







The following is the established format for referencing this article:
Ippi, S., B. Riovitti, V. Ferretti, V. S. Ojeda, M. Barrionuevo, A. A. Escalante, and M. A. Pacheco. 2026. Extremely low haemosporidian prevalence in a South American migratory aerial insectivore (Tachycineta leucopyga). Journal of Field Ornithology 97(2):7.ABSTRACT
Blood parasites can influence avian physiology, behavior, and reproduction, yet their prevalence and diversity vary across hosts, environments, and latitudes. We assessed haemosporidian infections in the migrant Chilean Swallow (Tachycineta leucopyga) at two widely separated breeding populations in Argentine Patagonia. Molecular screening of the parasite mitochondrial cytochrome b gene conducted on 209 individuals revealed a single Leucocytozoon-infected adult in Tierra del Fuego; no other infections were detected. Blast and phylogenetic analyses identified the parasite as belonging to the lineage DIUDIU11. This lineage has been previously reported in several South American passerine species and is closely related to Leucocytozoon fringillinarum. Our results revealed extremely low hemoparasite prevalence in this aerial insectivore, likely reflecting limited vector contact because of their behavior, low vector abundance, or the absence of infectious parasites at breeding and/or non-breeding sites. These findings establish a first baseline for hemoparasite dynamics in a long-term monitored South American swallow population and emphasize the importance of assessing migratory birds’ health across their full annual cycle under ongoing environmental change and climate warming.
RESUMEN
Los parásitos sanguíneos pueden influir en la fisiología, comportamiento y reproducción de las aves, aunque su prevalencia y diversidad varían entre hospedadores, ambientes y latitudes. Evaluamos infecciones por hemosporidios en la golondrina migratoria Tachycineta leucopyga en dos poblaciones reproductivas ampliamente separadas en la Patagonia argentina. El análisis de detección molecular del gen mitocondrial citocromo b del parásito, realizado sobre 209 individuos, reveló un único adulto infectado por Leucocytozoon en Tierra del Fuego; no se detectaron otras infecciones. Los análisis BLAST y filogenéticos identificaron al parásito como perteneciente al linaje DIUDIU11. Este linaje ha sido reportado previamente en varias especies de paseriformes sudamericanos y está estrechamente relacionado con Leucocytozoon fringillinarum. Nuestros resultados revelaron una prevalencia extremadamente baja de hemoparásitos en este insectívoro aéreo, reflejando probablemente un contacto limitado con los vectores debido a su comportamiento, la baja abundancia de vectores o la ausencia de parásitos infecciosos en los sitios reproductivos y/o no reproductivos. Estos hallazgos establecen una primera línea de base para la dinámica de hemoparásitos en una población sudamericana de golondrinas monitoreada a largo plazo y enfatizan la importancia de evaluar la salud de las aves migratorias a lo largo de todo su ciclo anual frente al actual cambio ambiental y calentamiento climático.
INTRODUCTION
Parasites are hypothesized to play a fundamental role in shaping host ecology and evolution, influencing physiological condition, behavior, and reproductive success (Hamilton and Zuk 1982, Sheldon and Verhulst 1996). Blood parasites have been widely studied in birds because of their potential effects on survival and fitness (Marzal et al. 2005, Martínez-de la Puente et al. 2010). The genera Haemoproteus, Plasmodium, and Leucocytozoon are commonly found in avian hosts, often impacting individual condition and reproductive investment (Merino et al. 2000). However, parasite prevalence and species composition can vary significantly across host populations, influenced by environmental conditions, vector abundance, local host species pool, and host immunity (Sanz et al. 2002, Hellgren et al. 2008, Pacheco et al. 2022). In the case of migratory swallows, for example, the arrival date after spring migration appears to be influenced by parasitic load (Møller et al. 2004), and breeding success decreases with later laying dates (Winkler et al. 2020). However, this delay differs across populations and between sexes (Møller et al. 2004).
Across the Americas, Leucocytozoon prevalence increases with latitude and altitude and consequently with decreasing temperature (Lotta et al. 2016, Fecchio et al. 2020), a pattern also reported in Chile and contrasting with that observed for Plasmodium and Haemoproteus (Merino et al. 2008, Doussang et al. 2019). These differences may reflect the environmental conditions required by the parasites for development within vectors. Haemosporidian infections are typically acquired during periods of high vector activity (e.g., breeding seasons in temperate regions), although transmission may also occur during migration or on wintering grounds (Piersma and van der Velde 2012, Pulgarín et al. 2018, de Angeli Dutra et al. 2021). Leucocytozoon, transmitted by Simuliidae (blackflies) may be transmitted at temperatures as low as 12 °C (Fecchio et al. 2020, Mozaffer et al. 2022). In contrast, Plasmodium and Haemoproteus, transmitted by Culicidae (mosquitoes), Hippoboscidae (louse flies) and Ceratopogonidae (biting midges), respectively, develop optimally above 24 °C (LaPointe et al. 2010, Zamora-Vilchis et al. 2012, Oakgrove et al. 2014, Mozaffer et al. 2022).
Currently, information on the prevalence of parasitic infections in wild bird populations in South America remains limited. Research has increased in recent years (e.g., Doussang et al. 2019, Fecchio et al. 2020, 2021), yet basic data on prevalence, vectors, and the effects of parasitic load on life-history traits remain scarce. Although hemoparasite loads appear to be generally low in this region (Merino et al. 2008), some bird species exhibit high prevalence, such as the migrant White-crested Elaenia (Elaenia albiceps; Merino et al. 2008) and the Rufous-collared Sparrow (Zonotrichia capensis; Doussang et al. 2019).
Among the less studied taxa are species within the genus Tachycineta, despite its wide distribution across America (Marion et al. 2020). Information on hemoparasites in this genus is limited and heterogeneous, with a strong bias toward the Northern Hemisphere Tree Swallow (Tachycineta bicolor; Table 1; Supplementary Material Table S1). Reported haematozoa in Tachycineta include single infections with Trypanosoma, Leucocytozoon, Plasmodium, or Haemoproteus, as well as mixed Leucocytozoon-Haemoproteus infections (Martinsen et al. 2008, Murdock 2009, Medeiros et al. 2015, Turcotte et al. 2018, Rodriguez et al. 2021).
The Chilean Swallow (Tachycineta leucopyga) is a Neotropical migratory passerine and the southernmost representative of this cavity-nesting genus. Only one study has assessed its hemoparasites, reporting no infections in 26 individuals sampled across three locations in Chile, two of which were islands (Merino et al. 2008). Cavity-nesters generally appear to be less affected by blood parasites than open-cup nesters (e.g., Rodriguez et al. 2021). However, both adults and nestlings can still become infected, as documented for cavity nesting birds in the Americas and Europe (Merino and Potti 1995, Turcotte et al. 2018).
In Argentina, the Chilean Swallow breeds along a broad latitudinal gradient from northern Andean Patagonia (~39° S; Bianchini 2024) to Tierra del Fuego (~54° S; Marion 2020). The harsher environmental conditions and shorter breeding season at higher latitudes likely impose distinct physiological and ecological pressures on individuals breeding in Tierra del Fuego compared to those in northern populations (Ippi et al. 2024). Parasite load could represent an important factor influencing behavioral and reproductive trade-offs in this species. However, information on the prevalence of haemosporidian parasites in T. leucopyga across its breeding range remains very limited.
In this study, we aimed to assess the prevalence of haemosporidians in Chilean Swallow populations from Argentine Patagonia and to compare their occurrence between two latitudinal extremes of the species’ breeding range. Additionally, we aimed to identify and characterize haemosporidian lineages infecting the Chilean Swallow and to examine their phylogenetic relationships with related lineages circulating in other South American bird species. Given the temperature of the study sites and the thermal optima of the parasites, we predicted that the southernmost population would show higher infection rates with Leucocytozoon, whereas the northern population would exhibit higher prevalence of Haemoproteus and Plasmodium (Merino et al. 2008, Doussang et al. 2019). Based on the generally lower haemoparasite prevalence reported for Neotropical compared to Nearctic regions (White et al. 1978), as well as the absence of records for this species in Chilean populations (Merino et al. 2009), we expected overall prevalence to be lower than that reported for Tachycineta swallows in the Northern Hemisphere.
METHODS
Study area
The study was conducted in two populations in Argentine Patagonia. The northernmost population is located in northwestern Patagonia near the city of San Carlos de Bariloche (41° S, 71° W) and it was monitored during 2021–2022 and 2022–2023 austral breeding seasons at Fortín Chacabuco and El Cóndor ranches, where 120 and 50 nest boxes are installed, respectively. The southernmost population is in Tierra del Fuego (54° S, 68° W) and was studied during the 2006–2007 and 2007–2008 breeding seasons.
Both nest box populations were monitored by using the standardized protocol of the Golondrinas de las Americas project (https://blogs.cornell.edu/golondrinas/), allowing for reliable comparisons. Briefly, this protocol involved regular visits to record breeding phenology and other reproductive variables, such as clutch size and nest size (variables also included in other projects). To minimize the risk of nest abandonment, breeding adults were captured only after egg hatching, using a wig-wag trap (Massoni et al. 2007), banded with both a color and a numbered metal ring, and sampled for blood. In the northern population (Bariloche), nestlings were similarly banded and blood-sampled at a mean age of 15 d (standard deviation = 0.97). Blood samples (~30 µl) were obtained from all individuals by puncturing the brachial vein (Owen 2011). Blood samples were preserved in 0.5 ml lysis buffer (100 mM Tris, 100 mM EDTA, 10 mM NaCl, 2% SDS).
Molecular and phylogenetic analysis
Preserved DNA was extracted from whole blood by using QIAamp DNA Micro Kit (QIAGEN GmbH, Hilden, Germany). Extracted DNA was screened for haemosporidians by using a nested polymerase chain reaction (PCR) protocol targeting the mitochondrial cytochrome b gene (cytb, 1,131 bp) with primers described by Pacheco et al. (2018, 2019). A key advantage of these primers is that they have been proven to be sensitive to amplify all haemosporidian genera across all known vertebrate hosts (Pacheco et al. 2018, 2019, 2022, 2025), and they generate a longer gene fragment containing more parsimony-informative sites compared to the MalAvi barcoding protocol (Bensch et al. 2009). This provides greater phylogenetic resolution and potential for species identification, which is required to understand haemosporidian relationships as more sequences become available in public databases (Pacheco et al. 2018). Primary PCR amplifications were carried out in a 50 μl volume with 2 and 5 μl of total genomic DNA (depending on the total DNA per sample, which varied between 10 and 350 ng/µl), 2.5 mM MgCl2, 1 × PCR buffer, 0.25 mM of each deoxynucleoside triphosphate, 0.4 μM of each primer, and 0.03 U/μl AmpliTaq polymerase (Applied Biosystems, Thermo Fisher Scientific, USA). External PCR primers used were forward AE298 5′-TGT AAT GCC TAG ACG TAT TCC 3′ and reverse AE299 5′-GT CAA WCA AAC ATG AAT ATA GAC 3′. Negative (distilled water) and two positive controls (samples from infected birds with Plasmodium lutzi and Haemoproteus erythrogravidus) were also included.
Primary PCR conditions included a partial denaturation at 94 °C for 4 min, followed by 36 cycles with 1 min at 94 °C, 1 min at 53 °C, and a 2 min extension at 72 °C. In the last cycle, a final 10 min extension at 72 °C was added. Nested PCR mix and conditions were the same as the primary PCR, but using 1 μl of the primary PCR and an annealing temperature of 56 °C. Internal PCR primers used were forward AE064 5′-T CTA TTA ATT TAG YWA AAG CAC 3′ and reverse AE066 5′-G CTT GGG AGC TGT AAT CAT AAT 3′.
PCR amplified products (50 μl) were excised from agarose gels and purified by the QIAquick Gel Extraction Kit (QIAGEN GmbH, Hilden, Germany). Both strands of the cytb gene fragments were directly sequenced using the Sanger method at GENEWIZ (Azenta Life Sciences, New Jersey, USA). Partial cytb gene sequence obtained here was deposited in GenBank under the accession number PZ065612.
After comparing the sequence obtained in this study (PZ065612) against sequences available in GenBank (Benson et al. 2015) and MalAvi (Bensch et al. 2009) using BLAST, a phylogenetic analysis was performed on a partial cytb gene alignment (N = 59 sequences; 466 bp excluding gaps). This alignment was constructed with sequences from well-known lineages closely related to the sequence reported here and parasite species based on morphology (Valkiūnas and Iezhova 2023) that were available in GenBank (Benson et al. 2015) and MalAvi (Bensch et al. 2009) databases at the time of this study (July 2025) using MAFFT v.7 using default parameters (Katoh and Standley 2013). Sequences from the genera Haemoproteus and Leucocytozoon (Akiba) caulleryi were used as an outgroup. Then, phylogenetic relationships were inferred by using a Bayesian method implemented in MrBayes v3.2.7 with the default priors (Ronquist and Huelsenbeck 2003). A general time-reversible model with gamma-distributed substitution rates and a proportion of invariant sites (GTR + Γ + I) was used, as the best substitution model determined by MEGA v7.0.14 (Kumar et al. 2016). Posterior probabilities for the nodes were estimated from two independent chains of 3 x 106 Markov Chain Monte Carlo (MCMC) steps by sampling every 1000 generations. Convergence was assumed when the value of the potential scale reduction factor (PSRF) was between 1.00 and 1.02, and the average standard deviation of the posterior probability was <0.01 (Ronquist and Huelsenbeck 2003). A “burn-in” of 25% of the sample was discarded. Phylogenetic trees were visualized by using FigTree v1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/). Parasite species names, GenBank accession numbers, and lineage name of the sequences used in this analysis are shown in the phylogenetic trees. Average evolutionary distance over sequence pairs of Leucocytozoon spp. lineages were estimated as substitutions per site using the Tamura-Nei substitution model implemented in MEGA v7.0.14 (Kumar et al. 2016).
RESULTS
We analyzed 38 DNA samples from adult swallows in Tierra del Fuego, and 106 samples from adults (49 females and 57 males), as well as 65 15-day-old nestlings from 19 nests in the Bariloche area. We detected only a single infected individual—a female carrying Leucocytozoon sp.—in the Tierra del Fuego population.
This parasite was identical to the Leucocytozoon sp. lineage DIUDIU11 reported in MalAvi (Submitted by Staffan Bensch on 5 May 2025; Supplementary Material Table S1A-C). It has been found only in the South American Diuca diuca (Thraupidae; MK947686: Argentina), Troglodytes musculus (Troglodytidae; ON399874: Argentina), Elaenia albiceps (Tyrannidae; MK947687: Argentina and Brazil), Aphrastura spinicauda (Furnariidae; MN083254: Chile), and Philydor erythrocercum (Furnariidae; MN458607: Peru).
With very high node supports (PP > 0.94), phylogenetic relationships showed that Leucocytozoon sp. DIUDIU11 shares a common ancestor with TROAED01, APSPI06, ELAALB06, CARBAR02, and TFUS12 lineages, all of them from South America (Fig. 1), with very low genetic distance between them (Table 2). This monophyletic group also shared a common ancestor with Leucocytozoon fringillinarum (Fig. 1). The genetic distance, estimated as substitutions per site, between Leucocytozoon sp. DIUDIU11 and Leucocytozoon fringillinarum (KY653765, Spinus spinus) was 0.017 ± 0.006 (1.7%).
DISCUSSION
Hemoparasite prevalence in aerial insectivores is generally low (White and Benett 1978, White et al. 1978, Valkiūnas 2005, Valkiūnas et al. 2020), and Chilean Swallows seem to follow this pattern. In our study, only one adult from Tierra del Fuego was infected with Leucocytozoon, and no chicks or adults from Bariloche tested positive for any haemosporidian, consistent with previous reports from Chile (Merino et al. 2008). Similar low prevalence has been observed in other aerial insectivores worldwide, such as Tree Swallows (only one infection among 221 examined via blood smears; Shutler et al. 2004) and Alpine Swifts (Tachymarptis melba: zero infections among 40 individuals, despite heavy infestations of louse flies; Tella et al. 1995). Nevertheless, some swallow species can exhibit high hemoparasite prevalence (~38%, Barn Swallow Hirundo rustica and Common House Martin Delichon urbicum; Grabow et al. 2024), with infections affecting mobility, foraging, and potentially fitness.
Within Tachycineta, prevalence is variable, although it was often assessed with small sample sizes (Table 1, Supplementary Material Table S1). Environmental conditions, local host density, and sex can influence infection rates (Sanz et al. 2002, Møller et al. 2004, Hellgren et al. 2008). Low prevalence in aerial insectivores is commonly attributed to reduced vector contact because of prolonged flight and short resting bouts (Santiago-Alarcón et al. 2012). However, bite frequency provides only a partial view of this host-parasite system (Eastwood et al. 2019). Although a tendency toward lower prevalence in cavity nesting species compared with open-cup nesters has been reported (Rodriguez et al. 2021), incubating females, nestlings confined to warm nests, and nest boxes located near water are expected to provide suitable conditions for vectors. Yet, no infections were detected in our northernmost population, suggesting either low vector abundance or the absence of local infectious hosts (Whitman and Wilson 1992, Di Iorio et al. 2008, Valkiūnas et al. 2020, Rodriguez et al. 2021, Graciolli and Laps 2024). Unfortunately, data on the abundance and diversity of Diptera in Patagonia remain scarce.
A possibility worth exploring is that infections are acquired on the wintering grounds, followed by high mortality during spring migration. This mechanism has been proposed to explain the absence of Haemoproteus infections in juveniles of Barn Swallow and Common House Martin during the European breeding season, where transmission is thought to occur mainly in sub-Saharan Africa (Valkiūnas 2005, Piersma and van der Velde 2012), a predominantly tropical region. In contrast, Chilean Swallows winter in subtropical to warm-temperate regions of South America, when dipteran availability may be lower or more seasonal (e.g., Stein et al. 2016). Additionally, these landscapes are often intensively farmed and exposed to high pesticide use (Alonso et al. 2018, Marion et al. 2020, Codesido and Bilenca 2021), which may further alter host-vector-parasite dynamics (e.g., Turcotte et al. 2018). Under this scenario, infected individuals in poor condition could be less likely to complete migration, reducing apparent prevalence (Bensch et al. 2007). However, in the absence of direct evidence in Chilean Swallows, this mechanism remains hypothetical.
An alternative, not mutually exclusive, explanation is that Chilean Swallows may exhibit higher resistance to haemosporidian infection (i.e., the ability to prevent infection through immune responses; see, e.g., Sorci 2013, Eastwood et al. 2019). Given the high energetic demands of migratory aerial insectivores, selection may favor the evolution of resistance to hemoparasites. However, host phylogeny has been shown to influence infection propensity (Gupta et al. 2020), and other migratory species of Tachycineta exhibit haemosporidian infections; therefore, strong resistance in Chilean Swallows appears less likely. Reduced immune function during migration and/or increased exposure to infected vectors en route have also been proposed as factors affecting hemoparasite prevalence (Owen and Moore 2008, de Angeli Dutra et al. 2021), suggesting that migration itself may modulate infection risk. In this context, differences in migratory distances among swallow species could contribute to interspecific variation in prevalence (Altizer et al. 2011), although the relatively intermediate distances traveled by Chilean swallows (1500–3000 km) overlap with those of species showing higher prevalence (2000–7000 km; see Table 1; Grabow et al. 2024, see also Møller et al. 2004).
The only hemoparasite detected in Chilean Swallows was Leucocytozoon in the southernmost population. This finding is consistent with expectations that Leucocytozoon prevalence and diversity increase with latitude, vegetation, and humidity (Merino et al. 2008, Fecchio et al. 2020). Phylogenetically, the lineage (DIUDIU11), previously reported only for South America (Supplementary Material Table S1), is closely related to Leucocytozoon fringillinarum Woodcock 1910; a parasite reported in over 200 passerine species (Valkiūnas 2005). The lineage we detected is shared among several other South American species belonging to different passerine families, such as Furnariidae, Tyrannidae (Suboscines), and Thraupidae (Oscines), reflecting broad host sharing common in avian haemosporidians (Bensch et al. 2000, Szymanski and Lovette 2005).
Swallow populations are declining in the Northern Hemisphere (Michel et al. 2016, Shipley et al. 2020), highlighting the need to understand ecological and parasitological factors affecting fitness and survival in South American populations, which remain poorly studied. Indeed, the combined pressures of parasitism, habitat degradation, and climate change could disproportionately affect migratory species that forage on the wing (Martínez and Merino 2011, Turcotte et al. 2018, Grabow et al. 2024). Climate warming may further enhance the transmission of some parasites by increasing their prevalence (Zamora-Vilchis et al. 2012, Mennerat et al. 2021). We emphasize the need to investigate the health of migratory birds—particularly hemoparasites and their vectors—across their full annual cycle, including both breeding and wintering grounds. Future studies integrating microscopy with PCR—targeting the mitochondrial cytb gene using both existing and newly updated primer sets—are required to fully characterize the haemosporidian assemblage associated with this bird species in South America. Whereas microscopy is essential to confirm parasite intensity and describe species, molecular methods are vital when parasitemia falls below the microscopic detection threshold (0.01%, or 1 parasite per 10,000 erythrocytes). PCR can detect extremely low-level infections, identifying approximately one parasite in 50,000 cells (Valkiūnas et al. 2009). As demonstrated in previous population studies utilizing the same protocol employed here, this approach significantly minimizes the likelihood of false-negative results (Pacheco et al. 2022, 2025).
Supplementary material is available at https://figshare.com/s/8f41da7de68039cafdf8.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Conceived the idea, designed the study, formulated questions or hypotheses: Silvina Ippi, María Pacheco, Melina Barrionuevo, Valeria S. Ojeda, Valentina Ferretti;
Collected data, conducted the research, including laboratory procedures: all authors;
Wrote the paper: Silvina Ippi, Valentina Ferretti, and María Pacheco. All authors critically contributed to the drafts and gave final approval for publication.
Developed or designed methods: Silvina Ippi, María Pacheco, Ananías Escalante;
Analyzed the data: María Pacheco.
ACKNOWLEDGMENTS
We thank the Fortín Chacabuco TNC crew (especially the G. Hulsegge and N. Rodríguez Argumedo family), as well as El Cóndor ranch. We are especially grateful to M. Liljesthröm for facilitating the blood samples from Tierra del Fuego, and L. Chazarreta, A. Ortiz, G. Ortiz, M. Cieri, Natalie Dudinszky, P. Canavari, A. Mayoral, P. Alvear, and L. Casalins for their field support during this study. We also thank B. Butcher, L. Campagna, and Irby Lovette for hosting and supporting the DNA extraction work in the Fuller Evolutionary Biology lab (Cornell Lab of Ornithology). This work was partially supported by Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación (grant PICT 2018-2756) and by the Consejo Nacional de Investigaciones Científicas y Técnicas (grant PIP 11220200102927CO). Fieldwork was conducted under permits No. 1523 to VO and No. 1593 to MB from the Dirección Regional Patagonia Norte (DRPN, APN), and No. 023932- SAYDS-2018 from the Secretaría de Ambiente y Cambio Climático, Río Negro. All procedures were approved by the INIBIOMA IACUC working protocol No. 51.
DATA AVAILABILITY
Data/code derived from public domain resources.
LITERATURE CITED
Alonso, L. L., P. M. Demetrio, M. A. Etchegoyen, and D. J. Marino. 2018. Glyphosate and atrazine in rainfall and soils in agroproductive areas of the pampas region in Argentina. Science of the Total Environment 645:89-96. https://doi.org/10.1016/j.scitotenv.2018.07.134
Altizer, S., R. Bartel, and B. A. Han. 2011. Animal migration and infectious disease risk. Science 21(331):296-302. https://doi.org/10.1126/science.1194694
Bensch, S., M. Stjernman, D. Hasselquist, Ö. Östman, B. Hannson, H. Westerdahl, and R. T. Pinheiro. 2000. Host specificity in avian blood parasites: a study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proceeding of Royal Society London B, Biological Science 267:1583-1589. https://doi.org/10.1098/rspb.2000.1181
Bensch, S., J. Waldenström, N. Jonzén, H. Westerdahl, B. Hansson, D. Sejberg, and D. Hasselquist. 2007. Temporal dynamics and diversity of avian malaria parasites in a single host species. Journal of Animal Ecology 76:112-122. https://doi.org/10.1111/j.1365-2656.2006.01176.x
Bensch, S., O. Hellgren, and J. Pérez-Tris. 2009. MalAvi: a public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Molecular Ecology Resources 9:1353-1358. https://doi.org/10.1111/j.1755-0998.2009.02692.x
Benson D. A., K. Clark, I. Karsch-Mizrachi, D. J. Lipman, J. Ostell, and E. W. Sayers. 2015. GenBank. Nucleic Acids Research 43(D1):D30-35. https://doi.org/10.1093/nar/gku1216
Bianchini, M. 2024. Actualización y ampliación de la distribución geográfica reproductiva de tres especies de golondrinas en la Patagonia (Progne tapera fusca, Tachycineta leucopyga y Hirundo rustica erythrogaster, Passeriformes: Hirundinidae) y aspectos observados durante el periodo de cría. Acta Zoológica Lilloana 68(1):171-198.
Codesido, M., and D. Bilenca. 2021. Avian assemblages associated with different grasslands managements in cattle production systems in the pampas of Argentina. Perspectives in Ecology and Conservation 19:464-474. https://doi.org/10.1016/j.pecon.2021.07.003
de Angeli Dutra, D., A. Fecchio, É. M. Braga, and R. Poulin. 2021. Migratory birds have higher prevalence and richness of avian haemosporidian parasites than residents. International Journal for Parasitology 51:877-882. https://doi.org/10.1016/j.ijpara.2021.03.001
Di Iorio, O., F. Bulit, F. Aldatz, and V. Massoni. 2008. Insects found in birds’ nests from Argentina: Tachycineta leucorrhoa leucorrhoa (Vieillot, 1817) [Aves: Hirundinidae], a new host for Acanthocrios furnarii (Cordero & Vogelsang, 1928) [Hemiptera: Cimicidae]. Zootaxa 1896:1-24. https://doi.org/10.11646/zootaxa.1896.1.1
Doussang, D., D. González-Acuña, L. G. Torres-Fuentes, S. C. Lougheed, R. B. Clemente-Carvalho, K. C. Greene, and J. A. Vianna. 2019. Spatial distribution, prevalence and diversity of haemosporidians in the Rufous-collared Sparrow, Zonotrichia capensis. Parasites & Vectors 12:2. https://doi.org/10.1186/s13071-018-3243-4
Durrant, K. L., J. S. Beadell, F. Ishtiaq, G. R. Graves, S. L. Olson, E. Gering, M. A. Peirce, C. M. Milensky, B. K. Schmidt, C. Gebhard, and R. C. Fleischer. 2006. Avian hematozoa in South America: a comparison of temperate and tropical zones. Ornithological Monographs 60:98-111. https://doi.org/10.2307/40166831
Eastwood, J. R., L. Peacock, M. L. Hall, M. Roast, S. A. Murphy, A. G. da Silva, and A. Peters. 2019. Persistent low avian malaria in a tropical species despite high community prevalence. International Journal for Parasitology: Parasites and Wildlife 8:88-93. https://doi.org/10.1016/j.ijppaw.2019.01.001
Fecchio, A., J. A. Bell, M. Bosholn, J. A. Vaughan, V. V. Tkach, H. L. Lutz, V. R. Cueto, C. A. Gorosito, D. González-Acuña, C. Stromlund, et al. 2020. An inverse latitudinal gradient in infection probability and phylogenetic diversity for Leucocytozoon blood parasites in New World birds. Journal of Animal Ecology 89:423-435. https://doi.org/10.1111/1365-2656.13117
Fecchio, A., R. M. Ribeiro, F. C. Ferreira, D. de Angeli Dutra, G. Tolesano-Pascoli, R. D. Alquezar, A. U. Khan, M. Pichorim, P. A. Moreira, M. J. Costa-Nascimento, et al. 2021. Higher infection probability of haemosporidian parasites in Blue-black Grassquits (Volatinia jacarina) inhabiting native vegetation across Brazil. Parasitology International 80:102204. https://doi.org/10.1016/j.parint.2020.102204
Grabow, M., W. Ullman, C. Landgraf, R. Sollman, C. Scholz, R. Nathan, S. Toledo, R. Lühken, J. Fickel, F. Jeltsch, et al. 2024. Sick without signs. Subclinical infections reduce local movements, alter habitat selection, and cause demographic shifts. Communications Biology 7:1426. https://doi.org/10.1038/s42003-024-07114-4
Graciolli, G., and R. R. Laps. 2024. A checklist of louse flies (Diptera: Hippoboscidae) and their avian hosts from the Americas. Integrative Systematics 7(2):103-153. https://doi.org/10.18476/2024.995170
Gupta, P., C. K. Vishnudas, V. V. Robin, and G. Dharmarajan. 2020. Host phylogeny matters: examining sources of variation in infection risk by blood parasites across a tropical montane bird community in India. Parasites & Vectors 13:536. https://doi.org/10.1186/s13071-020-04404-8
Hamilton, W. D., and M. Zuk. 1982. Heritable true fitness and bright birds: a role for parasites? Science 218(4570):384-387. https://doi.org/10.1126/science.7123238
Hellgren, O., S. Bensch, and B. Malmqvist. 2008. Bird hosts, blood parasites and their vector—associations uncovered by molecular analyses of blackfly blood meals. Molecular Ecology 17:1605-1613. https://doi.org/10.1111/j.1365-294X.2007.03680.x
Ippi, S., V. Ojeda, N. Dudinszky, M. Cieri, B. Riovitti, V. Ferretti, and M. Barrionuevo. 2024. Filling latitudinal gaps in Tachycineta swallows: reproductive strategies in a South American species. Emu 124(3-4):240-251. https://doi.org/10.1080/01584197.2024.2353380
Katoh K., and D. M. Standley. 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution 30(4):772-780. https://doi.org/10.1093/molbev/mst010
Kumar S., G. Stecher, and K. Tamura. 2016. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33:1870-1874. https://doi.org/10.1093/molbev/msw054
LaPointe, D. A., M. L. Goff, and C. T. Atkinson. 2010. Thermal constraints to the sporogonic development and altitudinal distribution of avian malarian Plasmodium relictum in Hawai’i. Journal of Parasitology 96:318-324. https://doi.org/10.1645/GE-2290.1
Lotta I. A., M. A. Pacheco, A. A. Escalante, A. D. González, J. S. Mantilla, L. I. Moncada, P. H. Adler, and N. E. Matta. 2016. Leucocytozoon diversity and possible vectors in the Neotropical highlands of Colombia. Protist 167(2):185-204. https://doi.org/10.1016/j.protis.2016.02.002
Marion, J. S. (2020). Chilean Swallow (Tachycineta leucopyga), version 1.0. In T. S. Schulenberg, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.chiswa1.01
Martínez, J., and S. Merino. 2011. Host-parasite interactions under extreme climatic conditions. Current Zoology 57(3):390-405. https://doi.org/10.1093/czoolo/57.3.390
Martínez-de la Puente, J., S. Merino, G. Tomás, J. Moreno, J. Morales, E. Lobato, S. García-Fraile, and E. J. Belda. 2010. The blood parasite Haemoproteus in a wild bird: a medication experiment. Biology Letters 6:663-665. https://doi.org/10.1098/rsbl.2010.0046
Martinsen, E. S., B. J. Blumberg, R. J. Eisen, and J. J. Schall. 2008. Avian haemosporidian parasites from northern California Oak Woodland and chaparral habitats. Journal of Wildlife Diseases 44:260-268. https://doi.org/10.7589/0090-3558-44.2.260
Marzal, A., F. de Lope, C. Navarro, and A. P. Møller. 2005. Malarial parasites decrease reproductive success: an experimental study in a passerine bird. Population Ecology 142:541-545. https://doi.org/10.1007/s00442-004-1757-2
Massoni, V., F. Bulit, and J. C. Reboreda. 2007. Breeding biology of the White-rumped Swallow Tachycineta leucorrhoa in Buenos Aires Province, Argentina. Ibis 149:10-17. https://doi.org/10.1111/j.1474-919X.2006.00589.x
Medeiros, M. C. I., R. Ricklefs, J. D. Brawn, and G. L. Hamer. 2015. Plasmodium prevalence across avian host species is positively associated with exposure to mosquito vectors. Parasitology 142:1612-1620. https://doi.org/10.1017/S0031182015001183
Mennerat, A., A. Charmantier, P. Perret, S. Hurtrez-Boussès, and M. M. Lambrechts. 2021. Parasite intensity is driven by temperature in a wild bird. Peer Community 1:e60. https://doi.org//doi.org/10.24072/pcjournal.65
Merino, S., J. Moreno, J. J. Sanz, and E. Arriero. 2000. Are avian blood parasites pathogenic in the wild? A medication experiment in Blue Tits (Parus caeruleus). Proceedings of Royal Society London B, Biological Sciences 267:2507-2510. https://doi.org/10.1098/rspb.2000.1312
Merino, S., J. Moreno, R. A. Vásquez, J. Martínez, I. Sánchez-Monsálvez, C. F. Estades, S. Ippi, P. Sabat, R. Rozzi, and S. McGehee. 2008. Haematozoa in forest birds from southern Chile: latitudinal gradients in prevalence and parasite lineage richness. Austral Ecology 33:329-340. https://doi.org/10.1111/j.1442-9993.2008.01820.x
Merino, S., and J. Potti. 1995. High prevalence of haematozoa in nestlings of a passerine species the Pied Flycatcher (Ficedula hypoleuca). The Auk 112:1041-1043. https://doi.org/10.2307/4089037
Michel N. L., A. C. Smith, R. G. Clark, C. A. Morrissey, and K. A. Hobson. 2016. Differences in spatial synchrony and interspecific concordance inform guild-level population trends for aerial insectivorous birds. Ecography 39:774-786. https://doi.org/10.1111/ecog.01798
Møller, A. P., F. de Lope, and N. Saino. 2004. Parasitism, immunity, and arrival date in a migratory bird, the Barn Swallow. Ecology 85(1):206-219. https://doi.org/10.1890/02-0451
Mozaffer F., G. I. Menon, and F. Ishtiaq. 2022. Exploring the thermal limits of malaria transmission in the western Himalaya. Ecology and Evolution 12(9):e9278. https://doi.org/10.1002/ece3.9278
Murdock, C. C. 2009. Studies on the ecology of avian malaria in an alpine ecosystem. Dissertation. University of Michigan, Ann Arbor, Michigan, USA.
Oakgrove K. S., R. J. Harrigan, C. Loiseau, S. Guers, B. Seppi, and R. N. M. Sehgal. 2014. Distribution, diversity and drivers of blood-borne parasite co-infections in Alaskan bird populations. International Journal for Parasitology 44(10):717-727. https://doi.org/10.1016/j.ijpara.2014.04.011
Owen, J. C. 2011. Collecting, processing, and storing avian blood: a review. Journal of Field Ornithology 82(4):339-354. https://doi.org/10.1111/j.1557-9263.2011.00338.x
Owen, J. C., and F. R. Moore. 2008. Swainson’s thrushes in migratory disposition exhibit reduced immune function. Journal of Ethology 26(3):383-388. https://doi.org/10.1007/s10164-008-0092-1
Pacheco, M. A., A. S. Cepeda, R. Bernotienë, I. A. Lotta, N. E. Matta, G. Valkiūnas, and A. A. Escalante. 2018. Primers targeting mitochondrial genes of avian haemosporidians: PCR detection and differential DNA amplification of parasites belonging to different genera. International Journal for Parasitology 48(8):657-670. https://doi.org/10.1016/j.ijpara.2018.02.003
Pacheco M. A., F. C Ferreira, C. J. Logan, K. B. McCune, M. P. MacPherson, S. Albino Miranda, D. Santiago-Alarcón, and A. A. Escalante. 2022. Great-tailed Grackles (Quiscalus mexicanus) as a tolerant host of avian malaria parasites. PLoS ONE 17(8):e0268161. https://doi.org/10.1371/journal.pone.0268161
Pacheco M. A., M. A. García-Amado, J. Manzano, N. E. Matta, and A. A. Escalante. 2019. Blood parasites infecting the Hoatzin (Opisthocomus hoazin), a unique neotropical folivorous bird. PeerJ 7:e6361. https://doi.org/10.7717/peerj.6361
Pacheco, M. A., E. A. Miller, J. Harl, F. C. Ferreira, A. S. Cepeda, G. Valkiūnas, S. Beckerman, M. Oswald, N. E. Mateus-Pinilla, and A. A. Escalante. 2025. Haemoproteus syrnii and other haemosporidians infecting owls from North America. Malaria Journal 24(1):365. https://doi.org/10.1186/s12936-025-05612-2
Piersma, T., and M. van der Velde. 2012. Dutch House Martins Delichon urbicum gain blood parasite infections over their lifetime, but do not seem to suffer. Journal of Ornithology 153:907-912. https://doi.org/10.1007/s10336-012-0826-2
Pulgarín, P. C., C. Gómez, N. J. Bayly, S. Bensch, A. M. FitzGerald, N. Starkloff, J. J. Kirchman, A. M. González-Prieto, K. A. Hobson, J. Ungvari-Martin, et al. 2018. Migratory birds as vehicles for parasite dispersal? Infection by avian haemosporidians over the year and throughout the range of a long-distance migrant. Journal of Biogeography 46:83-96. https://doi.org/10.1111/jbi.13453
Rodriguez, M. D., P. Doherty, A. J. Piaggio, and K. P. Huyvaert. 2021. Sex and nest type influence avian blood parasite prevalence in a high-elevation bird community. Parasites & Vectors 14(1):145. https://doi.org/10.1186/s13071-021-04612-w
Ronquist F., and J. P. Huelsenbeck. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572-1574. https://doi.org/10.1093/bioinformatics/btg180
Santiago-Alarcón, D., V. Palinauskas, and H. M. Schaefer. 2012. Diptera vectors of avian Haemosporidian parasites: untangling parasite life cycles and their taxonomy. Biological Reviews 87:928-964. https://doi.org/10.1111/j.1469-185X.2012.00234.x
Sanz, J. J., J. Moreno, E. Arriero, and S. Merino. 2002. Reproductive effort and blood parasites of breeding pied flycatchers: the need to control for interannual variation and initial health state. Oikos 96:299-306. https://doi.org/10.1034/j.1600-0706.2002.960212.x
Sheldon, B. C., and S. Verhulst. 1996. Ecological immunology: costly parasite defences and trade-offs in evolutionary ecology. Trends in Ecology and Evolution 11(8):317-321. https://doi.org/10.1016/0169-5347(96)10039-2
Shipley, J. R., C. W. Twining, C. C. Taff, M. N. Vitousek, A. Flack, and D. W. Winkler. 2020. Birds advancing lay dates with warming springs face greater risk of chick mortality. Proceedings of the National Academy of Sciences 117(41):25590-25594. https://doi.org/10.1073/pnas.2009864117
Shutler, D., A. Mullie, and R. G. Clark. 2004. Tree swallow reproductive investment, stress, and parasites. Canadian Journal of Zoology 82(3):442-448. https://doi.org/10.1139/z04-016
Smith, J. D., S. A. Gill, K. M. Baker, and M. J. Vonhof. 2018. Prevalence and diversity of avian Haemosporida infecting songbirds in southwest Michigan. Parasitology Research 117:471-489. https://doi.org/10.1007/s00436-017-5724-3
Sorci, G. 2013. Immunity, resistance and tolerance in bird-parasite interactions. Parasite Immunology 35(11):350-361. https://doi.org/10.1111/pim.12047
Stabler, R. M., and N. J. Kitzmiller. 1970. Hematozoa from Colorado birds. III. Passeriformes. The Journal of Parasitology 56(1):12-16. https://doi.org/10.2307/3277443
Stein, M., M. Santana, L. M. Galindo, E. Etchepare, J. A. Willener, and W. R. Almirón. 2016. Culicidae (Diptera) community structure, spatial and temporal distribution in three environments of the province of Chaco, Argentina. Acta Tropica 156:57-67. https://doi.org/10.1016/j.actatropica.2015.12.002
Szymanski, M. M., and I. J. Lovette. 2005. High lineage diversity and host sharing of malarial parasites in a local avian assemblage. The Journal of Parasitology 91(4):768-774. https://doi.org/10.1645/GE-417R1.1
Tajima, F., and M. Nei. 1984. Estimation of evolutionary distance between nucleotide sequences. Molecular Biology and Evolution 1:269-285. https://doi.org/10.1093/oxfordjournals.molbev.a040317
Tamura, K., and S. Kumar. 2002. Evolutionary distance estimation under heterogeneous substitution pattern among lineages. Molecular Biology and Evolution 19:1727-1736. https://doi.org/10.1093/oxfordjournals.molbev.a003995
Tella, J. L., C. Gortázar, Á. Gajón, and J. J. Osácar. 1995. Apparent lack of effects of a high louse-fly infestation (Diptera, Hippoboscidae) on adult colonial Alpine Swifts. Ardea 83:435-439.
Turcotte, A., M. Bélisle, F. Pelletier, and D. Garant. 2018. Environmental determinants of haemosporidian parasite prevalence in a declining population of Tree Swallows. Parasitology 145:961-970. https://doi.org/10.1017/S0031182017002128
Valkiūnas, G. 2005. Avian malaria parasites and other haemosporidia. CRC Press, Boca Raton, Florida, USA.
Valkiūnas, G., T. A. Iezhova, C. Loiseau, and R. N. M. Sehgal. 2009. Nested cytochrome B polymerase chain reaction diagnostics detect sporozoites of hemosporidian parasites in peripheral blood of naturally infected birds. The Journal of Parasitology 95:1512-1515. https://doi.org/10.1645/GE-2105.1
Valkiūnas, G., and T. A. Iezhova. 2023. Insights into the biology of Leucocytozoon species (Haemosporida, Leucocytozoidae): why is there slow research progress on agents of leucocytozoonosis? Microorganisms 11(5):1251. https://doi.org/10.3390/microorganisms11051251
Valkiūnas, G., M. Ilgūnas, C. R. F. Chagas, R. Bernotiené, T. A. Iezhova. 2020. Molecular characterization of swallow haemoproteids, with description of one new Haemoproteus species. Acta Tropica 207:105486. https://doi.org/10.1016/j.actatropica.2020.105486
White, E. M., E. C. Greiner, G. F. Bennett, and C. M. Herman. 1978. Distribution of the hematozoa of Neotropical birds. Revista de Biología Tropical 26(1):43-102.
White, E. M., and G. F. Bennett. 1978. Avian Haemosproteidae. 9. Description of Haemoproteus stellaris n.sp. and a review of the haemoproteids of the swallow family Hirundinidae. Canadian Journal of Zoology 56(10):2110-2116. https://doi.org/10.1139/z78-286
Whitman, J. S., and N. Wilson. 1992. Incidence of louse-flies (Hippoboscidae) in some Alaskan birds. North American Bird Bander 17(2):65-68.
Winkler, D. W., K. K. Hallinger, T. M. Pegan, C. C. Taff, M. A. Verhoeven, D. C. van Oordt, M. Stager, J. J. Uehling, M. N. Vitousek, M. J. Andesen, et al. 2020. Full lifetime perspective on the costs and benefits of lay-date variation in tree swallows. Ecology 101(9):e03109. https://doi.org/10.1002/ecy.3109
Zamora-Vilchis, I., S. E. Williams, and C. N. Johnson. 2012. Environmental temperature affects prevalence of blood parasites of birds on an elevation gradient: implications for disease in a warming climate. PLoS ONE 7(6):e39208. https://doi.org/10.1371/journal.pone.0039208
Fig. 1
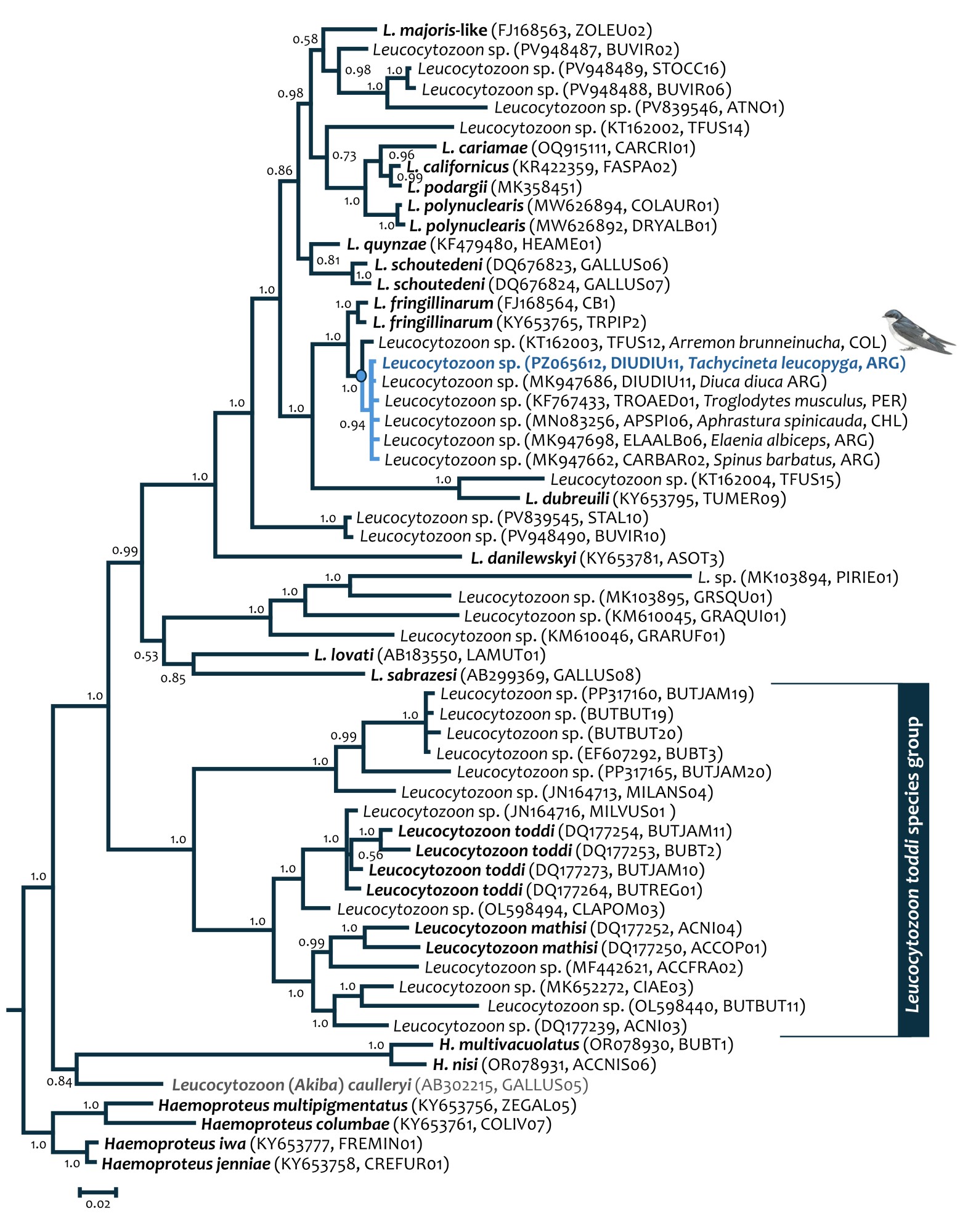
Fig. 1. Phylogenetic relationships of Leucocytozoon sp. DIUDIU11, showing its common ancestry with the TROAED01, APSPI06, ELAALB06, CARBAR02, and TFUS12 lineages, all from South America.
Table 1
Table 1. Prevalence of hemoparasites in Tachycineta species. Genus-level prevalence is shown where available. Abbreviations: N = sample size, H = Haemoproteus, P = Plasmodium, L = Leucocytozoon, T = Trypanosoma.
| Species | % prevalence (N) | Parasite tested | Parasite positive | Location (latitude, longitude) | Reference | ||||
| Tree Swallow (T. bicolor) |
0% (3) | P | Chicago, USA (41° N, 87° W) | Medeiros et al. (2015) | |||||
| 40% (5) T 80% (5) L |
H P L T | L T |
Colorado, USA (38° N, 106° W) | Murdock (2009) | |||||
| 0% (12) | H P | H P | Colorado, USA (40° N, 106° W) | Rodriguez et al. (2021) | |||||
| 0.45% (221) | H P L T | T | Saskatoon, Canada (52° N, 106° W) | Shutler et al. (2003) | |||||
| 19% (1562) all 7.9% (125) P 9.9% (155) L 0% H |
H P L T | P L |
Québec, Canada (52° N, 72° W) | Turcotte et al. (2018) | |||||
| 48% (89, sequenced 22) | H P | H P | New York, USA (40° N, 74° W) | Szymanski and Lovette (2005) | |||||
| 50% (2) | H P L | Mixed infection with H and P | Michigan, USA (44° N, 85° W) | Smith et al. (2018) | |||||
| Violet-green Swallow (T. thalassina) | 100% (1) | H P L T | Mixed infection by L and H | Colorado, USA (38° N, 106° W) | Murdock (2009) | ||||
| 71.4% (7) | H P L | H | California, USA (39° N, 123° W) | Martinsen et al. (2008) | |||||
| 88.9% (9) | H P L T microfilaria | H L T and microfilaria | Colorado, USA (40° N, 106° W) | Stabler and Kitzmiller (1970) | |||||
| White-rumped Swallow (T. leucorrhoa) | No information | H P | H | Uruguay (30-35° S, 53-58° W) | Durrant et al. (2006) | ||||
| Chilean Swallow (T. leucopyga) |
0% (26) | H P L | Chile (33-53° S, 70-72° W) | Merino et al. (2008) | |||||
| 0% (106) | H P L | Bariloche (41° S, 71° W) | Current study | ||||||
| 2.6% (38) | H P L | L | Tierra del Fuego (54° S, 68° W) | Current study | |||||
Table 2
Table 2. Estimates of evolutionary divergence between sequences. The number of base substitutions per site from between sequences are shown. Standard error estimate(s) are shown above the diagonal. Analyses were conducted using the Tajima-Nei model (Tajima and Nei 1984). The rate variation among sites was modeled with a gamma distribution (shape parameter = 1). The differences in the composition bias among sequences were considered in evolutionary comparisons (Tamura and Kumar 2002). The analysis involved 7 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated. There were a total of 477 positions in the final dataset. Evolutionary analyses were conducted in MEGA7 (Kumar et al. 2016).
| Leucocytozoon sp. (ID, lineage, host, country) | Genetic distance (standard error) | ||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| 1 | Leucocytozoon sp. (Sample 3499, DIUDIU11, Tachycineta leucopyga, ARG) | 0.000 | 0.002 | 0.002 | 0.002 | 0.002 | 0.004 | ||
| 2 | Leucocytozoon sp. (MK947686, DIUDIU11, Diuca diuca, ARG) | 0.000 | 0.002 | 0.002 | 0.002 | 0.002 | 0.004 | ||
| 3 | Leucocytozoon sp. (KF767433, TROAED01, Troglodytes musculus, PER) | 0.002 | 0.002 | 0.003 | 0.003 | 0.003 | 0.004 | ||
| 4 | Leucocytozoon sp. (MN083256, APSPI06, Aphrastura spinicauda, CHL) | 0.002 | 0.002 | 0.004 | 0.003 | 0.003 | 0.004 | ||
| 5 | Leucocytozoon sp. (MK947698, ELAALB06, Elaenia albiceps, ARG) | 0.002 | 0.002 | 0.004 | 0.004 | 0.003 | 0.004 | ||
| 6 | Leucocytozoon sp. (MK947662, CARBAR02, Spinus barbatus, ARG) | 0.002 | 0.002 | 0.004 | 0.004 | 0.004 | 0.004 | ||
| 7 | Leucocytozoon sp. (KT162003, TFUS12, Arremon brunneinucha, COL) | 0.006 | 0.006 | 0.008 | 0.008 | 0.008 | 0.008 | ||