







The following is the established format for referencing this article:
Fernández, G., J. Bonanomi, and J. de Pinho. 2023. Seasonal drivers of mixed-species flocks from tropical savannas: insights from the Pantanal wetland. Journal of Field Ornithology 94(3):3.ABSTRACT
Birds participate in different types of multi-species groups (e.g., mixed-species flocks) occurring from temperate to tropical forest and also savannas. However, the effects of seasonal variation in flocking dynamics and formation in tropical savannas, especially in seasonally flooded wetlands, have received comparatively less attention than tropical forests and temperate ecosystems. Because multi-species groups can reflect specific aspects of the bird community (e.g., temporal dynamics), the present study aims to describe the structure, organization, and seasonal variation of mixed-species flocks in the Pantanal, the world’s largest tropical wetland. During the dry, flood, and rainy seasons of 2019 and the flood season of 2020, we observed flocks within the natural mosaic of grassy-shrubby vegetation and patchy forest, and recorded the number of species, individuals (per species), and diversity. Seasonality had a strong influence on flocking structure and organization. During the rainy season, flocks had significantly fewer species, individuals, and diversity, whereas during the dry and flood seasons, flocks had more species, individuals, and diversity. Moreover, we suggest Polioptila dumicola and Casiornis rufus to act, respectively, as the nuclear and the sentinel species of flocks. These species were present in flocks during the year, but they were less frequent during the rainy season. The breeding period during the rainy season may negatively influence the formation of mixed-species flocks, whereas the access to food resources and predator protection may instead facilitate their formation in the dry and flood seasons, respectively. Our results help fill a knowledge gap on how mutualistic interactions among bird species are organized and vary in seasonal and heterogeneous environments, and also provide a starting point to subsequently understand the species role and the effects of human-induced environmental changes on mixed-species bird flock dynamics in the Pantanal and other world’s savannas and wetlands facing rapid landscape conversion.
RESUMEN
Las aves participan en diferentes tipos de grupos multi especies (e.g. bandadas mixtas) que ocurren desde bosques templados hasta tropicales y también en sabanas. Sin embargo, los efectos de la variación estacional en la dinámica y formación de bandadas en sabanas tropicales, especialmente en humedales inundables estacionales, ha recibido comparativamente menor atención que los ecosistemas de bosques tropicales y templados. Debido a que los grupos multi especies pueden reflejar aspectos específicos de la comunidad de aves (e.g. dinámica temporal), el objetivo de este estudio fue describir la estructura, organización y variación estacional de las bandadas mixtas en el Pantanal, el humedal más grande del mundo. Durante las temporadas seca, inundación y lluviosa del 2019 y la temporada de inundación del 2020, observamos las bandadas en un mosaico natural de vegetación de pastos y arbustiva y de parches de bosque y registramos el número de especies, individuos (por especie) y diversidad. La estacionalidad tuvo una influencia fuerte sobre la estructura y organización de las bandadas. Durante la temporada lluviosa, las bandadas tuvieron significativamente un menor número de especies, individuos y diversidad mientras que durante las temporadas secas y de inundación, las bandadas tuvieron más especies, individuos y diversidad. Sugerimos que Polioptila dumicola y Casiornis rufus actúan como especies nucleares y centinelas en la bandada, respectivamente. Estas especies estuvieron presentes en las bandadas a lo largo del año, pero fueron menos frecuentes durante la temporada lluviosa. El periodo de reproducción durante la temporada lluviosa puede influenciar negativamente la formación de bandadas mixtas de especies, mientras que el acceso a los recursos alimenticios y protección contra los depredadores puede facilitar su formación en las temporadas secas e inundable, respectivamente. Nuestros resultados ayudan a llenar el vacío de información sobre como las interacciones mutualistas entre especies de aves están organizadas y varían en ambientes estacionales y heterogéneos, y proveen un punto de partida para posteriormente comprender los roles de las especies y los efectos de los cambios ambientales inducidos por los humanos en la dinámica de las bandadas mixtas de especies en el Pantanal y otras sabanas y humedales del mundo con altas tasas de transformación del paisaje.
INTRODUCTION
Life in groups is a common feature across animal lineages. Mammals and fishes form herds and shoals, while invertebrates such as termites and ants assemble in cooperative colonies (Goodale et al. 2017). However, birds establish the most variable examples of living in groups, including individuals associated in single- and/or multi-species groups, temporal or permanent, occurring from tropical to temperate regions and from forests to savannas (Morse 1977, Greenberg 2000, Zou et al. 2018, Goodale et al. 2020). Mixed-species flocks are groups of individuals of two or more species that move and forage at close proximity over time (Goodale et al. 2009). Those species that stimulate the formation and cohesion of flocks are the nuclear species, whereas those species that tend to join the nuclear ones, increasing the flocking size, are the attendant species. Sentinel species are those that warn of predator presence (Moynihan 1962, Powell 1985, Greenberg 2000). Bird species may join mixed-species flocks to increase their foraging efficiency and/or their predator avoidance, the two main non-mutually exclusive hypotheses behind this behavior (Greenberg 2000, Sridhar et al. 2009, Goodale et al. 2020). Thus, several specific aspects about the life history of birds, such as annual survival, can be addressed through the study of mixed-species flocks.
In tropical regions, birds form complex mixed-species flocks, e.g., the same individuals of some species keeping in the same flock throughout the day and year, from rainforests (Munn and Terborgh 1979, Bell 1983, Munn 1985, Jullien and Thiollay 1998) to dry savannas (Alves and Cavalcante 1996, Vanderduys et al. 2012). Different from the temperate systems, mixed-species flocks from tropics are present throughout the year, but they are less frequent during the breeding period, maintaining and defending a stable territory (Powell 1985, Greenberg 2000, Jayarathna et al. 2013, Rutt and Stouffer 2021). Although research has mostly focused on mixed-species flocks from forests (reviewed in Zou et al. 2018), it is known that mixed-species flocks from non-forest ecosystems (e.g., savannas) can show similar properties to mixed-species flocks of forest birds, such as high stability and high species participation (Tubelis 2007). Thus, the study of mixed-species flocks from tropical savannas may also help us understand the bird communities inhabiting these environments. For example, the negative impacts of habitat fragmentation can be reflected on both flocking and community structure (Goodale et al. 2015). Seasonality can also be addressed by examining the variation of species composition of mixed-species flocks (Alves and Cavalcanti 1996, Tubelis 2004, Vanderduys 2012). However, the effects of seasonal variation have been poorly studied (Macedo 2002, Tubelis 2007, Lees et al. 2020), especially in those temporarily flooded tropical savannas (Fernández-Arellano et al. 2021). Hence, the study of avian mixed-species flocks from seasonally flooded tropical savannas may provide a realistic and evidence-based framework to understand their structure, organization, and temporal dynamics at local and/or landscape scale, and how they would respond to global human-induced environmental change.
The Pantanal is the largest tropical wetland and harbors one of the most species-rich bird faunas in the world (Tubelis and Tomás 2003). Bird richness in this ecosystem represents about 30% of the Brazilian avifauna, and species inhabit different ecological niches and types of vegetation (Sick 1997, Gwynne et al. 2010). Birds also participate in mixed-species flocks through a gradient of grassy-shrubby-forest vegetation (Amaral and Ragusa-Netto 2008), which can also be strongly influenced by seasonal change in the hydrological cycle (de Pinho et al. 2017). Moreover, the Pantanal comprises a natural mosaic of patchy forests, savannas, and grasslands, covered by water during the flood season (January to March), but more accessible during the dry (May to September) and the beginning of the rainy (October to April) season (Junk et al. 2006, Alho 2008, Alho et al. 2019). However, these mosaics of heterogeneous environments are increasingly threatened by a rapid landscape conversion to both agriculture and cattle raising activities, and by an increase of wildfires (Tomas et al. 2019). The extreme 2020 fire impacted up to 40% of the biome area (Garcia et al. 2021, Pletsh et al. 2021), and damage also extended to diverse vertebrate groups such as large-sized mammals and birds (Tomas et al. 2021). Likewise, the pervasive spread of woody vegetation into the native grassy-shrubby environments (i.e., islets of scattered shrubby and woody vegetation inserted into the vast and temporally flooded matrix of grassy vegetation, so-called earthmound fields) can threaten the native biodiversity (Nunes da Cunha and Junk 2004, Dorado-Rodrigues et al. 2015, Barbosa da Silva et al. 2016). Particularly, bird communities inhabiting savannas can be restructured (e.g., changes in species composition) by the spread of shrubby vegetation into grasslands (Sirami et al. 2009, Stanton et al. 2018). Evidence shows that grassland-dependent, more habitat-specialist bird species are mostly unable to adapt to perturbations associated with shrub encroachment, thus reducing their distribution in the grassy vegetation (Andersen and Steidel 2019). In seasonally flooded tropical savannas, anthropogenic changes in landscape and microclimatic perturbations may also have critical effects on flood pulse (Marengo et al. 2016, Thielen et al. 2020), with relevant impacts on bird communities (Tubelis and Tomás 1999).
However, pervasive knowledge gaps about the influence of seasonality, landscape structure, or ecological processes on bird communities of temporally flooded tropical savannas still remain. Because the Pantanal is a bird-rich wetland composed by a heterogeneous and highly dynamic landscape and faces the impacts of several anthropogenic actions and extreme climate events (e.g., prolonged droughts), with unexplored consequences on bird communities, mixed-species flocks constitute practical case studies to understand the spatial and temporal dynamics of bird communities from tropical savannas. We study the influence of seasonality (i.e., dry, rainy, and flood season) on both the structure and organization of mixed-species flocks inhabiting the grassy-shrubby vegetation (hereafter, earthmound fields) of the Pantanal. We hypothesize that the number of species, number of individuals, and species diversity are reduced during the rainy season (i.e., breeding period). During the breeding period, bird species would reduce participation in mixed-species flocks by prioritizing parental care; however, bird participation would increase during the dry and the flood seasons once mixed-species flocks facilitate the access to food resources (e.g., hidden insects) and the protection against predators (e.g., snakes) within the earthmound fields.
METHODS
Study area
This study was conducted in earthmound fields of the Brazilian Pantanal during the dry, rainy, and flood seasons of 2019 and also the flood season of 2020. The study area was located around the Retiro Novo cattle ranch, Mato Grosso, Brazil (16° 22′ 0.02″ S, 56° 18′ 0.00″ W), which is influenced by the annual flood-pulse of the Cuiabá and Piraim rivers. The size of the study area was approximately 2.8 km x 2.8 km, whereas the plots within the study area were approximately 600 m x 600 m. This region is characterized by a dry season from May to September, a rainy season from October to April, and also a brief flood season from January to March. Although the rainy season encompasses the flood season, we treated them as different events because the flood pulse generates an environmental change on open habitats that, subsequently, may influence the bird fauna. The mean annual precipitation is 1400 mm, with January and July as the wettest and driest months, respectively. The mean annual temperature is 25.8° C, ranging monthly, on average, from 20.9 to 32.5° C (Nunes da Cunha and Junk 2004, Junk et al. 2006).
The area is distinguished by its natural mosaic of open grassy-shrubby vegetation interspersed with patchy forest (Junk et al. 2006, Nunes da Cunha et al. 2015). The shrubby vegetation is dominated by Curatella americana, which grows on non-flooded termite mounds, which shape small-rounded (1-15 m²) dry islets (0.8-10 m high) imbedded into the grassy vegetation (da Silva et al. 2000, Nunes da Cunha and Junk 2004, Nunes da Cunha et al. 2006, 2007). The natural patchy forests comprise both seasonally flooded, evergreen environments (Landi forest) and non-flooding, semi-deciduous dry environments (Cordilheira forest). The Landi forest has a relative dense canopy and understory layer, varying from 3-15 m high, associated with wet soil conditions with negative relief that facilitate water drainage, and dominated by Alchornea discolor, Licania parviflora Huber, and Calophyllum brasiliensis. The Cordilheira forest has a relatively dense canopy and understory layer, varying from 8-20 m high (with emergent trees of 30 m), represented by semi-deciduous species such as Adelia membranifolia and Petiveria alliacea var. tetrandra, which increase the temperature and light exposition into the forest interior during the dry season. The understory in the Cordilheira forest is dominated by Bromelia balansae (da Silva et al. 2000, Nunes da Cunha et al. 2007, de Pinho and Marini 2014, Marques et al. 2016). Altogether, this vegetation is also connected across the landscape, inserted within a matrix of grasslands represented by Coleataenia stenodes, Axonopus purpusii, Anthenantia lanata, Andropogon selloanus, Mesosetum ansatum, and, occasionally, by the non-native Urochloa dictyoneura (Schessl 1999), creating a grassland-savanna-forest mosaic that, together with seasonality, may influence species participation on mixed-species flocks.
Mixed-species flock observations
Our study area was composed of four plots of 600 m² separated from each other by a minimum distance of 1 km (Fig. 1). We searched for mixed-species flocks within each plot and collected data on species composition in 30-minute observation periods, which ultimately were our sampling units (Goodale et al. 2009, Rutt and Stouffer 2021). Mixed-species flocks (hereafter “flocks”) were defined as a group of individuals of two or more species that forage together in a given space and time (Goodale et al. 2009). When following flocks, we collected data on the number of species and individuals, individuals per species, sex, and age (i.e., adults and young as a proxy for breeding) by direct observation with binoculars. Subsequently, we established if the flocking species were residents and/or migrants in the Pantanal by checking the literature (Nunes et al. 2021). Only birds within a distance of 10 m in relation to each other were considered to be part of the flock (Jullien and Thiollay 1998). Thus, flocks were followed by one experienced observer for one continuous hour during the morning, keeping behind flocks at a distance of 10-15 m. The observations of one hour per flock were divided into 2 blocks of 30-minute periods. Even if we had two repeated observations per flock, most individual flocks were different per season and across the year (Appendix 1). Before data collection, we randomly chose our study plot and, after its selection, we revisited the same plot for two or three consecutive days. Each plot was composed of a dominant matrix of grassy and shrubby vegetation interspersed with patchy forest. Flocks form within the grassy-shrubby vegetation, but also move along the forest edges and defend a stable territory throughout the day and year. By following one flock with a color-banded individual of Polioptila dumicola, we knew that flocks move in the same area, alternating their trajectory across days and throughout seasons. Unbanded flocks that were followed for one consecutive hour also gave us a good representation on space use. By additionally following flocks for two or three continuous hours across days and seasons, we were confident that flocks establish (G. J. Fernández-Arellano, J. Bonanomi, J. B. Pinho, unpublished data), defend, and move within their territory. Antagonistic events also gave us an idea on territorial boundaries and neighboring flocks. Sampling periods lasted 15 days in each season (dry: July, rainy: October, and flood season: late February-early March), giving a total of 40 blocks of observation periods during 20 h of flock-following only in 2019. During the flood season, grasslands are covered by water, from 0.5 to 1 m high, whereas the shrubby vegetation, dominated by C. americana, lies on mounds of earth that rise above of the water level. Thus, we walked slowly through the flooded open grassy vegetation while following flocks.
Data analysis
The mean number of species and mean number of individuals in flocks were compared among seasons, and we also tested for their relationship (i.e., the flocking richness and flocking size relationship). We also calculated and compared the species diversity in flocks among seasons by using the Shannon index (H), which considers the number of species and individuals, and also similarly considers rare and common species (Shannon 1948, Jost 2007). Additionally, we calculated sample-based species accumulation curves to assess whether the number of species in flocks was completely sampled by looking within and among seasons (McCune et al. 2002). The frequency of species attendance was calculated as a percentage that results from dividing the number of 30-minute periods of flock observations in which 1 species was observed by the total number of 30-minute periods of flock observations (Jullien and Thiollay 1998). Species attendance was calculated per species and season, but a mean value was also calculated across the seasons (e.g., 97.7% for Polioptila dumicola; see Appendix 2). Flocking structure (i.e., number of species, number of individuals and diversity) analysis was made by using only the observation data of 2019, while flocking organization description (i.e., frequency of species attendance) was made by using the observation data of 2019 and 2020 (the flood season).
Our data for flocks’ composition were organized into a matrix with rows representing flocks (the total number of 30-minute observation periods) and columns representing species. The matrix was filled with the number of individuals per species. By using our matrix, we tested for the influence of the seasonality (dry, rainy, and flood) on the number of species, individuals, and species diversity in flocks by fitting generalized linear mixed models (GLMMs). Thus, seasonality was included as a fixed factor, whereas flock and plot identity were added as random factors. Because the 30-minute periods come from the same flocks (16 individual flocks), we included flock nested within the plot in our model. Thus, we included a statistical control of these sources of variation to deal, as much as possible, with pseudoreplication. From the 16 individual flocks sampled during 2019, 12 flocks were sampled in 1 season and 4 flocks were sampled in 2 seasons (Appendix 1). The number of species and the number of individuals in flocks followed a Poisson error distribution, while species diversity followed a normal distribution. We used a log link function for the number of species and individuals, and an identity link for diversity. Multiple pairwise comparisons of means (Tukey’s post-hoc test) were performed to test for significant differences among the three levels of the fixed factor (i.e., seasonality: dry, rainy, and flood). All the analyses were performed in R v3.6.1 (R Core Team 2019), using the “lme4” (Bates et al. 2015) and the “multcomp” (Hothorn et al. 2008) packages. The sample-based species accumulation curves were based on a matrix combining data from 2019 and 2020 and the analysis was performed using the “vegan” package (Oksanen et al. 2019).
We performed a non-parametric Spearman correlation test to analyze the relationship between the number of species and individuals in flocks. Only the first 30-minute observation period per flock from 2019 was used in this analysis. We used the “vegan” package (Oksanen et al. 2019) to perform these analyses. Moreover, we used the mean frequency of species attendance to create four categories of species participation (organization; see Appendix 2) in flocks, following Jullien and Thiollay (1998): regular participants (> 75% of frequency of attendance), common participants (50-75%), uncommon participants (25-50%), and irregular participants (< 25%).
RESULTS
Organization of mixed-species flocks
We identified 69 species within 20 families, in 16 individual flocks sampled during 2019 and 2020 (Appendix 2). The mean frequency of species attendance through the three seasons showed that Polioptila dumicola (97.7%) was the most regular participant in flocks, followed by Myiarchus tyrannulus (73.6%) and Conirostrum speciosum (61.4%) as common participants. Moreover, six species (Lepidocolaptes angustirostris, Casiornis rufus, Hemitriccus margaritaceiventer, Camptostoma obsoletum, Myiopagis gaimardii, and Picumnus albosquamatus) were uncommon participants (29-40%). Conversely, 52 species were irregular participants (< 25%), with 11 species (Coryphospingus cuculatus, Veniliornis passerinus, Pachyramphus polychopterus, P. viridis, Synalaxis albilora, Nemosia pileata, Icterus cayannensis, Thamnophilus pelzelni, Xiphorhynchus picus, Galbula ruficauda, and Formicivora rufa) being even less irregular (10-21%) than the remaining 41 species (< 10%; Appendix 2).
Resident species were the main components of flocks across seasons, and the frequent species were local residents from the Pantanal (Appendix 2). A few migratory species were also participants in flocks. Some flycatcher species like Elaenia spp. were irregular participants of flocks during the rainy season, whereas Inezia inornata and Sublegatus modestus were irregular participants of flocks during the dry season. Moreover, tanagers (e.g., Volatinia jacarina) and tityrids (e.g., Tityra cayana) were irregular participants of flocks in the rainy and flood seasons, respectively.
Seasonal variation in the structure of mixed-species flocks
Seasonality had a significant effect on the mean number of species in flocks (χ22 = 26.03, P < 0.001). The mean number of species in the dry season was significantly higher than in the flood and rainy seasons, but the flood season was also significantly higher than the rainy season (Fig. 2). Seasonality also had a significant effect on the mean number of individuals in flocks (χ22 = 35.74, P < 0.001). That is, the mean number of individuals in the dry season was significantly higher than in the flood and rainy seasons, but the flood season was also significantly higher than the rainy season (Fig. 3). The mean number of species and individuals in flocks showed a significant positive correlation (Spearman rank correlation; r = 0.925, P < 0.001; see Appendix 3). Following the patterns for the mean number of species and individuals, seasonality also had a significant effect on species diversity in flocks (χ22 = 17.38, P < 0.001). Thus, species diversity in the dry and the flood seasons was similar, but also significantly higher than in the rainy season (Fig. 4). Moreover, the number of species in flocks per season seem to approach the asymptote as sampling effort increases (Appendix 4).
DISCUSSION
Our results show that the structure of mixed-species bird flocks in the Pantanal, a seasonally flooded tropical wetland, varies with seasonality. Specifically, flocks had fewer species, individuals, and diversity during the rainy season. These changes in flocking structure can be related to the interplay of several season-dependent processes, such as the timing of the breeding period (mostly during the rainy season in the Pantanal), resources availability, and/or landscape structure. Breeding period may constrain the participation of species in flocks, which consequently reduces flocking complexity (e.g., the number of species and individuals; Develey and Peres 2000). Similar results were observed in flocks from tropical savannas of Africa, Australia, and South America (Greig-Smith 1976, Alves and Cavalcanti 1992, Vanderduys et al. 2012) during the breeding period, which is commonly related with food resource availability (Stutchbury and Morton 2001). Otherwise, the complex flocking structure in the dry season seems to respond to the scarcity in food availability, when species may join flocks to increase their chances to access food resources by complementary foraging benefits (Develey and Peres 2000, Sridhar et al. 2009, Goodale et al. 2020). Likewise, differences in landscape configuration can also influence the flocking structure (Rutt and Stouffer 2021). When grasslands are covered by water during the flood season, species may join flocks to reduce their predation risk when moving through the sparse and non-flooded shrubby vegetation. For example, sentinel species can alert other flock members of predator presence (e.g., snakes) on those shrubby islets. Our results suggest that lower flocking complexity during the rainy season may be associated with parental care investment, whereas the higher complexity during the dry and flood seasons may reflect temporary benefits for species, such as a higher foraging success and predator avoidance, respectively.
Participation in flocks is regularly explained by two non-exclusive hypotheses, foraging and/or antipredator benefits (Morse 1977, Greenberg 2000, Goodale et al. 2020). Species can increase their foraging efficiency by simultaneously decreasing their own vigilance (Sridhar et al. 2009) and catching insects flushed by other species in flocks (Satischandra et al. 2007). For example, C. rufus can move close and below the other flock members to probably catch flushed arthropods. The scarcity of food resources during the dry season (e.g., the lower abundance of arthropods; Junk et al. 2006, Wantzen et al. 2016) may drive individuals to join flocks to access food resources by exploring the forest edges or by avoiding higher foraged areas (Sasvari and Hegyi 1998). In this regard, we observed color-banded individuals of L. angustirostris switching among flocks to probably exploit food resources within their large home range (Alves and Cavalcante 1996). Thus, flocks may facilitate the movement of individuals between open habitats and forests (Tubelis et al. 2006) and within forests (Jullien and Thiollay 1998). Simultaneously, individuals may join flocks to decrease their risk of predation (Jullien and Thiollay 1998, Sridhar et al. 2009) by using the early alarm calls of the sentinel species warning of predator presence (Munn 1986, Goodale and Kotagama 2008, Martínez and Zenil 2012). We observed, for example, one predation attempt of the large-sized hawk Chondrohierax uncinatus ambushing a C. rufus individual in the Landi forest edge, and two mobbing events against the owl species Megascops choliba and Glaucidium brasilianum at the edges of the Landi and Cordilheira forests, respectively (G. Fernández-Arellano; personal observation), during the flood season. Although we did not evaluate the foraging and the antipredator benefits in flocking species, it is feasible to think that the access to food resources and protection against predators may increase individuals’ participation in flocks during the dry and flood seasons, respectively.
Different from the dry and flood seasons, the rainy season may constrain individuals to join flocks (Alves and Cavalcanti 1996, Tubelis et al. 2006, but see Vanderduys et al. 2012 in Australian savannas) once an individual prioritizes breeding activities during this season, resulting in noticeably smaller flocks. This is a common response observed in flocks from tropical ecosystems, including savannas and temperate forests (Morse 1977, Greenberg 2000, Tubelis 2007). Prioritizing breeding activities during the rainy (October to April) season entails higher foraging of food resources for nestlings and/or fledglings’ growth, thus reducing their mortality rates during the dry (May to September) and the flood (January to March) seasons. Flocking structure can be positively influenced by the arrival of migratory species (e.g., Elaenia spp.) throughout the year but negatively influenced by those breeding resident species (e.g., P. murina) during the rainy season in the Pantanal. Overall, species start to breed almost at the same time, thus flocking structure seems unlikely to be influenced by the breeding of specific species (Alves and Cavalcanit 1996, Tubelis et al. 2006). For instance, Rutt and Stouffer (2021) reported that breeding nuclear species (e.g., Thamnomanes caesius) influence flocking behavior by reducing species participation and movement. Flocks from savannas show a similar response (Alves and Cavalcanti 1996, Tubelis et al. 2006). In the Pantanal, P. dumicola, the nuclear species, was present in flocks even when breeding outside and during the rainy season (de Pinho and Marini 2014; G. Fernández-Arellano, personal observation). The breeding period of songbirds in the Pantanal encompasses both the dry and rainy seasons (de Pinho and Marini 2014), but breeding reaches its peak during the first rains (de Pinho et al. 2006, Rubio and de Pinho 2008, de Nóbrega and de Pinho 2010, Bernardon et al. 2014) when flock complexity is reduced in relation to the dry and flood seasons. However, the participation in flocks during the rainy season suggests that species still derive some benefits (e.g., lower interspecific competition for food resources). Our results show that, like other savanna-like tropical systems, seasonality associated with breeding may influence the species’ participation in flocks in the Pantanal wetland.
The participation in flocks of multiple species is a regular feature across biogeographic regions but reaches greater complexity in the tropics (Greenberg 2000, Zou et al. 2018). Flocks of insectivorous birds are notable examples of mutualistic associations occurring in the Amazon rainforest (Munn and Terborgh 1979, Martinez and Gomez 2013) and in savanna-like ecosystems (Greig-Smith 1976, Vanderduys et al. 2012). In the Pantanal, nine species were more frequently in flocks, but only P. dumicola seemed to be a key species for the formation, cohesion, and direction of flocks because of its active foraging (e.g., up and down fantail balance) and displacement into new areas, which may facilitate the arrival of other flock members. Likewise, the conspicuous vocalization (e.g., frequent contact, short acute calls) and coloration (grayish-white body) of P. dumicola may induce the other species to follow it. Thus, P. dumicola shows the typical behavior of nuclear species in flocks (Moynihan 1962). In agreement with our results, Hutto (1994) and Gram (1998) suggested that the congeneric P. caerulea, a migratory species, plays an active nuclear role in flocks of a semi-deciduous dry forest in Mexico. Similar active behavior has been observed for other nuclear species such as the flycatcher Suiriri suiriri in our study area (de Vasconcelos et al. 2008), and in the southern Pantanal (Amaral and Ragusa-Netto 2008) during the dry season. However, we suggest that C. rufus, a flycatcher species, acts as a sentinel in flocks, however we have still not observed any situations involving alarm calls. In this regard, C. rufus shows a sit-and-wait from a perch behavior, sometimes below other members, as observed in sentinel species from Amazonian flocks (Thamnomanes schistogynus; Munn 1986), which probably helps it to catch arthropods flushed by other flock members. Further observations would be required to make more solid conclusions about the role this species plays in flocks. Contrary to our results, flocks on adjacent savannas (Cerrado biome) were composed by mid-sized gregarious tanagers as sentinel species (Neothraupis fasciata; Alves and Cavalcanti 1996, Cypsnagra hirundinacea; Ragusa-Netto 2000). Variation in species foraging behavior (e.g., myopic gleaners) may reflect differences in the degree of participation and the role of species in flocks, which may ultimately influence the flocking structure and organization.
In the Pantanal, most flocking bird species mainly inhabit the open earthmound fields, so the increasing and unceasing loss of this habitat may largely affect bird species assemblage. Still, the potential impact of these landscape changes on birds from open habitats of the Pantanal has been seriously neglected. The rapid and pervasive landscape conversion by plant species invasion, farming and crops, extreme droughts, and/or human-induced fires reduces bird community diversity in tropical savannas (Sirami et al. 2009, Woinarski and Legge 2013, McCleery et al. 2018). Studying flocks, as a set of interacting species and good ecological indicators of a bird community, can be relevant to understand the influence of the anthropogenic environmental change on the dynamic, functioning service and structure of mutualistic interactions within bird communities (Goodale et al. 2010, 2015). Hence, taking the interaction of habitat quality and seasonality together would ultimately help us to understand the dynamics of flocking species in space and time.
We showed the first description of the temporal variation of the structure and organization of flocks in a grassy-shrubby-forest vegetation gradient in the Pantanal, which were strongly influenced by seasonality, similarly to other savannas. The breeding period associated with the rainy season may negatively influence the flocking structure, whereas the access to food resources and protection may positively influence the flocking structure during the dry and the flood seasons. The study of flocks is needed to fully understand how mutualisms among bird species are organized and how they vary in seasonal and heterogeneous environments. We call for future research to examine the species’ role and the effects of human-induced environmental changes on the temporal and spatial dynamics of mixed-species flocks in the Pantanal, the most bird-rich wetland in the world, undergoing accelerating rates of fragmentation and native habitat loss.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
We thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for granting a PhD scholarship to GJFA and the National Institute of Science and Technology in Wetlands (INAU) for logistic support. We also thank Yasmin Columbano, Bruna Amaral, and Thayna Gomes for fieldwork assistance and Alberto L. Teixido for manuscript review. We confirm that there are no known conflicts of interest associated with this publication.
DATA AVAILABILITY
The data/codes are available here: https://doi.org/10.6084/m9.figshare.23642157.v1
LITERATURE CITED
Alho, C. J. R. 2008. Biodiversity of the Pantanal: response to seasonal flooding regime and to environmental degradation. Brazilian Journal of Biology 68(4):957-966. https://doi.org/10.1590/S1519-69842008000500005
Alho, C. J. R., S. B. Mamede, M. Benites, B. S. Andrade, and J. J. O. Sepúlveda. 2019. Threats to the biodiversity of the Brazilian Pantanal due to land use and occupation. Ambiente and Sociedade 22:e01891. https://doi.org/10.1590/1809-4422asoc201701891vu2019l3ao
Alves, M. A. S., and R. S. Cavalcante. 1996. Sentinel behavior, seasonality, and the structure of biro flocks in a Brazilian savanna. Ornitologia Neotropical 7:43-51.
Amaral, P. P., and J. Ragusa-Netto. 2008. Bird mixed-flocks and nuclear species in a tecoma savanna in the Pantanal. Brazilian Journal of Biology 68(3):511-518. https://doi.org/10.1590/S1519-69842008000300007
Andersen, E. M., and R. J. Steidl. 2019. Woody plant encroachment restructures bird communities in semiarid grasslands. Biological Conservation 240:108276. https://doi.org/10.1016/j.biocon.2019.108276
Barbosa da Silva, F. H., J. Arieira, P. Parolin, C. Nunes da Cunha, and W. J. Junk. 2016. Shrub encroachment influences herbaceous communities in flooded grasslands of a neotropical savanna wetland. Applied Vegetation Science 19(3):391-400. https://doi.org/10.1111/avsc.12230
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1):1-48. https://doi.org/10.18637/jss.v067.i01
Bell, H. L. 1983. A bird community of lowland rainforest in New Guinea. 5. Mixed-species feeding flocks. Emu - Austral Ornithology 82:256-275. https://doi.org/10.1071/MU9820256s
Bernardon, B., P. F. A. de Nóbrega, and J. B. de Pinho. 2014. Reproductive biology and nest-site selection of the Mato Grosso Antbird Cercomacra melanaria in the Brazilian Pantana. Revista Brasileira de Ornitologia 22(3):270-277. https://doi.org/10.1007/BF03544258
da Silva, M. P., R. Mauro, G. Mourão, and M. Coutinho. 2000. Distribuição e quantificação de classes de vegetação do Pantanal através de levantamento aéreo. Brazilian Journal of Botany 23:143-152. https://doi.org/10.1590/S0100-84042000000200004
de Nóbrega, P. F. A., and J. B. de Pinho. 2010. Biologia reprodutiva e uso de habitat por Cantorchilus leucotis (Lafresnaye, 1845) (Aves, Troglodytidae) no pantanal, Mato Grosso, Brasil. Papéis Avulsos de Zoologia 50(31):511-516. https://doi.org/10.1590/S0031-10492010003100001
de Pinho, J. B., M. Aragona, K. Y. P. Hakamada, and M. Â. Marini. 2017. Migration patterns and seasonal forest use by birds in the Brazilian Pantanal. Bird Conservation International 27(3):371-387. https://doi.org/10.1017/S0959270916000290
de Pinho, J. B., L. E. Lopes, D. H. de Morais, and A. M. Fernandes. 2006. Life history of the Mato Grosso Antbird Cercomacra melanaria in the Brazilian Pantanal. Ibis 148(2):321-329. https://doi.org/10.1111/j.1474-919X.2006.00542.x
de Pinho, J. B., and M. Â. Marini. 2014. Birds nesting parameters in four forest types in the Pantanal wetland. Brazilian Journal of Biology 74(4):890-898. https://doi.org/10.1590/1519-6984.08713
de Vasconcelos, M. F., L. E. Lopes, D. Hoffmann, L. F. Silveira, and F. Schunck. 2008. Noteworthy records of birds from the Pantanal, Chiquitano dry forest and Cerrado of south-western Brazil. Bulletin of the British Ornithologists’ Club 128(1):57-67.
Develey, P. F., and C. A. Peres. 2000. Resource seasonality and the structure of mixed species bird flocks in a coastal Atlantic forest of southeastern Brazil. Journal of Tropical Ecology, 16(1), 33-53. https://doi.org/10.1017/S0266467400001255
Dorado‐Rodrigues, T. F., V. M. G. Layme, F. H. B. Silva, C. Nunes da Cunha, and C. Strüssmann. 2015. Effects of shrub encroachment on the anuran community in periodically flooded grasslands of the largest Neotropical wetland. Austral Ecology 40(5):547-557. https://doi.org/10.1111/aec.12222
Fernández‐Arellano, G. J., A. L. Teixido, B. Bernardon, E. R. Bueno, T. V. Ferreira, S. R. Gonçalves, M. Jesus, K. P. C. S. Thomas, M. Zucchetto, V. Q. Piacentini, and J. B. Pinho. 2021. Knowledge gaps and biases in the Pantanal indicate future directions for ornithological research in large wetlands. Ibis 163(3):784-797. https://doi.org/10.1111/ibi.12931
Garcia, L. C., J. K. Szabo, F. de Oliveira Roque, A. de. Matos Martin Pereira, C. Nunes da Cunha, G. A. Damasceno-Júnior, R. G. Morato, W. M. Tomas, R. Libonati, and D. B. Ribeiro. 2021. Record-breaking wildfires in the world’s largest continuous tropical wetland: integrative fire management is urgently needed for both biodiversity and humans. Journal of Environmental Management 293:112870. https://doi.org/10.1016/j.jenvman.2021.112870
Goodale, E., G. Beauchamp, R. D. Magrath, J. C. Nieh, and G. D. Ruxton. 2010. Interspecific information transfer influences animal community structure. Trends in Ecology and Evolution 25(6):354-361. https://doi.org/10.1016/j.tree.2010.01.002
Goodale, E., G. Beauchamp, and G. D. Ruxton. 2017. Mixed-species groups of animals: behavior, community structure, and conservation. Academic Press, London, UK.
Goodale, E., P. Ding, X. Liu, A. E. Martínez, X. Si, M. Walters, and S. K. Robinson. 2015. The structure of mixed-species bird flocks, and their response to anthropogenic disturbance, with special reference to East Asia. Avian Research 6(1):14. https://doi.org/10.1186/s40657-015-0023-0
Goodale, E., and S. W. Kotagama. 2008. Response to conspecific and heterospecific alarm calls in mixed-species bird flocks of a Sri Lankan rainforest. Behavioral Ecology 19(4):887-894. https://doi.org/10.1093/beheco/arn045
Goodale, E., B. Z. Nizam, V. V. Robin, H. Sridhar, P. Trivedi, S. W. Kotagama, U. K. G. K. Padmalal, R. Perera, P. Pramod, and L. Vijayan. 2009. Regional variation in the composition and structure of mixed-species bird flocks in the Western Ghats and Sri Lanka. Current Science 97(5):648-663.
Goodale, E., H. Sridhar, K. E. Sieving, P. Bangal, G. J. Colorado Z., D. R. Farine, E. W. Heymann, H. H. Jones, I. Krams, A. E. Martínez, F. Montaño-Centellas, J. Muñoz, U. Srinivasan, A. Theo., and K. Shanker. 2020. Mixed company: a framework for understanding the composition and organization of mixed‐species animal groups. Biological Reviews 95(4):889-910. https://doi.org/10.1111/brv.12591
Gram, W. K. 1998. Winter participation by Neotropical migrant and resident birds in mixed-species flocks in northeastern Mexico. Condor 100(1):44-53. https://doi.org/10.2307/1369895
Greenberg, R. S. 2000. Birds of many feathers: the formation and structure of mixed species flocks of forest birds. Pages 521-559 in S. Boinski and P. Garger. On the move: how and why animals travel in groups. University of Chicago Press, Chicago, Illinois, USA.
Greig‐Smith, P. W. 1978. The formation, structure and function of mixed‐species insectivorous bird flocks in West African savanna woodland. Ibis 120(3):284-297. https://doi.org/10.1111/j.1474-919X.1978.tb06789.x
Gwynne, J. A., R. S. Ridgely, G. Tudor, and M. Argel. 2010. Aves do Brasil: Pantanal e Cerrado. Editora Horizonte, Sao Paulo, Brasil.
Hothorn, T., F. Bretz, and P. Westfall. 2008. Simultaneous inference in general parametric models. Biometrical Journal 50(3):346-363. https://doi.org/10.1002/bimj.200810425
Hutto, R. L. 1994. The composition and social organization of mixed-species flocks in a tropical deciduous forest in western Mexico. Condor 96(1):105-118. https://doi.org/10.2307/1369068
Jayarathna, A., S. W. Kotagama, and E. Goodale. 2013. The seasonality of mixed-species bird flocks in a Sri Lankan rainforest in relation to the breeding of the nuclear species, Orange-billed Babbler Turdoides rufescens. Forktail 29:138-139.
Jost, L. 2007. Partitioning diversity into independent alpha and beta components. Ecology 88(10):2427-2439. https://doi.org/10.1890/06-1736.1
Jullien, M., and J.-M. Thiollay. 1998. Multi‐species territoriality and dynamic of neotropical forest understorey bird flocks. Journal of Animal Ecology 67(2):227-252. https://doi.org/10.1046/j.1365-2656.1998.00171.x
Junk, W. J., C. Nunes da Cunha, K. M. Wantzen, P. Petermann, C. Strüssmann, M. I. Marques, and J. Adis. 2006. Biodiversity and its conservation in the Pantanal of Mato Grosso, Brazil. Aquatic Sciences 68(3):278-309. https://doi.org/10.1007/s00027-006-0851-4
Lees, A. C., K. V. Rosenberg, V. Ruiz-Gutierrez, S. Marsden, T. S. Schulenberg, and A. D. Rodewald. 2020. A roadmap to identifying and filling shortfalls in Neotropical ornithology. Auk 137(4):ukaa048. https://doi.org/10.1093/auk/ukaa048
Macedo, R. H. F. 2002. The avifauna: ecology, biogeography, and behavior. Pages 242-266 in P. S. Oliveira and R. J. Marquis, editors. The cerrados of Brazil: ecology and natural history of a neotropical savanna. Columbia University Press, New York, New York, USA. https://doi.org/10.7312/oliv12042-012
Marengo, J. A., L. M. Alves, and R. R. Torres. 2016. Regional climate change scenarios in the Brazilian Pantanal watershed. Climate Research 68(2-3):201-213. https://doi.org/10.3354/cr01324
Marques, M. I., A. M. de Figueiredo, G. B. dos Santos, W. O. Sousa, S. Ide, and L. D. Battirola. 2016. Diversity of soil beetles (Hexapoda, Coleoptera) in an area at the Pantanal of Poconé, Mato Grosso, Brazil. EntomoBrasilis 9(2):89-96. https://doi.org/10.12741/ebrasilis.v9i2.539
Martínez, A. E., and J. P. Gomez. 2013. Are mixed-species bird flocks stable through two decades? American Naturalist 181(3):E53-E59. https://doi.org/10.1086/669152
Martínez, A. E., and R. T. Zenil. 2012. Foraging guild influences dependence on heterospecific alarm calls in Amazonian bird flocks. Behavioral Ecology 23(3):544-550. https://doi.org/10.1093/beheco/arr222
Mccleery, R., A. Monadjem, B. Baiser, R. Fletcher, Jr., K. Vickers, and L. Kruger. 2018. Animal diversity declines with broad-scale homogenization of canopy cover in African savannas. Biological Conservation 226:54-62. https://doi.org/10.1016/j.biocon.2018.07.020
Mccune, B., J. B. Grace, and D. L. Urban. 2002. Analysis of ecological communities. MjM software design, Gleneden Beach, Oregon, USA.
Morse, D. H. 1977. Feeding behavior and predator avoidance in heterospecific groups. BioScience 27(5):332-339. https://doi.org/10.2307/1297632
Moynihan, M. H. 1962. The organization and probable evolution of some mixed species flocks of neotropical birds. Smithsonian Miscellaneous Collections, Smithsonian Institute, Washington, D.C., USA.
Munn, C. A. 1985. Permanent canopy and understory flocks in Amazonia: species composition and population density. Ornithological Monographs 36:683-712. https://doi.org/10.2307/40168312
Munn, C. A. 1986. Birds that ‘cry wolf’. Nature 319(6049):143-145. https://doi.org/10.1038/319143a0
Munn, C. A., and J. W. Terborgh. 1979. Multi-species territoriality in Neotropical foraging flocks. Condor 81(4):338-347. https://doi.org/10.2307/1366956
Nunes, A. P., S. R. Posso, A. V. B. da Frota, B. D. Vitorino, R. R. Laps, R. J. Donatelli, F. C. Straube, M. A. C. Pivatto, D. M. M. de Oliveira, B. Carlos, A. V. de Melo, W. M. Tomas, G. O. de Freitas, R. A. D. de Souza, M. Benites, S. Mamede, and R. S. Moreira. 2021. Birds of the Pantanal floodplains, Brazil: historical data, diversity, and conservation. Papéis Avulsos de Zoologia 61:e20216182. https://doi.org/10.11606/1807-0205/2021.61.82
Nunes da Cunha, C., and W. J. Junk. 2004. Year-to-year changes in water level drive the invasion of Vochysia divergens in Pantanal grasslands. Applied Vegetation Science 7(1):103-110. https://doi.org/10.1111/j.1654-109X.2004.tb00600.x
Nunes da Cunha, C., W. J. Junk, and H. F. Leitão-Filho. 2007. Woody vegetation in the Pantanal of Mato Grosso, Brazil: a preliminary typology. Amazoniana 19(3/4):159-184. https://pure.mpg.de/rest/items/item_1506673_1/component/file_1506672/content
Nunes da Cunha, C., M. T. F. Piedade, and W. J. Junk. 2015. Classificação e delineamento das áreas úmidas brasileiras e de seus macrohabitats. EdUFMT, Cuiabá, Brasil. http://cppantanal.org.br/wp-content/uploads/2017/04/E-book-Classificacao-e-Delineamento-das-AUs.pdf
Nunes da Cunha, C., P. Rawiel, K. M. Wantzen, W. J. Junk, and A. Lemes do Prado. 2006. Mapping and characterization of vegetation units by means of Landsat imagery and management recommendations for the Pantanal of Mato Grosso (Brazil), north of Poconé. Amazoniana 19(1/2):1-32. https://pure.mpg.de/rest/items/item_1506769_1/component/file_1506768/content
Oksanen, F. J., F. G. Blanchet, and M. Friendly. 2019. vegan: community ecology package. R package version 2.5-5. R Foundation for Statistical Computing, Vienna, Austria. https://CRAN.R-project.org/package=vegan
Pletsch, M. A. J. S., C. H. L. Silva, Jr., T. V. Penha, T. S. Körting, M. E. S. Silva, G. Pereira, L. O. Anderson, and L. E. O. C. Aragão. 2021. The 2020 Brazilian Pantanal fires. Anais da Academia Brasileira de Ciências 93(3):e20210077. https://doi.org/10.1590/0001-3765202120210077
Powell, G. V. 1985. Sociobiology and adaptive significance of interspecific foraging flocks in the Neotropics. Ornithological Monographs 36:13-732. https://doi.org/10.2307/40168313
Ragusa-Netto, J. 2000. Raptors and “campo-cerrado” bird mixed flock led by Cypsnagra hirundinacea (Emberizidae: Thraupinae). Revista Brasileira de Biologia 60(3):461-467. https://doi.org/10.1590/S0034-71082000000300011
R Development Core Team. 2019. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/
Rubio, T. C., and J. B. de Pinho. 2008. Biologia reprodutiva de Synallaxis albilora (Aves: Furnariidae) no Pantanal de Poconé, Mato Grosso. Papéis Avulsos de Zoologia 48(17):181-197. https://doi.org/10.1590/S0031-10492008001700001
Rutt, C. L., and P. C. Stouffer. 2021. Seasonal dynamics of flock interaction networks across a human‐modified landscape in lowland Amazonian rain forest. Ecological Applications 31(2):e02235. https://doi.org/10.1002/eap.2235
Sasvári, L., and Z. Hegyi. 1998. How mixed-species foraging flocks develop in response to benefits from observational learning. Animal Behaviour 55(6):1461-1469. https://doi.org/10.1006/anbe.1998.0734
Satischandra, S. H. K., E. P. Kudavidanage, S. W. Kotagama, and E. Goodale. 2007. The benefits of joining mixed-species flocks for greater racket-tailed drongos Dicrurus paradiseus. Forktail 23:145-148.
Schessl, M. 1999. Floristic composition and structure of floodplain vegetation in the northern Pantanal of Mato Grosso, Brazil. Phyton (Horn, Austria) 39(2):303-336. https://www.zobodat.at/pdf/PHY_39_2_0303-0336.pdf
Shannon, C. E. 1948. A mathematical theory of communication. Bell System Technical Journal 27(3):379-423. https://people.math.harvard.edu/~ctm/home/text/others/shannon/entropy/entropy.pdf
Sick, H. 1997. Ornitologia Brasileira. Nova Fronteira SA, Rio de Janeiro, Brasil.
Sirami, C., C. Seymour, G. Midgley, and P. Barnard. 2009. The impact of shrub encroachment on savanna bird diversity from local to regional scale. Diversity and Distributions 15(6):948-957. https://doi.org/10.1111/j.1472-4642.2009.00612.x
Sridhar, H., G. Beauchamp, and K. Shanker. 2009. Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Animal Behaviour 78(2):337-347. https://doi.org/10.1016/j.anbehav.2009.05.008
Stanton, Jr., R. A., W. W. Boone, IV, J. Soto‐Shoender, R. J. Fletcher, Jr., N. Blaum, and R. A. Mccleery. 2018. Shrub encroachment and vertebrate diversity: a global meta‐analysis. Global Ecology and Biogeography 27(3):368-379. https://doi.org/10.1111/geb.12675
Stutchbury, B. J., and E. S. Morton. 2001. Behavioral ecology of tropical birds. Academic Press, San Diego, California, USA.
Thielen, D., K. L. Schuchmann, P. Ramoni-Perazzi, M. Marquez, W. Rojas, J. I. Quintero, and M. I. Marques. 2020. Quo vadis Pantanal? Expected precipitation extremes and drought dynamics from changing sea surface temperature. PLoS ONE 15(1):e0227437. https://doi.org/10.1371/journal.pone.0227437
Tomas, W. M., C. N. Berlinck, R. M. Chiaravalloti, G. P. Faggioni, C. Strüssmann, R. Libonati, C. R. Abrahĕo, G. do Valle Alvarenga, A. E. de Faria Bacellar, F. R. de Queiroz Batista, T. S. Bornato, A. R. Camilo, J. Castedo, A. M. E. Fernando, G. O. de Freitas, C. M. Garcia, H. S. Gonçalves, M. B. de Freitas Guilherme, V. M. G. Layme, A. P. G. Lustosa, A. C. De Oliveira, M. da Rosa Oliveira, A. de Matos Martins Pereira, J. A. Rodrigues, T. B. F. Semedo, R. A. D. de Souza, F. R. Tortato, D. F. P. Viana, L. Vicente-Silva, and R. Morato. 2021. Distance sampling surveys reveal 17 million vertebrates directly killed by the 2020’s wildfires in the Pantanal, Brazil. Scientific Reports 11(1):23547. https://doi.org/10.1038/s41598-021-02844-5
Tomas, W. M., F. de Oliveira Roque, R. G. Morato, P. E. Medici, R. M. Chiaravalloti, F. R. Tortato, J. M. F. Penha, T. J. Izzo, L. C. Garcia, R. F. F. Lourival, P. Girard, N. R. Albuquerque, M. Almeida-Gomes, M. H. da Silva Andrade, F. A. S. Araujo, A. C. Araujo, E. C. de Arruda, V. A. Assunção, L. D. Battirola, M. Benites, F. P. Bolzan, J. C. Boock, I. M. Bortolotto, M. da Silva Brasil, A. R. Camilo, Z. Campos, M. A. Carniello, A. C. Catella, C. C. Cheida, P. G. Crawshaw, Jr., S. M. A. Crispim, G. A. D. Junior, A. L. J. Desbiez, F. A. Dias, D. P. Eaton, G. P. Faggioni, M. A. Farinaccio , J. F. A. Fernandes, V. L. Ferreira, E. A. Fischer, C. E. Fragoso, G. O. Freitas, F. Galvani, A. S. Garcia, C. M. Garcia, G. Graciolli, R. D. Guariento, N. M. R. Guedes, A. Guerra, H. M. Herrera, R. Hoogesteijn, S. C. Ikeda, R. S. Juliano, D. L. Z. K. Kantek, A. Keuroghlian, A. C. R. Lacerda, A. L. R. Lacerda, V. L. Landeiro, R. R. Laps, V. Layme, P. Leimgruber, F. L. Rocha, S. Mamede, D. K. S. Marques, M. I. Marques, L. A. F. Mateus, R. N. Moraes, T. A. Moreira, G. M. Mourĕo, R. D. Nicola, D. G. Nogueira, A. P. Nunes, C. da Nunes da Cunha, M. D. Oliveira, M. R. Oliveira, G. M. Paggi, A. O. Pellegrin, G. M. F. Pereira, I. A. H. F. S. Peres, J. B. Pinho, J. O. P. Pinto, A. Pott, D. B. Provete, V. D. A. dos Reis, L. K. dos Reis, P.-C. Renaud, D. B. Ribeiro, O. C. Rossetto, J. Sabino, D. Rumiz, S. M. Salis, D. J. Santana, S. A. Santos, Â. L. Sartori, M. Sato, K.-L. Schuchmann, E. Scremin-Dias, G. H. F. Seixas, F. Severo-Neto, M. R. Sigrist, A. Silva, C. J. Silva, A. L. Siqueira, B. M. A. Soriano, L. M. Sousa, F. L. Souza, C. Strussmann, L. S. M. Sugai, N. Tocantins, C. Urbanetz, F. Valente-Neto, D. P. Viana, A. Yanosky, and W. J. Junk. 2019. Sustainability agenda for the Pantanal Wetland: perspectives on a collaborative interface for science, policy, and decision-making. Tropical Conservation Science 12. https://doi.org/10.1177/1940082919872634
Tubelis, D. P. 2004. Species composition and seasonal occurrence of mixed-species flocks of forest birds in savannas in central Cerrado, Brazil. Ararajuba 12(2):105-111. http://www.revbrasilornitol.com.br/BJO/article/view/2603/pdf_295
Tubelis, D. P. 2007. Mixed-species flocks of birds in the Cerrado, South America: a review. Ornitologia Neotropical 18:75-97.
Tubelis, D. P., A. Cowling, and C. Donnelly. 2006. Role of mixed‐species flocks in the use of adjacent savannas by forest birds in the central Cerrado, Brazil. Austral Ecology 31(1):38-45. https://doi.org/10.1111/j.1442-9993.2006.01541.x
Tubelis, D. P., and W. M. Tomás. 1999. Distribution of birds in a naturally patchy forest environment in the Pantanal wetland, Brazil. Revista Brasileira de Ornitologia 7(11):81-89. http://www.revbrasilornitol.com.br/BJO/article/view/1104/pdf_180
Tubelis, D. P., and W. M. Tomás. 2003. Bird species of the Pantanal wetland, Brazil. Ararajuba 11(1):5-37. http://www.faunaparaguay.com/Tub11.pdf
Vanderduys, E. P., A. S. Kutt, J. J. Perry, and G. C. Perkins. 2012. The composition of mixed-species bird flocks in northern Australian savannas. Emu - Austral Ornithology 112(3):218-226. https://doi.org/10.1071/MU11041
Wantzen, K. M., M. R. Marchese, M. I. Marques, and L. D. Battirola. 2016. Invertebrates in neotropical floodplains. Pages 493-524 in Invertebrates in D. Batzer and D. Boix, editors. Invertebrates in freshwater wetlands. Springer International, Basel, Switzerland. https://doi.org/10.1007/978-3-319-24978-0_14
Woinarski, J. C. Z., and S. Legge. 2013. The impacts of fire on birds in Australia’s tropical savannas. Emu - Austral Ornithology 113(4):319-352. https://doi.org/10.1071/MU12109
Zou, F., H. Jones, G. J. Colorado Z., D. Jiang, T.-M. Lee, A. Martínez, K. Sieving, M. Zhang, Q. Zhang, and E. Goodale. 2018. The conservation implications of mixed-species flocking in terrestrial birds, a globally-distributed species interaction network. Biological Conservation 224:267-276. https://doi.org/10.1016/j.biocon.2018.06.004
Fig. 1
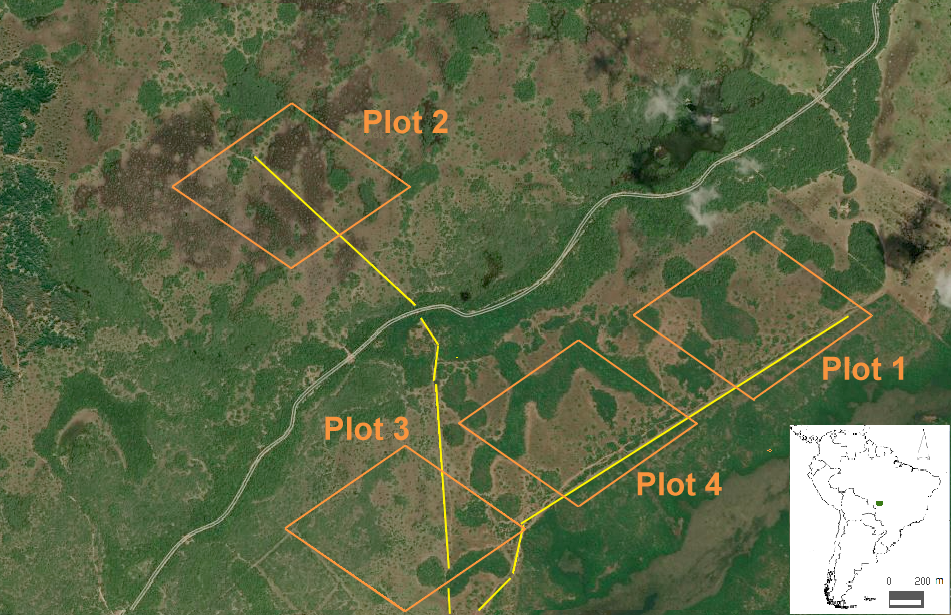
Fig. 1. Study area around the Retiro Novo cattle ranch, Poconé, Mato Grosso, Brazil. Plots encompassed the mosaic of patchy forest (dark and light green colors) and temporary flooded grassy-shrubby vegetation (light and dark brown but also the black color; earthmound fields). Gray color indicates a temporary drought watercourse. The orange boxes indicate each plot and the yellow lines indicate the main two trails.
Fig. 2
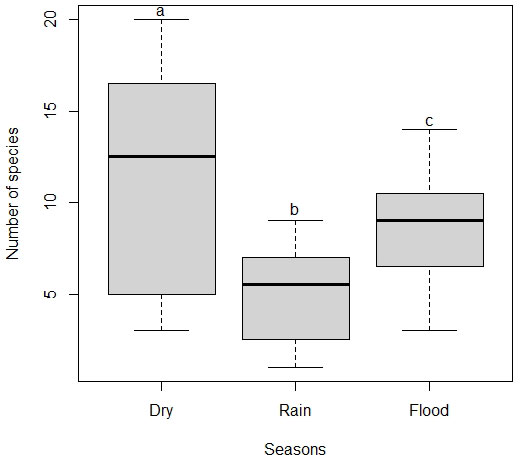
Fig. 2. Variation in the number of species in mixed-species flocks per season in earthmound fields (i.e., islets of scattered shrubby vegetation inserted into a temporally flooded matrix of grassy vegetation) of the Pantanal. Different letters indicate significant differences (P < 0.05).
Fig. 3
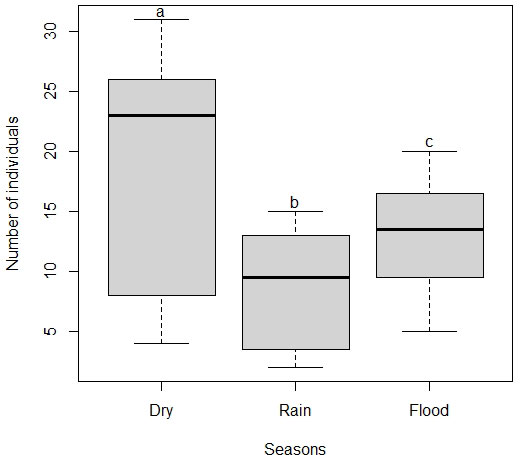
Fig. 3. Variation in the number of individuals in mixed-species flocks per season in earthmound fields (i.e., islets of scattered shrubby vegetation inserted into a temporally flooded matrix of grassy vegetation) of the Pantanal. Different letters indicate significant differences (P < 0.05).
Fig. 4
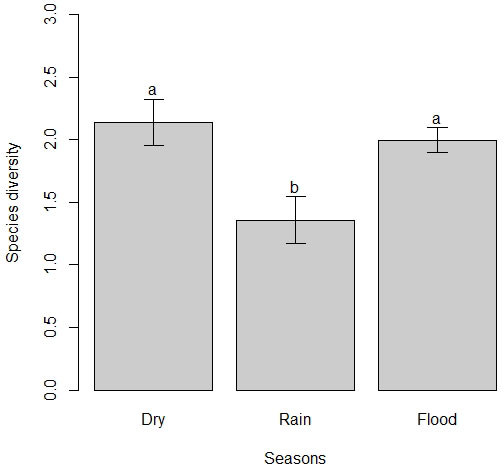
Fig. 4. Variation on species diversity in mixed-species flocks per season in earthmound fields (i.e., islets of scattered shrubby vegetation inserted into a temporally flooded matrix of grassy vegetation) of the Pantanal. Different letters indicate significant differences (P < 0.05).