, we contribute sex-specific return rates; describe timing of molt of primaries and body feathers; link individuals to breeding areas in North America; describe the timing and location of migration routes, stopovers, and stationary non-breeding areas; and offer a preliminary analysis of habitat use, activity patterns, and daily movements. The hypothesis of low migratory connectivity in Common Nighthawks (indiscriminate mixing in non-breeding areas; Knight et al. 2021a) predicts that nighthawks from northeastern Argentina should spend the breeding season at a variety of sites across North and Central America. In contrast, the hypothesis of chain migration predicts that Common Nighthawks from Argentina will breed in the southeastern United States or Central America. Considering the widely-held idea that Common Nighthawks prefer to forage over open water, we predicted that we would often detect them in flight over a large reservoir at our study area in northeastern Argentina. If Common Nighthawks molt flight feathers while stationary at a non-breeding area (as suggested by Oberholser [1914] and Brigham et al. [2020] and assumed for most Neotropical-Nearctic migrants, because of the energetic cost of molt), they should arrive in northeastern Argentina with a complete set of worn flight feathers, molt sequentially over 2–3 months, and leave for their breeding areas with a full set of new feathers. Our aim is not to explain why Common Nighthawks or other aerial insectivores are declining, but to fill critical gaps that currently prevent in-depth studies and conservation of these birds during the two-thirds of their lives spent in South America.
METHODS
Study area
We studied Common Nighthawks from January 2017 to March 2022 in grasslands of northeastern Argentina (northern Corrientes and southern Misiones provinces), primarily at the Reserva Natural Rincón de Santa María (RNRSM) Important Bird Area, Ituzaingó Department, Corrientes (27°31’ S, 56°36’ W; elevation 90 m; Krauczuk and Di Giacomo 2005). RNRSM is situated in the transition zone between the Campos District and the Atlantic Forest, with influences from the humid Chaco, Esteros del Iberá, and Delta e Islas del Paraná eco-regions (Cabrera 1971, Burkart et al. 1999). To the north, it adjoins the 160,000-ha reservoir created by the Yacyretá hydroelectric dam on the Paraná River. To the east, south, and west, it is surrounded by private properties with cattle pastures and tree plantations.
RNRSM is characterized by humid grassland with Andropogon lateralis and A. virgatus (2474 ha), dry grasslands of Elionurus muticus (274 ha), flooded grasslands of Andropogon lateralis (42 ha), native secondary forest (5 ha), and abandoned plantations of exotic pines and Eucalyptus, as well as areas invaded by these species (571 ha; Montiel et al. 2016). The soil has low permeability and slow drainage, such that the grasslands remain flooded by rain much of the time; however, there are three drier areas in the north, south, and west with slightly higher ground (100 m asl), where the soil is more permeable and drainage is faster (Hansen 1998). The grasslands are peppered with shrubs and vines such as guayaba chica (Psidium guineense, Myrtaceae), yuquerí (Mimosa pigra, Fabaceae), and ramilletes de campo (Eupatorium spp., Asteraceae). The gallery forest includes early successional species such as ambay (Cecropia pachystachya, Cecropiaceae), burro caá (Casearia sylvestris, Salicaceae), and fumo bravo (Solanum granulosum-leprosum, Solanaceae). There have been 21 fires in the RNRSM since 1994, with an average size of 466 ha. The largest fires occurred on 9 November 2000 (1500 ha burned), 22 January 2013 (1500 ha burned), and 24 October 2019 (1811 ha burned). The RNRSM also includes part of a paved dyke (7.4 km); artificial rocky platforms constructed to contain the reservoir; a mown-grass landing strip for small planes; and ~15 km of lightly-used, partly overgrown dirt roads. Several canals remain from rice plantations and dam construction prior to the creation of the reserve in 1994. Pine plantations, within and adjacent to the reserve, produce seeds that invade the native grasslands. To control the invasions, contractors remove pine saplings and some stands of larger trees, leaving behind the cut trees and stumps. Reserve staff enrich the secondary forests by planting native trees to assist natural regeneration.
We include some nighthawk captures and other incidental field observations (1998–2022) from Ituzaingó, Corrientes (27°35’ S, 56°41’ W); Reserva Mbaracayá, contiguous with RNRSM (Ituzaingó, Corrientes, Argentina; 27°29’ S, 56°42’ W); Itatí (Corrientes, Argentina; 27°16’ S, 58°14’ W); Reserva Natural Provincial de Iberá (Corrientes, Argentina; 27°39’ S, 57°02’ W); pasture adjacent to Reserva Urutaú, a peri-urban area near Candelaria, Misiones, Argentina (27°30’ S, 55°48’ W); Parque Nacional Chaco, Chaco, Argentina (26°48’ S, 59°36’ W); Laguna Corá, Chaco, Argentina (27°24’ S, 58°50’ W); Isla del Cerrito, Chaco, Argentina (27°17’ S, 58°37’ W); Vuelta de Obligado (Buenos Aires, Argentina; 33°36’ S, 59°49’ W); and Arroyo Tagatiyamí (Concepción, Paraguay; 22°40’ S, 57°33’ W).
Field methods
We searched for nighthawks and nightjars year-round using vehicles, reflectors, and flashlights, for a total of 530 nights, 518 of which were at RNRSM, where we searched on dirt roads, the paved dyke, artificial rocky platforms, and the landing strip (see details in Bodrati et al. 2019). We captured birds by spotlighting and using a dip net, a technique that we piloted in February 2016 after several years of experience capturing other species of nightjars by hand (without a net). A dip net is appropriate for Common Nighthawks in our study area because they rest on open ground, startle easily, and flush straight upward into the net.
We took measurements of each nighthawk (not reported here) and banded it using a numbered aluminum band. We determined age and sex by plumage (Selander 1954, Pyle 1997, Brigham et al. 2020). We examined molt of primaries (right wing) and body feathers. All nighthawks were released at the site of capture. To determine migratory routes and breeding sites, we attached Pinpoint-10 GPS loggers (Lotek) to 4 females in January 2019 and 3 females and 6 males in January and February 2020. We programmed loggers to take “swift” fixes throughout the year (every 7–10 days from January to mid-March, every 4–10 days from mid-March to mid-May, every 10–15 days from mid-May to mid-September, and every 5–10 days from mid-September to late January, while the tag battery lasted). The loggers stored GPS points and required recapture to download. To study use of habitat within the non-breeding site, we programmed another set of loggers to take frequent fixes over the course of 2 to 5 nights and affixed these loggers to an additional 5 females and 1 male. We updated the timing of attempted fixes as we learned about movements from each logger we retrieved. We attached loggers to birds using a backpack-style harness made with elastic cord (Stretch Magic) and crimp tubes. Birds were processed, from capture to release, in 12 minutes or less. We interpreted GPS fixes when the logger reached 5 or more satellites. Note that we did not recapture any males with GPS loggers, so we provide tracking results on females only.
We made opportunistic observations and recordings of Common Nighthawk foraging, roosting, vocalizations, and mechanical sounds at RNRSM and other sites in southern South America. To better understand the seasonal and daily timing of nighthawk vocalizations and mechanical sounds in South America, we searched for audio records using the online platforms xeno-canto (https://xeno-canto.org), Macaulay Library of Natural Sounds (https://www.macaulaylibrary.org), and WikiAves (https://www.wikiaves.com.br). For each recording, we read the notes on timing and behavior, and contacted authors for clarification when necessary.
Unless otherwise indicated we present results in terms of mean ± standard deviation and refer to non-breeding seasons in Argentina (November–March) by the new year in January (e.g., November 2020–March 2021 would be the 2021 season). We used https://www.timeanddate.com to determine sunrise and sunset times.
RESULTS
Seasonality and return rates at RNRSM
We made a total of 237 captures of 95 individuals (46 females, 44 males, 5 unknown sex) over 6 non-breeding seasons at RNRSM. We captured an additional 2 females and 1 male at Reserva Urutaú (January and February 2020) and 1 female and 1 male at Reserva Mbaracayá (January 2020). Common Nighthawks arrived at RNRSM from late November to January (see Migration and breeding grounds, below). For each season, our earliest captures of Common Nighthawks occurred on 19 November 2018, 3 December 2019, 4 December 2020, and 1 December 2021. Our latest captures were 15 March 2019, 13 March 2020, and 13 March 2022. Peak capture rates occurred in late December (females) and January (both sexes; Fig. 1). Both sexes were present from the first week of December to mid-March.
Within-season recaptures were common. If we captured an individual at the very beginning of the season (i.e., before 8 December) or the very end of the season (i.e., March), we usually also captured it mid-season (17 of 20 individuals). Of the birds we captured in December at RNRSM, 34% were recaptured in January to March of the same non-breeding season, suggesting that Common Nighthawks remain at RNRSM from arrival until northern migration.
Overall, 31 ± 14% (range: 19%–54%, n = 5 years) of birds marked in a given non-breeding season were recaptured the following non-breeding season (Table A2.1). After two seasons, we recaptured 33 ± 11% (range: 20%–46%, n = 4). After three seasons, we recaptured 22 ± 13% (range: 7%–29%, n = 3), and after four seasons, 15 ± 1% (range: 14%–15%, n = 2). However, return rates differed by sex: we were more likely to recapture females (44%) than males (25%) in the following non-breeding season (chi-squared test, chi² = 4.26, P = 0.039). One individual (a male) marked in the 2017 season was recaptured as late as the 2022 season.
Migration and breeding grounds
We retrieved GPS loggers with migration data from three out of nine of the originally tagged individuals, all adult females (a total of 105 good fixes; Fariña et al. 2023). We tagged these individuals, identified by their band numbers, at RNRSM in January 2019 (RSM140 and RSM198) and January 2020 (RSM152). We recovered the loggers at RNRSM in December 2019, January 2020, and January 2021, respectively. All three individuals remained at RNRSM throughout February and the first half of March. They left the reserve in the second half of March, spent the breeding season (May to August) in Florida, United States, and spent ~3 months in the Brazilian Cerrado before returning to RNRSM in late December or January (Fig. 2, Figs. A2.1–A2.3). When traveling northward, they took a fairly direct route through the lowlands of Bolivia (20 and 24 March), the western Amazon (western Brazil and northeastern Peru; 25–30 March), and central Colombia. At least two (and possibly all three) of the three individuals stopped for 2 to 3 weeks in the Orinoco region of Colombia during the month of April (RSM198: at least 1–21 April 2019; RSM152: at least 4–19 April 2020). These individuals occupied sites that were 42 km from one another along the same dirt road. They crossed the Caribbean either from northern Colombia/northern Venezuela (RSM140 and RSM198) or off the east coast of central America (RSM152). They spent the breeding season in Florida at Citrus Wildlife Management Area (RSM140), in the city of Naples (RSM198), and in the Marjorie Harris Carr Cross Florida Greenway (RSM152). They left Florida in early September and traveled south through Colombia and the western Amazon of Brazil (September), then eastward across the southern Amazon/northern Cerrado of Brazil (early October), where they stopped for up to 3 months. RSM140 stayed at least from 14 November until 14 December (when the tag battery died) at Brasnorte, Mato Grosso state and was recaptured on 22 December at RNRSM. RSM198 stayed from 16 October until 8 January (when the tag battery died) at Ilha do Bananal, Tocantins state, and was recaptured on 31 January at RNRSM. RSM152 stayed at least from 8 to 22 October (when the tag battery died) at Río Teles Pires, Mato Grosso, and was recaptured on 15 January at RNRSM.
Molt
Nearly all adults arrived in Corrientes part way through their flight feather molt (Fig. 3). We recaptured RSM140 at RNRSM 8 days after its last GPS fix in the Cerrado, with P1-8 new, P9 80% grown, and P10 old, indicating that the new P9 was almost certainly growing during the 1800-km journey south from Brasnorte to RNRSM. Of our 84 captures from November to 14 January, 79 birds (94%) had at least one flight feather (primary, secondary, or retrix) missing or growing. Fifty-nine of these birds had at least one primary missing or growing, and 20 of the remaining 25 had at least one secondary or tail feather missing or growing; eight of these were classed as juveniles (i.e., age < 1 year; primaries not molted). Two birds did not have any molt in flight feathers, and three were missing data on molt (possibly because they were not molting). We have molt data for only one individual captured in November (19 November): primaries P1-6 were new and fully grown, P7 was 60% grown, and P8-10 were old. Birds captured in the first week of December typically had new primaries 1-7, often with P8 growing or complete, and P9-10 old (Fig. 3). Thereafter they molted P9 (peak in the third week of December) and then P10 (peak in mid-January; Fig. 3). Typically, in any given capture, if one primary was missing or < 10% grown, the previous primary was 80%–100% grown, and the remaining primaries were fully grown. From mid-January through mid-March, all adults captured were molting body feathers (n = 53).
Daytime social roosting
We observed Common Nighthawks roosting during the day, always on horizontal branches, and often in groups, although it was not possible to confirm the total number of individuals unless we visited the roost at dusk (Table A2.2). At RNRSM, at least four GPS-tagged individuals shared a single roost area. On 4 March 2021 we found one Common Nighthawk sleeping there, about 20 m above ground, on a horizontal branch of a Pinus elliottii. It flew up from the roost at 19:15, 1 minute before sunset. The following day, 5 March, we returned to the roost site, and a Common Nighthawk, presumably the same individual, was sleeping on the same branch, in the same position as the previous day. This time it left the roost at 19:18, 5 minutes after sunset. At least seven other individuals flew up from trees within a 50 m radius around the same time. On 8 March the nighthawk was again sleeping in the same position, and left the roost at 19:24, 14 minutes after sunset, accompanied by at least three other individuals. All of these nighthawks flew up from the pines suddenly, traveling southward (away from the reservoir) above the canopy of the mature pines.
Daily activity patterns, foraging, and sounds
We observed nighthawks foraging at a height as low as 2 m and displaying at a height of ~50 m, nearly always around dusk. At a point 700 m south of the shared roost site, on 13 days in March 2021 and February 2022, OV, NF, AB and ES recorded flight displays, foraging, vocalizations, and mechanical sounds. On leaving their roost, several individuals congregated and made circular flights in a radius of about 100 m, above the pine and eucalyptus plantations, where they foraged and occasionally made sounds for a few minutes, always before dark. Every time we observed them at this foraging location, birds appeared between 2 minutes before and 16 minutes after sunset, and they dispersed between 16 and 22 minutes after sunset, always before dark. On one of the evenings, 19 March 2021, five individuals foraged within a large swarm of dragonflies (Odonata). Upon departing, birds appeared to mostly fly off in the same direction, often toward the south (away from the reservoir), but it is unclear whether they resumed foraging at another location or ceased their evening activity. Additionally, on 29 January 2021 at 20:16 h (34 minutes after sunset) we observed an individual foraging approximately 8 m above the grassland beside the dyke at RNRSM. On 12 January 2023 at 20:10 h (24 minutes after sunset), during the strong winds and lightning leading up to an electrical storm, we observed and filmed at least six individuals foraging less than 1 m above the grassland and overgrown grassy roads at RNRSM.
We have only observed nighthawks foraging around dawn on two occasions. OV and NF observed one nighthawk foraging low over the grassland at RNRSM at 6:10 h (26 minutes before sunrise) on 25 February 2022, at the location where we previously observed groups foraging and vocalizing, 700 m south of the shared roost. AB observed 2 Common Nighthawks foraging at a height of 15–20 m over the Arroyo de los Cueros, a natural creek surrounded by espinal (thorny deciduous shrubland) and talar de barranca (dry forest on riverbank) around sunrise at Vuelta de Obligado (Buenos Aires, Argentina), in December 2000. During six dawn searches (2:30–6:30 AM) in December 2020 and January 2021, we did not observe Common Nighthawks foraging.
We heard vocalizations (calls in flight) or mechanical sounds (wing boom in flight) between 6 and 20 minutes after sunset on 9, 10, 11, and 12 March 2021 (Table A2.3; https://xeno-canto.org/728651). Birds emitted wing booms when descending rapidly in a U shape from an altitude of 40–50 m (see Miller 1925). Additionally, OV and NF heard a Common Nighthawk vocalizing in flight above the city of Ituzaingó on 23 March 2022, at dusk. Other than alarm calls of captured birds, we have not heard Common Nighthawk sounds at night in South America.
In the sound collections at https://xeno-canto.org, https://www.macaulaylibrary.org, and https://www.wikiaves.com.br (reviewed 5 February 2023), there are only 13 recordings of Common Nighthawks from South America (Table A2.3). They include “peent” calls, clucking, and wing booms. Wing booms (i.e., displaying males), however, are restricted to the beginning (September) and end (March, April) of the non-breeding period in South America. Recordists indicated time of day on nine of these recordings. Eight occurred between 16 minutes before and 75 minutes after sunset, and one was 60 minutes before sunrise (Table A2.3). Referring to his recording of Common Nighthawks at Praia do Laranjal (2016, XC308563/WA2005814), F. Jacobs (2023, personal communication) reported that the small group of birds vocalized for a short time, before sunset, while there was still considerable light, and apparently just after they left a roost. They flew at medium height initially, then gained height with somewhat irregular circling movements, and after reaching a certain height they disappeared in a southerly direction. They repeated the behavior for a few days, and then they were gone. Jacobs noted that he has never heard Common Nighthawks vocalize at any other time of day, neither at dawn nor at night.
We obtained a total of 483 GPS fixes at RNRSM, from eight tagged individuals (Fig. 4). They included 22 fixes from the three females tracked over the full annual cycle, and 461 additional fixes from five adult females tracked over the short term. We obtained 50 fixes during the day when birds were presumably roosting (five individuals over 13 days). We obtained 69 fixes in the 45 minutes after sunset (dusk; three individuals over seven days), 357 at night (eight individuals over 41 nights), and seven in the 45 minutes before sunrise (dawn; two individuals over eight days). We have tracking data for two birds at sunset on one day each; they were still in their roost at sunset, and were flying (i.e., fixes were in different locations, birds apparently foraging) five and 15 minutes after sunset, respectively. We last recorded birds in flight (moving) 38 ± 5 minutes (range: 32–45 min) after sunset, and we first recorded them resting 43 ± 6 minutes (range: 34–50 min) after sunset (n = 3 individuals on seven nights). Thereafter, throughout the night, 350 of the 357 points for the five individuals tracked frequently (every two to 60 minutes across a total of 18 nights) fell within 10 m (i.e., the error of the GPS tags) of one of seven locations, or along a 230-m stretch of road where they were apparently resting. Six points fell within 15 m of one of these locations, and the remaining point was 85 m from one of these locations (Fig. 4C).
Use of habitat in Argentina
We captured Common Nighthawks at RNRSM at night on the rocky platforms (filled areas) beside the dyke, on the dyke itself, on some dirt roads, and on the landing strip. We rarely saw a Common Nighthawk on the main gravel road that passes the RNRSM, although other nightjars (Hydropsalis torquata, Setopagis parvula, Nyctidromus albicollis, and to a lesser extent Eleothreptus anomalus) frequently rest, at night, on this road, where they are sometimes killed by vehicles. We also did not find Common Nighthawks resting on roads completely overgrown with grass. Instead, they were usually found in areas partly grown over with sparse grass, especially on rocky soil with bare patches. They began to rest at night in the grassland north of the dyke after it was burned on 24 October 2019. They were common on the landing strip in 2019 (12 captures) and 2020 (16 captures), when the grass was sparse, but rare there in 2021 (four captures) and 2022 (one capture) when the grass completely covered the ground.
The Common Nighthawks we tracked by GPS tag at RNRSM generally stayed over land. They traveled within RNRSM and to neighboring properties, 2–7.4 km from their roost sites (Fig. 4). All daytime roosts located by GPS tag (six individuals) were in mature/abandoned pines near the edge of the Yacyretá Reservoir (Fig. 4A). During the night, the eight tagged birds appeared to be resting on the ground or on a log or stump, always within native grassland, on a little-used road through grassland, or on rocks adjacent to grassland (357 fixes, Fig. 4C). The two points where RSM1151 rested at night in February 2022 were both within grassland that had burned 28 days previously. In December 2021, RSM401 rested for three consecutive nights at two locations within 44 m of one another, in a grassland that was restored by clearcutting adult pines in 2018 (leaving branches and stumps). When birds appeared to be foraging on the wing (locations changing, within 45 minutes after sunset; three individuals) they were located over native grassland (28 fixes), tree plantations (21 fixes), or pine-invaded grassland (eight fixes; Fig. 4B).
Habitat use in other non-breeding areas
We obtained eight nighttime and two daytime fixes from two GPS-tagged individuals in the Orinoco region of Colombia, during their April stopover. Whereas the two daytime fixes were in gallery forest, the eight nighttime fixes were in open areas, apparently grassland, near a dirt road/track, with one fix on the road itself (Fig. A2.2). We obtained 20 nighttime and two daytime fixes from our GPS-tagged individuals at their first non-breeding site in the Cerrado (Fig. A2.3). All but two nighttime fixes were in or over what appeared to be open scrub. The remaining two nighttime fixes were in or over cultivated fields beside gallery forest, and the two daytime fixes were in gallery forest.
DISCUSSION
Our study revealed several interesting patterns in Common Nighthawk non-breeding ecology, some of them quite unexpected. The hypothesis of low migratory connectivity in Common Nighthawks (indiscriminate mixing in non-breeding areas; Knight et al. 2021a) predicted that nighthawks from northeastern Argentina should spend the breeding season at a variety of sites across North and Central America. Instead, we found that all three GPS-tagged birds spent the breeding season in Florida, United States, a finding that aligns with the chapmani subspecies evidence from museum specimens and the possibility of chain migration. Contrary to the hypothesis that Common Nighthawks forage over large bodies of water (Brigham et al. 2020), we did not detect any individuals foraging over the large artificial lake at RNRSM. Instead, they foraged over grasslands and exotic tree plantations. Also contrary to expectations based on published literature (Oberholser 1914, Brigham et al. 2020), our results show that flight feather molt in Common Nighthawks extends across at least two non-breeding areas, and likely also occurs during migration. Unexpectedly, we also found higher return rates for females than males. There are still important gaps in our understanding of Common Nighthawk non-breeding ecology, but our results suggest some important conclusions and new hypotheses that will support future research and conservation of aerial insectivores in South America.
Seasonality, migration, and implications for migratory connectivity
The consistent December-January arrival of Common Nighthawks at RNRSM and the GPS evidence from the three individuals that we tracked across the annual cycle suggest that Common Nighthawks only arrive in Corrientes after first spending several weeks or months at a stationary non-breeding site in the northern Cerrado. Likewise, 40 of the 43 individuals tagged by Ng et al. (2018) and Knight et al. (2021a) across North America had a stationary non-breeding site in the northern Cerrado or southern Amazon, and at least seven occupied two stationary non-breeding sites within South America. The Cerrado is a tropical savanna ecosystem that has received limited research and conservation attention compared to other ecoregions in South America, but it plays an important year-round role supporting non-breeding populations of migratory birds. For example, beyond its use by Common Nighthawks from October to December (or March), the Cerrado is occupied in November to January by Purple Martins (Progne subis) that breed throughout North America; in January to February by Veeries (Catharus fuscescens) that breed in Delaware, United States, and British Columbia, Canada; and in May to October by White-crested Elaenias (Elaenia albiceps) that breed in Patagonian forest of Argentina (Dornas and Pinheiro 2011, Bravo et al. 2017, Heckscher et al. 2021).
The breeding location of the three GPS-tracked individuals is consistent with the chapmani subspecies that breeds in the southeastern United States (Coues 1888), matching the evidence from museum specimens in southern South America (Oberholser 1914, Navas and Bó 1988). The coloration and measurements of individuals at RNRSM are also broadly consistent with the chapmani subspecies (wing: 187 ± 7 mm, range = 172–196; tail: 103 ± 4 mm, range: 94–110; mass = 72 ± 7 g, range = 60–85; n = 31 captures of individuals in February-March, not molting primaries or rectrices; Proyecto Atajacaminos, unpublished data). However, some of these measurements also overlap with those of C. m. neotropicalis, C. m. panamensis, C. m. aserriensis, and C. gundlachii, and measurements varied year-to-year in the same individuals. The migration routes of our tagged birds and their timing through northern South America and the Caribbean are broadly consistent with the results from birds tagged across Canada and the United States (Knight et al. 2021a). What differs is that we reliably detected only C. m. chapmani (breeding in Florida) at our field site, whereas the birds tagged across North America (except Florida) by Knight et al. (2021a) showed low migratory connectivity, apparently mixing indiscriminately in their stationary non-breeding areas, mostly in the Cerrado and Amazon. GPS-tagging nighthawks at other sites in Argentina and in the Cerrado, as well as those breeding in Florida, Mexico, Central America, and the Caribbean (including C. gundlachii), would vastly improve our understanding of their migratory connectivity. Foremost, to test the hypothesis of chain migration, it is important to identify the non-breeding areas of the populations that breed in Central America and determine whether all birds breeding in Florida, or just a subset, spend January to March in southern South America.
The Common Nighthawks at RNRSM exhibited annual return rates of 44% for females and 25% for males, with some birds returning to the same non-breeding area during at least 5 years. Common Nighthawks also exhibit relatively high fidelity to breeding sites. Ng et al. (2018) were able to track seven males from the boreal forest of Alberta using GPS tags, and all seven returned to within 1.3 km of their breeding site the following year. In the same region, Knight et al. (2022) found evidence for high male fidelity to specific nest sites, with 10 males showing significant overlap of wing boom areas across two years. Likewise, one female in Ohio, United States, returned to nest on the same rooftop in six separate years (Dexter 1961). In contrast, breeding Common Nighthawks exhibited very low fidelity to a highly-used foraging site, where hundreds of individuals gather along a short stretch of river in British Columbia, Canada, every night from June to late August (zero recaptures and zero detections with radio-transmitters, despite marking 10% of the population; McGuire et al. 2021). At RNRSM, we captured as many individual males as females, but we were much more likely to recapture the females, both within and between years. Males may exhibit larger non-breeding home ranges, lower site fidelity, or lower survival, or they may simply be more skittish and difficult to catch. Our high recapture rates for females meant that we were able to contribute information about the movements and habitat use of females, previously the under-represented sex in the literature on Common Nighthawks in South America (e.g., Ng et al. 2018, Knight et al. 2021a). Further studies could investigate sex differences in site fidelity, habitat use, foraging behavior, and survival in non-breeding areas.
Activity patterns and habitat use
Our results from observations and GPS tags suggest that Common Nighthawks at RNRSM left their roost between 1 minute before and 14 minutes after sunset, and they remained in flight until ~40 minutes after sunset, giving a 26–41 minute foraging period, which is shorter than foraging periods observed from June to August in North America. At Coeur d’Alene, Idaho, United States (48° N), Rust (1947) recorded the greatest numbers of nighthawks in flight over evening periods that lasted 75 minutes in June, 80 minutes in July, and 100 minutes in August. At Okanagan Falls, British Columbia, Canada (49° N), the average time between the first and last foraging nighthawks was 88 minutes (Brigham 1990). Near Winnipeg, Manitoba, Canada (50° N), nighthawks were typically observed foraging from 15 minutes before sunset until 40 minutes after sunset, for a total of 55 minutes (Taylor 2009). Brigham (1990) and Aldridge and Brigham (1991) suggested that the need for ambient light restricts foraging of Common Nighthawks to this short window before nightfall, and they might meet their energy needs quickly by capturing more than one prey item per attack. Shorter foraging periods at RNRSM could reflect the shorter twilight period closer to the equator. For example, nautical twilight, when the sun is 12° below the horizon, occurred 55 minutes after sunset on 31 January 2023 at RNRSM (28° S), but 88 minutes after sunset on 31 July 2022 at Okanagan Falls. Short foraging periods also suggest a high abundance of prey that allows Common Nighthawks to meet their energy requirements in a short time. The timing of foraging at RNRSM indicates that studies of their insect prey at subtropical south latitudes should focus within 40 minutes of sunset, and that GPS points taken outside of this period likely correspond to resting sites on the ground (nighttime) or in trees (daytime), not foraging. Moreover, although GPS tags, and possibly radio telemetry, could be used to study local movements and home range, points taken more than 40 min after sunset are likely to vastly under-represent both movements and home range size, given that birds rest for hours in the same location, night after night.
At no time did we detect Common Nighthawks at RNRSM foraging directly over the large body of open water that was adjacent to their roost sites. In contrast, in southern British Columbia (Canada), Common Nighthawks regularly travel up to 12 km from their roost sites to feed on the abundant insects over the Okanagan River at sx̌ʷəx̌ʷnitkʷ Provincial Park (McGuire et al. 2021). Although we did not record birds foraging directly over the reservoir at RNRSM, the whole region comprises a mosaic of temporarily flooded grasslands, rivers, lakes, and ponds, with little elevational relief, such that it is not clear whether insects captured by nighthawks over the grasslands and pines are terrestrial or aquatic. Based on their finding that terrestrial insects (especially beetles) comprised 65% of prey items in four nestling diet samples, Knight et al. (2018) suggested that Common Nighthawks in the boreal forest of Canada may be less dependent on aquatic habitats than previously thought. Upcoming genetic analyses of fecal samples will shed light on the nature of nighthawk diets at RNRSM and other sites, and help identify the sources of insect prey across the annual cycle, as a first step toward understanding how the fate of Common Nighthawks might be linked to that of their prey populations in South America.
Beyond their brief ~40 min period of foraging at RNRSM, Common Nighthawks roosted all day in trees, and rested all night in native grasslands or on the dyke. Our result that they rested, at night, exclusively in open areas suggests that widespread afforestation of grasslands with exotic pines, as currently practiced across much of Corrientes and southeastern Brazil, could threaten non-breeding habitat for nighthawks, as already indicated for other species of resident and migratory grassland birds (Azpiroz et al. 2012). Removal of invasive pines from grasslands (as underway at RNRSM) would increase nocturnal resting habitat and, we hypothesize, would also increase prey availability, if, as we expect, flooded native grasslands are more productive than exotic pines as habitat for high-quality insect prey. Although nighthawks used the abundant pines at RNRSM for daytime roosting, this use is probably opportunistic: they also roosted in gallery forest in the Orinoco and the Cerrado, and in a wide variety of native tree species across northeastern Argentina.
Filling research and conservation gaps for long-distance migratory aerial insectivores in South America
Our study, focused on the Common Nighthawks that converge at a single protected area in December to March, helps to fill some knowledge gaps from the annual cycle of this long-distance migratory aerial insectivore. To confirm or refute our main findings with respect to site fidelity, daily activity patterns, habitat use, and timing of molt, researchers will need to conduct fieldwork at other sites in South America. In particular, we strongly recommend field-based studies in the northern Cerrado, where it appears most Common Nighthawks and many other aerial insectivores spend at least part of their annual cycle.
Researchers have successfully employed auditory surveys, including listening points and automated recording units (ARUs), to assess habitat selection, population trajectories, and distributions of Common Nighthawks in North America (Knight et al. 2021b,c). Given the paucity of vocalizations and mechanical sounds we detected at RNRSM and in recordings from across South America, auditory surveys would likely be of limited use in the non-breeding season. Cohn-Haft (2012, XC98994; 2023 personal communication) remarked that his recording was the first and only time he has heard Common Nighthawks in South America, in 35 years living in the Amazon. This coincides with the experience of AB, who has lived within the range of Common Nighthawks in central and northern Argentina since childhood (~43 years observing birds) and heard Common Nighthawks only one time, at the shared roost at RNRSM. Unfortunately, the silence of Common Nighthawks in South America means they can easily be confused with other nighthawks, especially the closely-related Antillean Nighthawk (Chordeiles gundlachii), which is virtually indistinguishable in flight, and has a little-known but apparently overlapping non-breeding range (Guzy 2020, Perlut and Levesque 2020, Verón 2021, Stiles et al. 2022). Given the social roosting behavior of Common Nighthawks in South America, their return to previously-used roost sites, and the timing of vocalizations around dusk, we suggest searching for dormitories in trees, particularly at points used by GPS-tagged birds (e.g., in the Cerrado), and counting individuals as they emerge to forage.
Taken together with the data from Knight et al. (2021a), our finding of two widely separated stationary non-breeding areas in South America contributes to the growing understanding of the complexity of avian migration ecology. Widely separated non-breeding areas, or nomadic-like movements, have now been observed in several species of migratory birds, including species that breed in South America, and species, like Common Nighthawk, which breed in North America (e.g., Heckscher et al. 2011, Renfrew et al. 2013, Bravo et al. 2017, Albert et al. 2020). Nevertheless, the concept of “the South American wintering grounds” is still widely employed in research, conservation, and bird-watching circles, even for birds like Common Nighthawks that do not experience winter at any point in their lives. Rejecting this oversimplification (and misnomer) is an important first step toward developing conservation initiatives that fully encompass the complex movements and physiological stages of migratory birds across their annual cycle.
Studying another long-distance Neotropical-Nearctic migrant, the Bobolink (Dolichonyx oryzivorous), which eats seeds and insects, Renfrew et al. (2013) found a close link between movements within South America, and net primary productivity (measured by the Normalized Difference Vegetation Index; NDVI). Bobolinks left Venezuela as local primary productivity declined, moving southward to Bolivia and Argentina as primary productivity increased there. Referring to aerial insectivores that breed in North America, these authors noted that although several species, including Common Nighthawk, “migrate south of the equator to take advantage of flushes of insects,” movements of these species “do not appear to include long stops or timing that would match peak NDVI values.” Our study shows that at least one population of Common Nighthawks does undertake long stops, with movements between two widely distant stationary areas in South America. Their timing in Corrientes (December to March) roughly coincides with austral greening and a period of high net primary productivity in the grasslands (Vassallo et al. 2013). However, net primary productivity also remains high in the Cerrado throughout the spring and summer rainy season from November to April (Ferreira and Huete 2004), suggesting that, unlike Bobolinks leaving Venezuela, Common Nighthawks are probably not triggered to migrate southward from their first stationary non-breeding area (in December-January) by local declines in net primary productivity. The reasons why some nighthawks move 1800 km southward to Argentina for two to three months, whereas others remain in the Brazilian Cerrado, remain to be studied.
Recognizing the complexity of movements within South America is key to developing the tools and research to understand, among other questions, when and where feathers molt in long-distance migratory birds. Descriptions of molt and subspecies identification from North America were generally unsuitable for the birds we captured (half-way through the non-breeding season and nearly always mid-molt). Although molt-migration overlap is unusual, presumably because of the energetic costs (Newton 2009), we confirmed that at least one Common Nighthawk was molting P8 during its 1800-km migration from the northern Cerrado to Corrientes. Considering that 94% of birds captured between 1 December and 14 January (i.e., soon after arrival) were molting at least one flight feather, and for 70% this was a primary, we propose that the GPS-tagged individual is representative, and that many Common Nighthawks travel from the Cerrado to Corrientes while molting flight feathers. If the rate of molt while they are stationary at RNRSM is any indication, they may begin molting primaries well north of the Cerrado, perhaps in Central America. We tentatively suggest that Common Nighthawks employ a continuous molt-migration strategy (sensu Tonra and Reudink 2018). We caution that stable isotope studies of birds captured or collected on the breeding grounds (e.g., as recommended recently by Stiles et al. 2022) will likely provide limited information about molting and non-breeding areas unless researchers can sample many feathers from each individual. We suggest that isotope analysis would be most useful if researchers in breeding areas take advantage of incidental roadkills to collect all flight feathers (individually labeled) and a few body feathers from the same individual.
Our study illuminates some important potential threats to Common Nighthawks, even those that spend much of their annual cycle in protected areas. The birds that spend January to March at RNRSM may occupy protected native grasslands in Argentina and protected open pine forest in Florida but still face significant habitat loss and pesticide use on their first stationary non-breeding grounds in the Cerrado. There, large scale (industrial) agriculture has replaced 46% of the native vegetation (grassland and savannah), with 32% more predicted by 2050, leading to the expected extinction of 480 endemic plant species (Strassburg et al. 2017). In the Bananal/Cantão Island Protected Area (Tocantins, Brazil), an area used by one of our tagged nighthawks for at least 12 weeks, the area planted with soybeans recently increased by 1700% over a period of eight years, and 84% of farmers admitted to using banned pesticides (Moreira et al. 2019). Conversion to non-native vegetation and accompanying pesticide use are also widespread in the grasslands of Argentina (Etchegoyen et al. 2017), and could be an important driver of declines in multiple species of migratory birds (Jahn et al. 2017). Additionally, climate change is increasing the severity, intensity, and size of wildfires throughout the non-breeding habitat of Common Nighthawks (Ellis et al. 2022, Saucedo et al. 2023). Although nighthawks are known to rest in and forage over recently-burned areas, it is not clear how these wildfires and thick smoke affect their food supply and ability to forage.
We have demonstrated that deploying and re-deploying a small number of GPS loggers, as part of a multi-year, site-based monitoring program in South America, can provide critical data on the migratory movements, daily activity patterns, and use of non-breeding habitat (and eventually habitat selection) of a widely-ranging migratory aerial insectivore. We encourage international partnerships and networks to donate (new and used) GPS loggers to other multi-year monitoring projects led by Latin American teams, such as the growing network of Brazilian bird observatories (Figueira 2021). Such efforts are likely to have an outsized impact on our understanding of habitat use, and help fill major gaps in our understanding of the ecology of Neotropical-Nearctic migrants across their annual cycle.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
LGP, NF, OV and AB developed the questions and designed the study in collaboration with KLC and ARN; NF and OV led the field research with support from LGP, AB, KLC, and EDS; KLC, ARN, NF, EDS, and OV obtained funding, with significant help from Wendy Easton and Krista De Groot; NF, OV, and KLC managed the data; KLC analyzed the data and wrote most of the paper; all authors contributed to literature review, fact-checking, revising and editing. Please see Appendix 3 for a reflexivity statement describing how equity was promoted during this project.
ACKNOWLEDGMENTS
For their support of the project, we thank Lisandro Cardinale, Wendy Easton, Facundo Di Sallo, Carlos Ferreyra, Agostina Juncosa, Krista De Groot, Kathy Martin, Diego Monteleone, Mariela Pilar Morales, Helen Pargeter, Diego Pérez, Gabriel Ruiz Díaz, and Agustina Tello. For funding and equipment we are grateful to Neotropical Grassland Conservancy, Idea Wild, Environment & Climate Change Canada, Eastern Bird Banding Association, International Conservation Fund of Canada, Centro de Investigaciones del Bosque Atlántico (CeIBA), Maxwell Hanrahan Foundation and Wilson Ornithological Society. The Dirección de Parques y Reservas de la Provincia de Corrientes and the Entidad Binacional Yacyretá provided research permits and accommodation. Fundación Temaikèn financed and facilitated data collection at Reserva Urutaú, which was conducted with permission from IMiBio and the Ministerio de Ecología y RNR de Misiones.
DATA AVAILABILITY
The datasets generated during the current study are available in the Movebank Data Repository, https://doi.org/10.5441/001/1.287 (Fariña N., O. Villalba, L. G. Pagano, A. Bodrati, E. Stein, A. R. Norris, and K. L. Cockle. 2023).
LITERATURE CITED
Åkesson, S., P. W. Atkinson, A. Bermejo, J. de la Puente, M. Ferri, C. M. Hewson, J. Holmgren, E. Kaiser, L. Kearsley, R. H. G. Klaassen, et al. 2020. Evolution of chain migration in an aerial insectivorous bird, the common swift (Apus apus). Evolution 74(10):2377-2391. https://doi.org/10.1111/evo.14093
Albert, S., J. D. Wolfe, J. Kellerman, T. Sherry, B. J. M. Stutchbury, N. J. Bayly, and A. Ruiz-Sánchez. 2020. Habitat ecology of Nearctic-Neotropical migratory landbirds on the nonbreeding grounds. Ornithological Applications 122(4):duaa055. https://doi.org/10.1093/condor/duaa055
Aldridge, H. D. J. N., and R. M. Brigham. 1991. Factors influencing foraging time in two aerial insectivores: the bird Chordeiles minor and the bat Eptesicus fuscus. Canadian Journal of Zoology 69(1):62-69. https://doi.org/10.1139/z91-010
Azpiroz, A. B., J. P. Isacch, R. A. Días, A. S. Di Giacomo, C. S. Fontana, and C. Morales Palarea. 2012. Ecology and conservation of grassland birds in southeastern South America: a review. Journal of Field Ornithology 83(3):217-246. https://doi.org/10.1111/j.1557-9263.2012.00372.x
Bayly, N. J., L. Cárdenas Ortiz, M. Rubio, and C. Gómez. 2014. Migration of raptors, swallows and other diurnal migratory birds through the Darien of Colombia. Ornitología Neotropical 25:63-71.
Belton, W. 1978. Supplementary list of new birds for Rio Grande do Sul, Brazil. Auk 95(2):413-415. https://www.jstor.org/stable/4085464
Bodrati, A. 2005. Notas sobre la avifauna del Parque Nacional Chaco, el Parque Provincial Pampa del Indio y otros sectores de la provincia de Chaco, Argentina. Nuestras Aves 49:15-23.
Bodrati, A., N. Fariña, O. Villalba, L. Pagano, and K. L. Cockle. 2019. Notas sobre la biología reproductiva y el ciclo anual del Atajacaminos Ala Negra (Eleothreptus anomalus) en Argentina. Ornitología Neotropical 30:157-162. https://doi.org/10.58843/ornneo.v30i0.493
Bravo, S. P., V. R. Cueto, and C. A. Gorosito. 2017. Migratory timing, rate, routes and wintering areas of White-crested Elaenia (Elaenia albiceps chilensis), a key seed disperser for Patagonian forest regeneration. PLoS ONE 12(2):e0170188. https://doi.org/10.1371/journal.pone.0170188
Brigham, R. M. 1990. Prey selection by Big Brown Bats (Eptesicus fuscus) and Common Nighthawks (Chordeiles minor). American Midland Naturalist 124(1):73-80. https://doi.org/10.2307/2426080
Brigham, R. M., J. Ng, R. G. Poulin, and S. D. Grindal. 2020. Common Nighthawk (Chordeiles minor), version 1.0. In A. F. Poole, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.comnig.01
Brodkorb, P. 1938. Further additions to the avifauna of Paraguay. Occasional Papers of the Museum of Zoology, University of Michigan 394:1-5.
Burkart, R., N. O. Bárbaro, R. O. Sánchez, and D. A. Gómez. 1999. Eco-Regiones de la Argentina. Administración de Parques Nacionales - Programa de Desarrollo Institucional Ambiental. Presidencia de la Nación, Administración de Parques Nacionales. Buenos Aires, Argentina.
Cabrera, A. L. 1971. Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica 14:1-42.
Cohn-Haft, M. 2012. Observation number XC98994. https://xeno-canto.org/contributor/OXIRTCXSVH
Costa, T. V. V., A. Bodrati, and K. L. Cockle. In press. Neotropical nightjars: what is known and what is still missing? In C. Camacho, editor. Nightjars: from mystery to model in ecology and evolution. Wildlife Research Monographs, Springer, Cham, Switzerland.
Coues, E. 1888. New forms of North American Chordiles. Auk 5:37.
Cuello, J. 1966. First specimens of Buteo albicaudatus and Chordeiles minor in Uruguay. Condor 68(3):301. https://doi.org/10.2307/1365565
Dornas, T., and R. T. Pinheiro. 2011. Ilha do Bananal e Planície do Cantão. Pages 111-115 in R. M. Valente, J. M. Cardoso da Silva, F. C. Straube, and J. L. X. do Nascimento, editors. Conservação de aves migratórias neárticas no Brasil. Conservação Internacional, Belém, Brazil.
Dexter, R. W. 1961. Further studies on nesting of the Common Nighthawk. Bird Banding 32(2):79-85. https://doi.org/10.2307/4510861
Ellis, T. M., D. M. J. S. Bowman, P. Jain, M. D. Flannigan, and G. J. Williamson. 2022. Global increase in wildfire risk due to climate‐driven declines in fuel moisture. Global Change Biology 28:1544-1559. https://doi.org/10.1111/gcb.16006
Environment Canada. 2016. Recovery strategy for the Common Nighthawk (Chordeiles minor) in Canada. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa, Ontario, Canada.
Etchegoyen, M. A., A. E. Ronco, P. Almada, M. Abelando, and D. J. Marino. 2017. Occurrence and fate of pesticides in the Argentine stretch of the Paraguay-Paraná basin. Environmental Monitoring and Assessment 189:63. https://doi.org/10.1007/s10661-017-5773-1
Fariña, N., O. Villalba, L. G. Pagano, A. Bodrati, E. Stein, A. R. Norris, and K. L. Cockle. 2023. Data from: Non-breeding ecology of a Neotropical-Nearctic migrant, the Common Nighthawk (Chordeiles minor): habitat, activity patterns, molt, and migration. Movebank Data Repository. https://doi.org/10.5441/001/1.287
Ferreira, L. G., and A. R. Huete. 2004. Assessing the seasonal dynamics of the Brazilian Cerrado vegetation through the use of spectral vegetation indices. International Journal of Remote Sensing 25(10):1837-1860. https://doi.org/10.1080/0143116031000101530
Figueira, L. 2021. O que são observatórios de aves? Livro de Resumos, XXVII Congresso Brasileiro de Ornitologia, 1-5 August 2021.
Fraser, K. C., A. Shave, A. Savage, A. Ritchie, K. Bell, J. Siegrist, J. D. Ray, K. Applegate, and M. Pearman. 2017. Determining fine-scale migratory connectivity and habitat selection for a migratory songbird by using new GPS technology. Journal of Avian Biology 48(3):339-345. https://doi.org/10.1111/jav.01091
García-Pérez, B., and K. A. Hobson. 2014. A multi-isotope (δ2H, δ13C, δ15N) approach to establishing migratory connectivity of Barn Swallow (Hirundo rustica). Ecosphere 5(2):21. http://dx.doi.org/10.1890/ES13-00116.1
Guzy, M. J. 2020. Antillean Nighthawk (Chordeiles gundlachii), version 1.0. In A. F. Poole and F. B. Gill, editors. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.antnig.01
Hansen, H. O. 1998. Plan de manejo Reserva Natural Santa María. Fundación Facultad de Ciencias Exactas, Química y Naturales, Universidad Nacional de Misiones, Posadas, Misiones, Argentina.
Hartert, E., and S. Venturi. 1909. Notes sur les oiseaux de la Republique Argentine. Novitates Zoologicae 16:159-267. https://doi.org/10.5962/bhl.part.21963
Heckscher, C. M., D. Mendez, K. A. Hobson, A. A. Aispuro, N. Kleponis, A. H. Kneidel, M. L. Mancuso, and K. Kardynal. 2021. Miniature GPS data loggers reveal habitat affiliations and movement of veeries (Catharus fuscescens) during their first non-transient period in South America. Ornitología Neotropical 32(1):1-9. https://doi.org/10.58843/ornneo.v32i1.777
Heckscher, C. M., S. M. Taylor, J. W. Fox, and V. Afanasyev. 2011. Veery (Catharus fuscescens) wintering locations, migratory connectivity, and a revision of its winter range using geolocator technology. Auk 128(3):531-542. https://doi.org/10.1525/auk.2011.10280
Imlay, T. L., K. A. Hobson, A. Roberto-Charron, and M. L. Leonard. 2018. Wintering areas, migratory connectivity and habitat fidelity of three declining Nearctic-Neotropical migrant swallows. Animal Migration 5(1):1-16. https://doi.org/10.1515/ami-2018-0001
Ingels, J., Y. Oniki, and E. O. Willis. 1999. Opportunistic adaptations to man-induced habitat changes by some South American Caprimulgidae. Revista Brasileira de Biologia 59(4):563-566. https://doi.org/10.1590/S0034-71081999000400005
Jahn, A. E., V. Bejarano, V. R. Cueto, A. S. Di Giacomo, and C. S. Fontana. 2017. Movement ecology research to advance conservation of South America’s grassland migratory birds. Perspectives in Ecology and Conservation 15(3):209-215. http://dx.doi.org/10.1016/j.pecon.2017.07.003
Johnson, E. I., and J. D. Wolfe. 2017. Molt in Neotropical birds: life history and aging criteria. Studies in Avian Biology 51. CRC Press, Boca Raton, Florida, USA. https://doi.org/10.4324/9781315119755
Knight, E. C., R. M. Brigham, and E. Bayne. 2021b. Specialist or generalist? It depends. Context-dependent habitat relationships provide insight into forest disturbance effects for a boreal bird species. Forest Ecology and Management 502:119720. https://doi.org/10.1016/j.foreco.2021.119720
Knight, E. C., R. M. Brigham, and E. R. Bayne. 2022. The big boom theory: the Common Nighthawk wing-boom display delineates exclusive nesting territories. Ornithology 139(1):ukab066. https://doi.org/10.1093/ornithology/ukab066
Knight, E. C., J. W. Ng, C. E. Mader, R. M. Brigham, and E. M. Bayne. 2018. “An inordinate fondness for beetles”: first description of Common Nighthawk (Chordeiles minor) diet in the boreal biome. Wilson Journal of Ornithology 130(2):525-531. https://doi.org/10.1676/16-219.1
Knight, E. C., A.-L. Harrison, A. L. Scarpignato, S. L. Van Wilgenburg, E. M. Bayne, J. W. Ng, E. Angell, R. Bowman, R. M. Brigham, B. Drolet, et al. 2021a. Comprehensive estimation of spatial and temporal migratory connectivity across the annual cycle to direct conservation efforts. Ecography 44(5):665-679. https://doi.org/10.1111/ecog.05111
Knight, E. C., A. C. Smith, R. M. Brigham, and E. M. Bayne. 2021c. Combination of targeted monitoring and Breeding Bird Survey data improves population trend estimation and species distribution modeling for the Common Nighthawk. Ornithological Applications 123(2):duab005. https://doi.org/10.1093/ornithapp/duab005
Kramer, G. R., D. E. Andersen, D. A. Buehler, P. B. Wood, S. M. Peterson, J. A. Lehman, K. R. Aldinger, L. P. Bulluck, S. Harding, J. A. Jones, et al. 2018. Population trends in Vermivora warblers are linked to strong migratory connectivity. Proceedings of the National Academy of Sciences 115(14):E3192-E3200. https://doi.org/10.1073/pnas.1718985115
Krauczuk, E., and A. S. Di Giacomo. 2005. Reserva Natural Rincón de Santa María. Pages 147-148 in A. S. Di Giacomo, editor. Áreas importantes para la conservación de las aves en la Argentina. Sitios prioritarios para la conservación de la biodiversidad. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires, Argentina.
McGuire, L. P., J. G. Boyles, and R. M. Brigham. 2021. Lack of foraging site fidelity between years by Common Nighthawks (Chordeiles minor). American Midland Naturalist 185(1):139-144. https://doi.org/10.1674/0003-0031-185.1.139
Michel, N. L., A. C. Smith, R. G. Clark, C. A. Morrissey, and K. A. Hobson. 2016. Differences in spatial synchrony and interspecific concordance inform guild-level population trends for aerial insectivorous birds. Ecography 39:774-786. https://doi.org/10.1111/ecog.01798
Miller, A. H. 1925. The boom-flight of the Pacific Nighthawk. Condor 27(4):141-143. https://doi.org/10.2307/1362990
Montiel, M. del R., M. G. Gatti, J. L. Fontana, I. Zanone, and S. C. Zaninovich. 2016. Caracterización de las comunidades vegetales de la Reserva Natural Rincón de Santa María (Ituzaingó, Corrientes) y el impacto de la represa hidroeléctrica Yacyretá sobre ellas. Extensionismo, Innovación y Transferencia Tecnológica - Claves para el Desarrollo 3:243-250. https://doi.org/10.30972/eitt.302767
Moreira, D. C., E. Collicchio, and F. B. Gamba. 2019. Panorama do cultivo e produtividades da soja na APA Ilha do Bananal/Cantão Tocantins: Safras 2008/2009 a 2015/2016. Journal of Bioenergy and Food Science 6(4):119-131. https://doi.org/10.18067/jbfs.v6i4.262
Navas, J. R., and N. A. Bó. 1988. Aves nuevas o poco conocidas de Misiones, Argentina. II. Comunicaciones Zoológicas del Museo de Historia Natural de Montevideo 12:1-9.
Nebel, S., A. Mills, J. D. McCracken, and P. D. Taylor. 2010. Declines of aerial insectivores in North America follow a geographic gradient. Avian Conservation and Ecology 5(2):1. https://doi.org/10.5751/ACE-00391-050201
Newton, I. 2009. Moult and plumage. Ringing and Migration 24(3):220-226. https://doi.org/10.1080/03078698.2009.9674395
Ng, J. W., E. C. Knight, A. L. Scarpignato, A.-L. Harrison, E. M. Bayne, and P. P. Marra. 2018. First full annual cycle tracking of a declining aerial insectivorous bird, the Common Nighthawk (Chordeiles minor), identifies migration routes, nonbreeding habitat, and breeding site fidelity. Canadian Journal of Zoology 96(8):869-875. https://doi.org/10.1139/cjz-2017-0098
Oberholser, H. C. 1914. A monograph of the genus Chordeiles Swainson, type of a new family of goat-suckers. Bulletin of the United States National Museum 86. Smithsonian Institution, Washington, D.C., USA. https://doi.org/10.5479/si.03629236.86.i
Olrog, C. C. 1979. Nueva lista de la avifauna argentina. Opera Lilloana 27:1-324.
Perlut, N., and A. Levesque. 2020. Light-level geolocation reveals the migration route and non-breeding location of an Antillean Nighthawk (Chordeiles gundlachii). Journal of Caribbean Ornithology 33:49-53. https://doi.org/10.55431/jco.2020.33.49-53
Pyle, P. 1997. Identification guide to North American birds. Part I. Slate Creek Press, Point Reyes Station, California, USA.
Renfrew, R. B., S. J. K. Frey, and J. Klavins. 2011. Phenology and sequence of the complete prealternate molt of Bobolinks in South America. Journal of Field Ornithology 82(1):101-113. https://doi.org/10.1111/j.1557-9263.2010.00312.x
Renfrew, R. B., D. Kim, N. Perlut, J. Smith, J. Fox, and P. Marra. 2013. Phenological matching across hemispheres in a long-distance migratory bird. Diversity and Distributions 19(8):1008-1019. https://doi.org/10.1111/ddi.12080
Rust, H. J. 1947. Migration and nesting of nighthawks in northern Idaho. Condor 49(5):177-188. https://doi.org/10.2307/1364338
Salvador, S. A., and A. Bodrati. 2017. Biología reproductiva y otros aspectos de la historia natural del Ñacunda (Chordeiles nacunda nacunda) en Argentina. Historia Natural 7:39-54.
Saucedo, G. I., R. Perucca, and D. Kurtz. 2023. Las causas de los incendios de principio del año 2002 en la provincia de Corrientes. Ecología Austral 33(1):273-284. https://doi.org/10.25260/EA.23.33.1.0.2020
Selander, R. K. 1954. A systematic review of the booming Nighthawks of western North America. Condor 56(2):57-82. https://doi.org/10.2307/1364662
Somenzari, M., P. P. do Amaral, V. R. Cueto, A. C. Guaraldo, A. E. Jahn, D. Mendes Lima, P. Cerqueira Lima, C. Lugarini, C. G. Machado, J. Martinez, J. L. X. do Nascimento, et al. 2018. An overview of migratory birds in Brazil. Papéis Avulsos de Zoologia 58:e20185803. http://doi.org/10.11606/1807-0205/2018.58.03
Spiller, K. J., and R. Dettmers. 2019. Evidence for multiple drivers of aerial insectivore declines in North America. Condor 121(2):duz010. https://doi.org/10.1093/condor/duz010
Steullet, A. B., and E. A. Deautier. 1935-1946. Catálogo sistemático de las aves de la República Argentina. Obra del Cincuentenario del Museo de La Plata 1:1-1006.
Stiles, F. G., O. Acevedo-Charry, and A. M. Cuervo. 2022. The first Colombian records of the Antillean Nighthawk (Chordeiles gundlachii), with notes on migrant Chordeiles in South America. Ornitología Colombiana 22:16-24. https://doi.org/10.59517/oc.e541
Stotz, D. F., R. O. Bierregaard, M. Cohn-Haft, P. Petermann, J. Smith, A. Whittaker, and S. V. Wilson. 1992. The status of North American migrants in Central Amazonian Brazil. Condor 94(3):608-621. https://doi.org/10.2307/1369246
Strassburg, B. B. N., T. Brooks, R. Feltran-Barbieri, A. Iribarrem, R. Crouzeilles, R. Loyola, A. E. Latawiec, F. J. B. Oliveira Filho, C. A. de M. Scaramuzza, F. R. Scarano, B. Soares-Filho, and A. Balmford. 2017. Moment of truth for the Cerrado hotspot. Nature Ecology and Evolution 1:0099. https://doi.org/10.1038/s41559-017-0099
Taylor, P. 2009. Late-summer feeding and migration behaviour and numerical trends of Common Nighthawks, Chordeiles minor, near Pinawa, Manitoba, 1976-2009. Canadian Field-Naturalist 123(4):338-345. https://doi.org/10.22621/cfn.v123i4.1003
Tonra, C. M., and M. W. Reudink. 2018. Expanding the traditional definition of molt-migration. Auk 135(4):1123-1132. https://doi.org/10.1642/AUK-17-187.1
Vassallo M. M., H. D. Dieguez, M. F. Garbulsky, E. G. Jobbágy, and J. M. Paruelo. 2013. Grassland afforestation impact on primary productivity: a remote sensing approach. Applied Vegetation Science 16:390-403. https://doi.org/10.1111/avsc.12016
Verón, S. M. 2021. Primer registro del Querequeté (Chordeiles gundlachii) para Argentina. El Hornero 36(2):127-130. https://doi.org/10.56178/eh.v36i2.376
Zotta, A. R. 1937. Lista sistemática de las aves argentinas. El Hornero 6(3):531-554.
Fig. 1
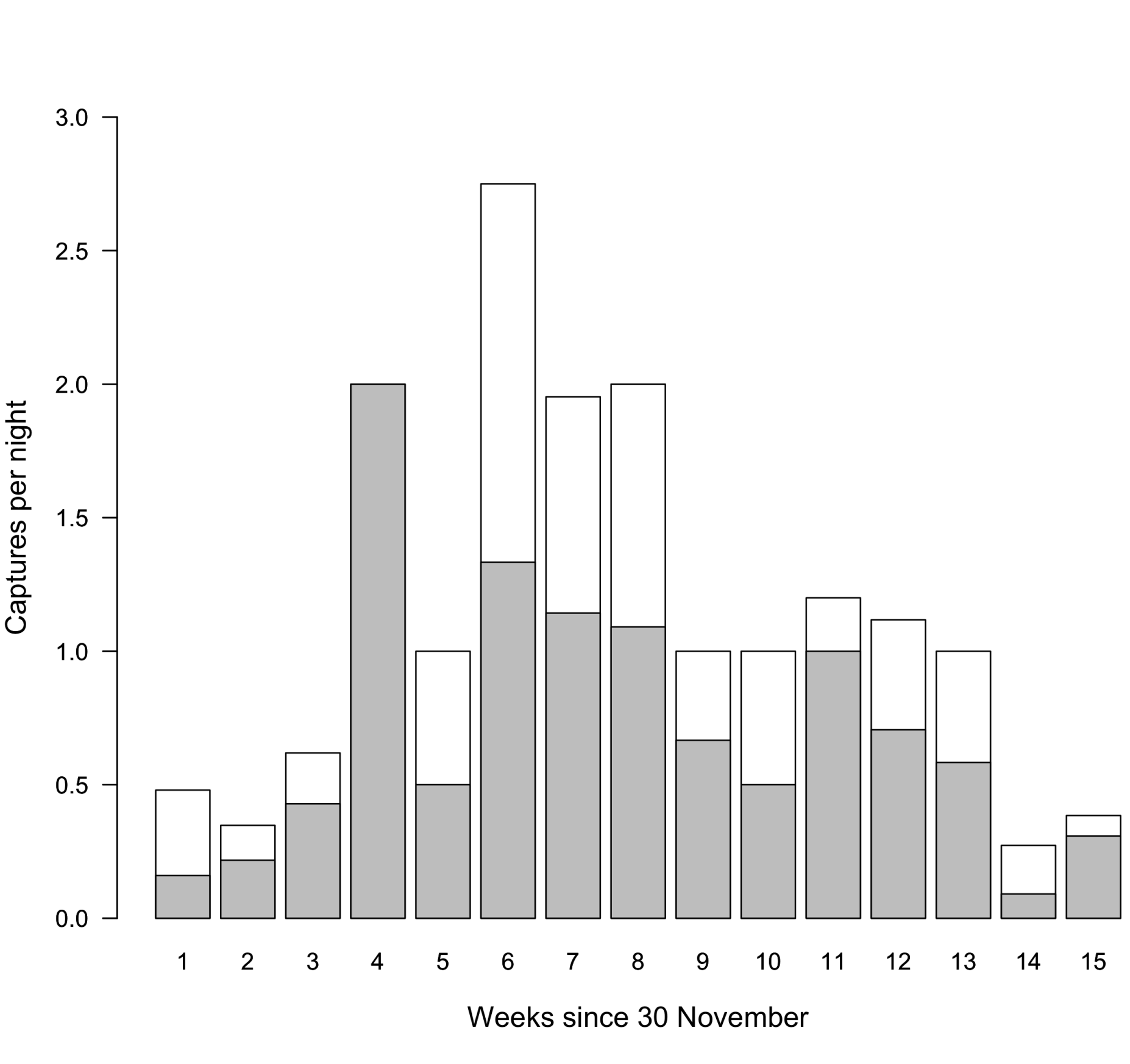
Fig. 1. Total number of captures of Common Nighthawk females (gray) and males (white) per sampling night, by week (30 November to 15 March), at Reserva Natural Rincón de Santa María, Corrientes, 2017–2022.
Fig. 2
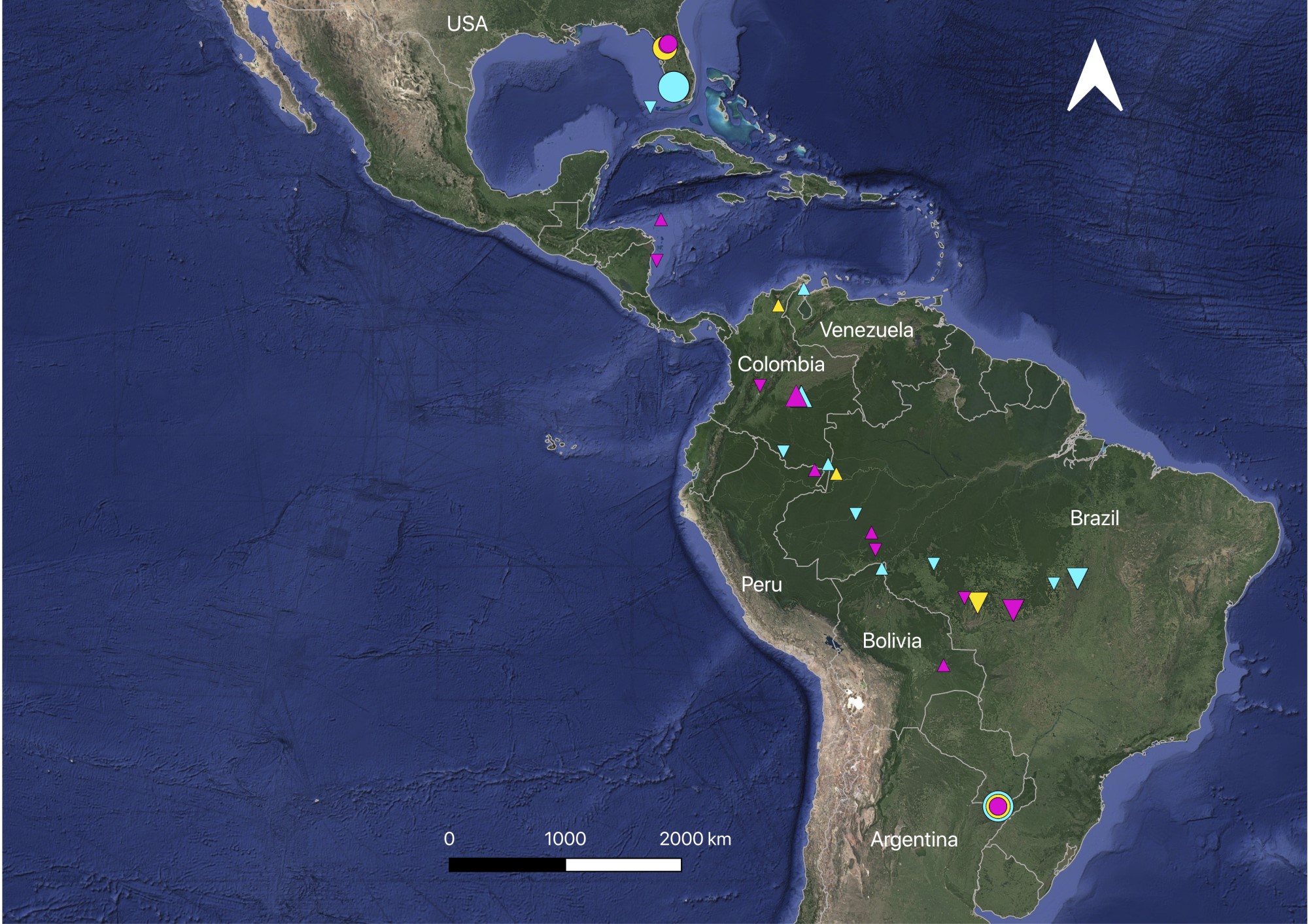
Fig. 2. Locations of three adult female Common Nighthawks across the annual cycle, from Pinpoint-10 GPS loggers affixed to birds in Reserva Natural Rincón de Santa María, Corrientes, Argentina (southern concentric circles) in January 2019 (Band RSM140; yellow; Band RSM198: turquoise) and January 2020 (Band RSM152: pink). Northernmost circles indicate breeding grounds in Florida, USA (GPS locations throughout May–August). Triangles pointing upward indicate GPS fixes on northward migration toward breeding grounds (late March to late April); triangles pointing downward indicate GPS fixes on southward migration toward RNRSM (September to January). Small triangles indicate single points (birds in transit); large triangles indicate multiple GPS fixes over a period of 2–12 weeks.
Fig. 3
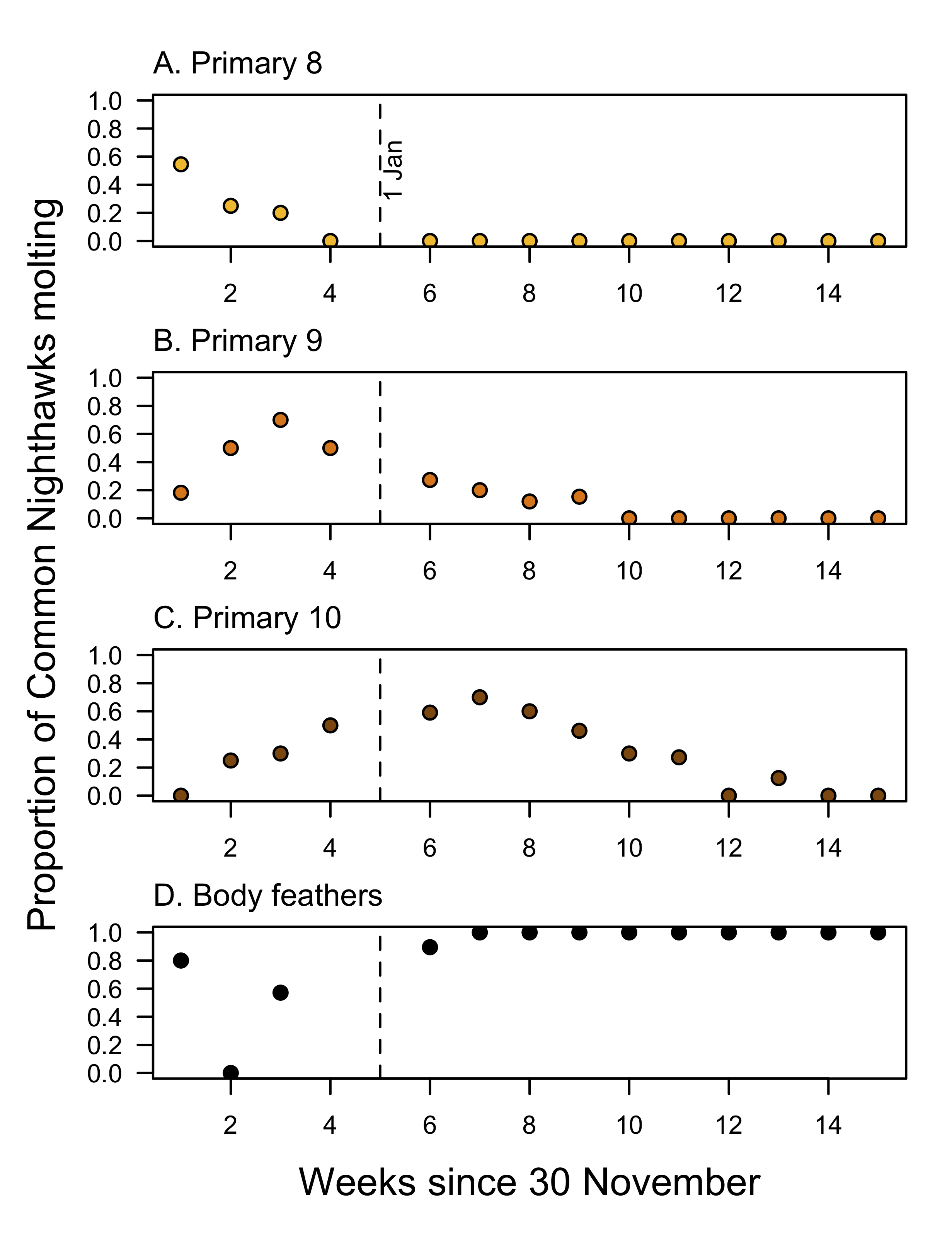
Fig. 3. Proportion of Common Nighthawks molting (A) Primary 8, (B) Primary 9, (C) Primary 10, and (D) body feathers, across the 15 weeks from 30 November to 15 March, based on 223 captures at Reserva Natural Rincón de Santa María, Corrientes, Argentina.
Fig. 4
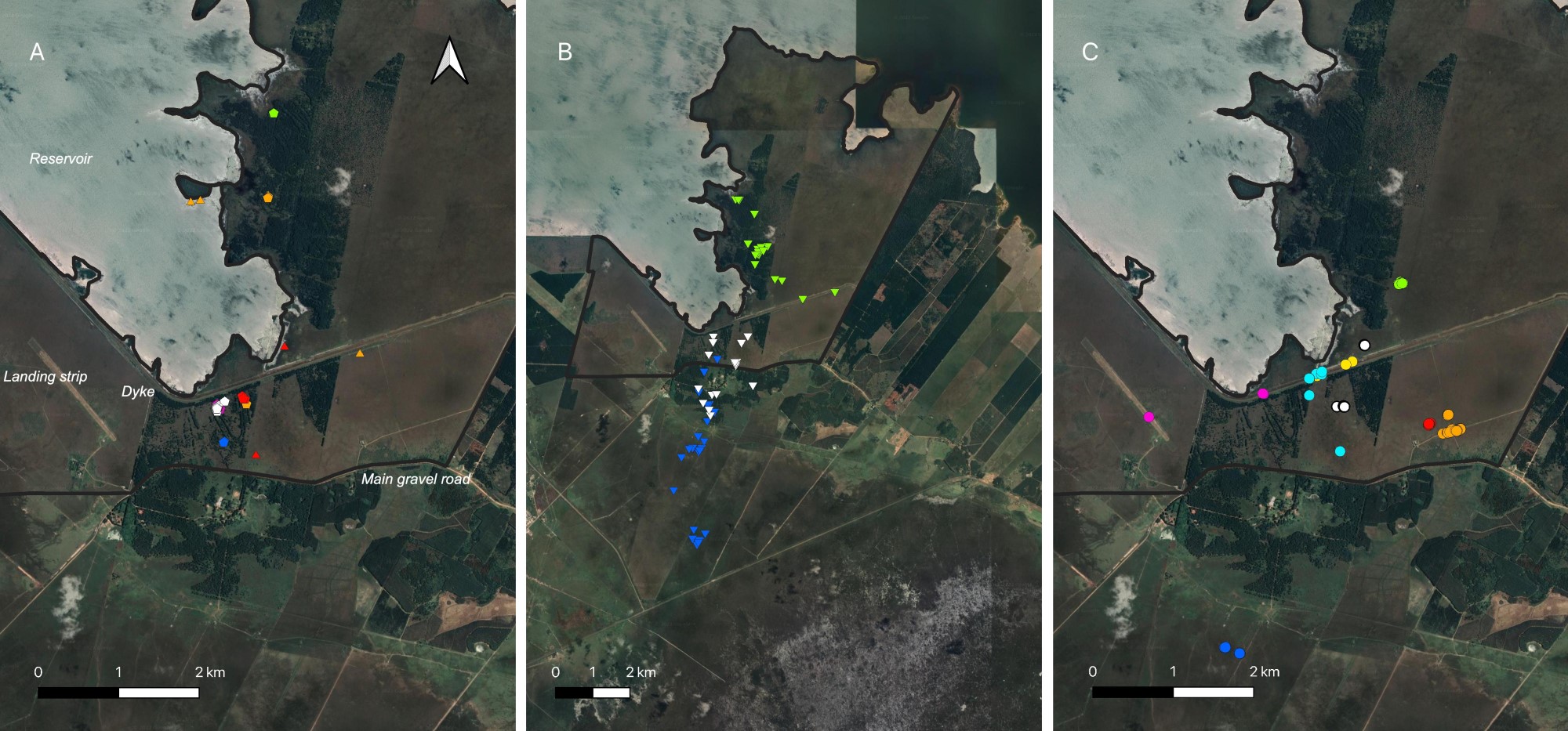
Fig. 4. Locations (colored points) of 8 adult female Common Nighthawks tracked using Pinpoint-10 GPS loggers at Reserva Natural Rincón de Santa María, Corrientes, Argentina (reserve outlined in black). Individual birds are identified by color. Yellow (RSM140; 18 January – 19 March 2019), turquoise (RSM198; 18 January – 15 March 2019), and pink (RSM152; 25 January – 15 March 2020) symbols correspond to the individuals tracked to Florida and shown in Figure 2. White (RSM429; 19–21 January 2021), green (RSM401; 18–21 December 2021), red (RSM424; 24–29 January 2022), orange (RSM1152; 24–30 January 2022), and blue (RSM1151; 8–10 February 2022) symbols indicate birds that were only tracked locally. Brown and light green areas are grasslands and dark green are mature exotic pine or eucalyptus, either plantations or invasions. (A) Daytime roost sites (pentagons; six individuals) and records within 45 min before sunrise (upward pointing triangles; two individuals). (B) Records within 45 min after sunset (downward pointing triangles; three individuals). (C) Records throughout the night (circles; eight individuals; many points hidden).