







The following is the established format for referencing this article:
Valine, A., D. Alexander, A. Bracey, S. R. Kolbe, and A. Grinde. 2026. Demographic patterns and habitat associations of non-breeding Northern Shrikes (Lanius borealis) in Minnesota. Journal of Field Ornithology 97(2):3.ABSTRACT
Despite its ecological importance as a predatory passerine, the Northern Shrike (Lanius borealis) remains severely understudied. In North America, this species breeds in the boreal taiga and taiga-tundra ecotone and winters across much of the northern continental United States and southern Canada. Because of its remote breeding range, population estimation for this species is challenging, and little is known about its population structure, movement, or habitat use. However, wintering populations in Minnesota provide a unique opportunity to study Northern Shrikes’ non-breeding ecology, as the birds are more accessible, and their habitat preferences are easier to observe during this time. To fill these gaps in knowledge, we (1) analyzed morphometric and demographic data on Northern Shrikes (n = 58) banded in northeastern Minnesota between 2021 and 2024, (2) deployed light-level geolocators (n = 31) on non-breeding Northern Shrikes between 2021 and 2022, and (3) identified land cover preferences throughout Minnesota based on 1185 Northern Shrike eBird observations. We found a significantly higher number of adults present during the winter. Adult males averaged significantly larger than adult females by wing chord and tail length, and adults were significantly longer winged than juveniles. A combination of poor non-breeding site fidelity and near-zero recapture rates prevented the recovery of any geolocator units. Using resource selection functions, we identified that Northern Shrikes used grassland, water, wetland, and developed land in greater proportion to what was available on the landscape, but selected against cropland and forest. Northern Shrikes also preferentially selected for areas with a greater number of land cover types present, indicating a potential preference for heterogeneous landscapes. Our results provide the first description of Northern Shrike demographics, tracking technology deployment, and large-scale non-breeding habitat associations for this species in Minnesota.
RESUMEN
A pesar de su importancia ecológica como paseriforme depredador, Lanius borealis sigue siendo una especie escasamente estudiada. En América del Norte, esta especie se reproduce en la taiga boreal y en el ecotono taiga-tundra, y pasa el invierno en gran parte del norte de Estados Unidos continental y del sur de Canadá. Debido a su remota área de reproducción, la estimación de la población de esta especie resulta difícil, y se conoce poco sobre su estructura poblacional, movimientos o uso del hábitat. Sin embargo, las poblaciones invernantes en Minnesota ofrecen una oportunidad singular para estudiar la ecología no reproductiva de L. borealis, ya que los individuos son más accesibles y sus preferencias de hábitat son más fáciles de observar durante este periodo. Para llenar estos vacíos de conocimiento, (1) analizamos datos morfométricos y demográficos de L. borealis (n = 58) anillados en el noreste de Minnesota entre 2021 y 2024, (2) colocamos geolocalizadores basados en niveles de luz (n = 31) en individuos no reproductivos entre 2021 y 2022, y (3) identificamos las preferencias de cobertura del suelo en todo Minnesota a partir de 1185 observaciones de eBird de la especie. Encontramos un número significativamente mayor de adultos presentes durante el invierno. Los machos adultos fueron, en promedio, significativamente más grandes que las hembras adultas en cuanto a la cuerda alar y la longitud de la cola, y los adultos mostraron alas significativamente más largas que los juveniles. Una combinación de baja fidelidad al sitio durante el período no reproductivo y tasas de recaptura cercanas a cero impidió la recuperación de algún geolocalizador. Mediante funciones de selección de recursos, identificamos que L. borealis utilizó pastizales, cuerpos de agua, humedales y áreas desarrolladas en una proporción mayor que la disponible en el paisaje, pero seleccionó negativamente cultivos y bosques. L. borealis también seleccionó preferentemente áreas con un mayor número de tipos de cobertura de suelo, lo que indica una posible preferencia por paisajes heterogéneos. Nuestros resultados proporcionan la primera descripción de la demografía, implementación de tecnología de seguimiento y asociaciones de hábitat a gran escala durante el período no reproductivo de esta especie en Minnesota.
INTRODUCTION
The Northern Shrike (Lanius borealis) is a predatory passerine with a unique ecological niche, often referred to as a “butcher bird” because of its habit of impaling prey on thorns or barbed wire in a larder. This behavior is a key adaptation for hunting in cold, resource-scarce environments (Cade and Swem 1995, Atkinson 1997). The species specializes in hunting small mammals and insects but will also take small birds, making it a top avian predator in its non-breeding habitat. The Northern Shrike breeds in remote areas in the taiga and taiga-tundra ecotone and occurs in low densities across the non-breeding range in the continental United States and Canada (Paruk et al. 2020). This species is relatively understudied (Paruk et al. 2020), with notable prior studies focusing on breeding biology (Cade and Swem 1995), winter territory selection (Atkinson 1993), hunting techniques (Atkinson 1997), and winter diet (Atkinson and Cade 1993). Assessing population trends is difficult for this species (Paruk et al. 2020) because of inherent limitations of survey methods such as the Breeding Bird Survey (Ziolkowski et al. 2023) and Christmas Bird Count (National Audubon Society 2025) and is further confounded by apparent cyclic movements in eastern North America (Davis 1937, Hess 2000). Despite these limitations, current population estimates for Northern Shrike suggest a yearly decline of between 1.3% (1965–2003, Niven et al. 2004) and 1.67% (1993–2015, Sauer et al. 2017), raising conservation concerns. The drivers of this decline are not well understood but may be linked to habitat loss on both breeding and non-breeding grounds or shifts in prey availability because of a changing climate.
Two distinct Northern Shrike breeding populations occur east and west of Hudson Bay, believed by some (Miller 1931) but not others (Pyle 1997, Ridgeway 1904, as cited in Paruk et al. 2020) to be separate subspecies. Though identification in the field is unreliable (Paruk et al. 2020), both eastern and western individuals are believed to mix during the non-breeding season in the western Great Lakes region (Miller 1931). Northern Shrikes, like many other bird species, may also exhibit differential migration, where age or sex classes migrate at different times, along different routes, or to different non-breeding areas (Cristol et al. 1999). Such patterns could result in demographic groups occupying distinct habitats or regions during the non-breeding season, potentially exposing them to varying environmental pressures or survival risks (Marra and Holmes 2001, Briedis and Bauer 2018). Understanding these patterns is important for interpreting observed age and sex ratios and for identifying factors influencing habitat use and survival during winter. Northern Shrikes are known to occupy discrete territories during the non-breeding season, with the non-breeding territory size reported at approximately 0.5 km² in Idaho (Atkinson 1993). There is also some evidence of non-breeding site fidelity in both this species (Rimmer and Darmstadt 1996, Small 2017) and in the closely related Loggerhead Shrike (Lanius ludovicianus, Donahue et al. 2024), suggesting that territoriality and familiarity with local habitat may interact with differential migration to shape age- and sex-specific distribution patterns during winter.
Minnesota hosts an estimated 7% of the global population of Northern Shrikes during the non-breeding season (Fink et al. 2023) and sits in the assumed mixing zone of eastern and western breeding populations. Prior research on non-breeding Northern Shrikes has primarily occurred in Idaho on the western edge of the species’ range (Atkinson 1993, Atkinson and Cade 1993, Atkinson 1997); therefore, the non-breeding population of the western Great Lakes region is worthy of additional study.
As a low-density species inhabiting northern ecosystems, understanding Northern Shrike population dynamics and habitat requirements is crucial for predicting how they might respond to ongoing ecological changes. Because little is known about Northern Shrike demographics, site fidelity, or habitat use in this region, our objectives were to (1) compare demographic and morphometric data of Northern Shrikes captured during the non-breeding season, (2) investigate non-breeding site fidelity and return rates, and (3) apply resource selection functions (RSFs) using eBird data (eBird 2025) to determine Northern Shrike land cover preferences in Minnesota. We hypothesized that morphometric measurements would align with those reported by Brady et al. (2009) in nearby northwestern Wisconsin. Last, we predicted that non-breeding Northern Shrikes would preferentially select open landscapes such as mesic areas, agricultural fields, and grasslands, consistent with prior data on breeding and non-breeding habitat preferences in other regions (Atkinson 1993, Paruk et al. 2020).
METHODS
Study sites and field methods
Our fieldwork was conducted within a roughly 6000 km² area of northeastern Minnesota in St. Louis, Carlton, Aitkin, and Itasca Counties between January 2021 and February 2024, with birds captured between December and March each winter (Fig. 1). The majority of shrikes were trapped in the Sax-Zim Bog area (47.17°N, 92.65°W; Fig. 1) of St. Louis County, approximately 60 km northwest of Duluth, MN. Others were captured around Palisade, MN in Aitkin County (46.81°N, 93.48°W) and throughout Carlton County (46.51°N, 92.59°W; Fig. 1).
We initially located shrikes by driving repeatedly through areas with known shrike presence (personal observation or eBird reports) or potential shrike habitat (e.g., areas characterized by a mosaic of open spaces, dense shrubs, and tall hunting perches) and attempting to capture any shrikes detected. To find individuals in the years following geolocator deployments, our strategy focused on a combination of intensive and exploratory searches. Resighting surveys of tagged shrikes were conducted via driving all passable roads within a 5 km-radius of the original banding location while carefully scanning available hunting perches and attempting to capture any newly detected shrike. Each site of a geolocator deployment was revisited 5–10 times throughout each winter (between December and March). Additionally, we expanded our search to new areas with suitable habitat to locate both new and returning birds that may have dispersed. These exploratory searches were conducted via driving passable roads within the greater 6000 km² study site, with the most time spent within 15–20 km of geolocator deployment locations. This dual approach of focusing on previously occupied sites while also exploring new territory ensured a comprehensive effort was employed to relocate banded individuals.
Northern Shrikes were captured using a round Potter trap (Craig 1997) or a modified bal-chatri/noose carpet (AV personal design) baited with a live Western European house mouse (Mus musculus domesticus). Each shrike was fitted with a unique United States Geological Survey (USGS) numeric aluminum leg band and, in the winter of 2021–2022, a unique combination of 2 wraparound Darvic color bands. For each bird, we recorded mass (g), wing chord (mm), tail length (mm), and length of black (mm) on the inner vane of the outermost rectrix (r6). Individuals were aged as either adult (definitive cycle basic) or juvenile (first cycle formative) based on molt and other plumage characteristics (Pyle 1997, Wolfe et al. 2010). Adults were sexed by plumage characteristics and validated by the discriminant function equation using tail length and amount of black on r6 developed by Brady et al. (2009).
Northern Shrikes captured in the winters of 2020–2021 (n = 10) and 2021–2022 (n = 21) were fitted with light-level geolocators from Migrate Technology (model Intigeo-W65C2) using a leg-loop harness attachment (Rappole and Tipton 1991). The total weight of the geolocator and harness was 0.8 g, which is 1.2% of the average mass (69 g) of the Northern Shrikes included in our study.
Age and sex ratios and morphometrics
We used goodness-of-fit G-tests to compare capture rates between adults and juveniles and between adult males and adult females. The alpha value for these tests was set to 0.05. We also used two-sample t-tests to compare morphometric measurements (wing chord, tail length, and mass) between adult males and adult females, and between adults and juveniles. We assessed the distribution of each morphometric variable for normality using the Shapiro-Wilk test and by visual inspection of histograms and Q-Q plots. We then tested for variance (F-test) for each variable and found unequal variance in male versus female mass and adult versus juvenile tail length. Where variance was unequal, each group’s variance was estimated separately using the Welch modification to the degrees of freedom. Given the small sample sizes and correlation among morphometric variables, we used a Bonferroni correction, resulting in an alpha value of 0.017.
Resource selection functions
We used a resource selection function (RSF) to assess whether Northern Shrikes preferentially selected specific land cover types during the non-breeding season in Minnesota. Using a use- versus-available RSF (Design I, Type A as described in Manly et al. 2002), we estimated the probability that a Northern Shrike utilizes a given land cover type relative to its availability across the Minnesota landscape.
For used locations, we analyzed data obtained from eBird, a citizen-science project where volunteers report bird observations via “checklists” of species seen along with location (i.e., spatial coordinates) and effort data (i.e., duration of time spent observing birds; Sullivan et al. 2009). We downloaded all Northern Shrike observations in Minnesota through February 2024 but constrained our analysis to reports occurring within the last 10 years (2015–2024) to best align with the land cover data used. We then filtered the data to retain only checklists with the most precise locations possible. Because reporters can choose to submit checklists from predefined larger areas called “hotspots” (e.g., an entire state park or wildlife refuge) or select a “traveling” effort protocol, which may cover a large distance from the reported checklist coordinates, we filtered the data according to the recommended best practices for eBird data (Johnston et al. 2021) to include only checklists submitted from personally selected locations using the “stationary” or “incidental” effort protocols by five or fewer observers. We also filtered temporally to include only observations between November and March to reduce the chance of sampling Northern Shrikes observed during migration stopover. This filtering resulted in 3850 individual observations, many of which were clustered in areas with concentrated birding activity (e.g., Sax-Zim Bog, the Twin Cities, and Duluth). To address this strong spatial sampling bias, we created hexagonal grid cells with 5 km spacing across the state of Minnesota and randomly sampled one Northern Shrike observation from each cell according to the recommended eBird best practices (Johnston et al. 2021) using the dggridR (Barnes and Sahr 2024) and auk packages (Strimas-Mackey et al. 2023) in R (v.4.2.2, R Core Team 2024). The total filtering and subsampling resulted in 1185 “used” locations (Fig. 2A). Of the available 5 km grid cells spanning Minnesota, 83% did not contain any eBird reports of Northern Shrike. Although this percentage reflects the expected gaps in citizen science coverage and the species’ natural density, we emphasize that our goal was not to model occupancy or abundance, but to use the grid-based subsampling to de-cluster the data. This ensured that over-sampled locations (e.g., birding hotspots) did not disproportionately influence the habitat signatures in our models. By thinning the data to one observation per 5 km, we maintained a dataset more representative of the diverse land cover types utilized by shrikes across the state’s broader geography.
We found that the vast majority of Northern Shrikes reported via eBird were plotted along roads. Many birders observe birds by car, particularly during the winter season when roads provide easy access to otherwise inaccessible regions and vehicles provide shelter from extreme winter temperatures. Although roadsides are not the sole habitat for Northern Shrikes, they are effective for surveying because the linear infrastructure of utility wires and fences provides continuous perches, which potentially provide foraging opportunities (O'Brien and Ritchison 2011, Crouch et al. 2019). To address this inherent sampling bias, we constrained our “available” locations for the RSF analysis to the surveyed roads. This approach allowed us to model habitat selection from the perspective of a shrike using this linear feature, ensuring that our model reflects the actual choices birds are making within the available road-based landscape. Using ArcGIS Pro version 3.2 (Environmental Systems Research Institute 2023), we generated 500,000 random points on the Minnesota public road system and subsampled these using the same methods used for the eBird Northern Shrike reports (i.e., one point per 5 km hexagonal grid cell). Because of the constraints of the road system, we generated 6649 non-overlapping “available” points (Fig. 2B), resulting in a roughly 1:6 ratio of used to available locations for the RSF. This is slightly lower than the 1:10 ratio recommended by Nad’o and Kaňuch (2018) but was the highest ratio possible given our stated constraints.
In ArcGIS, we generated buffers around each used and available point, using the smallest non-breeding territory size of Northern Shrike reported in Idaho (0.554 km²) by Atkinson (1993) as an approximation of the area most likely to be used by an individual non-breeding shrike. We calculated the proportion of 15 different land cover types within each buffer using the 2021 National Land Cover Database (NLCD: available at https://www.mrlc.gov/data; Fig. 2D). The NLCD has a fundamental cell size (spatial resolution) of 30 × 30 meters, providing detailed coverage of habitat composition across the study area. These proportions were used as predictor variables in our generalized linear mixed models (GLMMs) to assess Northern Shrike habitat use. We chose GLMMs to account for spatial variation across Minnesota, using the four major ecosystem provinces (Laurentian Mixed Forest, Eastern Broadleaf Forest, Prairie Parklands, and Tallgrass Aspen Parklands; Aaseng et al. 2011; Fig. 2C) as a random effect. We aggregated the 15 NLCD land cover types (Table 1: NLCD Classifications) into seven broader categories (Table 1: Variable) and used the relative proportion of each land cover type as predictor variables. We chose to classify “hay pasture” as grassland habitat rather than cropland because both land cover types lack tree canopy and are characterized by low-lying grassy vegetation. During the winter, this results in similar habitat structures and snow depths, providing functionally similar habitat for Northern Shrikes. Similarly, because barren land in Minnesota is predominantly open-pit mines, gravel pits, and other human-driven land modifications, we combined the “barren” NLCD category with other developed land.
Initial inspection of the land cover data, consisting of seven continuous predictor variables, revealed low to moderate pairwise correlations (max r = 0.58). However, preliminary multivariate models failed to converge or exhibited signs of multicollinearity, likely because of the inherent constraints of compositional land cover data. Consequently, we analyzed each land cover type in a separate GLMM framework to prioritize model stability and interpretability. Although we recognize that analyzing variables in isolation is not ideal for capturing complex interactions, we believe this approach provides the most reliable estimates given the data structure, and we interpret our results with this limitation in mind. The binary response variable indicated whether a location was used (1) or available (0), and univariate models allowed us to assess the effect of each land cover type individually. Each model produced a selection coefficient (β), with positive values indicating selection for a variable and negative values indicating selection against it; β values farther from zero reflect stronger selection. All univariate models were compared against a null model containing only the random effect (ecosystem province) to determine if resource selection was statistically significant; this comparison was formalized using a Likelihood Ratio Test (LRT). To test the secondary hypothesis that Northern Shrikes preferentially use areas with high land cover diversity, we constructed a separate GLMM. The response variable was the same binomial use/availability variable, and the fixed predictor was the number of land cover categories present (i.e., the count of land cover types with a non-zero proportion in each used or available buffer). The alpha value for each model was set at 0.05. All statistical analyses were performed using R version 4.2.2 (R Core Team 2024) and the lme4 package (Bates et al. 2015).
RESULTS
Age and sex ratios and morphometrics
We banded 58 non-breeding Northern Shrikes between 2021 and 2024, with 10 captured in the winter of 2020–2021, 21 in the winter of 2021–2022, 16 in the winter of 2022–2023, and 11 in the winter of 2023–2024. Variation in numbers reflects differences in sampling effort rather than notable differences in the number of shrikes present. The majority of shrikes were banded in St. Louis County (47) with the remainder in Carlton (7), Aitkin (3), and Itasca (1) Counties.
Of the 58 individuals captured, 45 (78%) were adults. The ratio of juveniles to adults varied somewhat between winters, ranging from 1 of 16 (6%) in 2022–2023 to 8 of 21 (38%) in 2021–2022. In total, adults were captured significantly more often than juveniles (X² = 18.68, P < 0.001). Of the 45 adults banded, there were slightly more males than females (26:19). Adult males outnumbered adult females every winter except 2021–2022, though the total sex ratios were not significantly different (X² = 1.09, P = 0.30).
Results of the two-sample t-test showed that adult males had significantly longer wing chords (t43 = 4.24, P < 0.001) and tails (t42 = 3.19, P = 0.003) than adult females, but there was not a significant difference in mass between the sexes (t26 = 2.00, P = 0.056; Table 2). Adults were generally larger and heavier than juveniles, but only wing chord differed significantly between age classes (t56 = 5.15, P < 0.001; Table 3).
Return rates and site fidelity
We identified two color-banded shrikes occupying the same territories in subsequent winters after capture, but these were the only individuals that definitively returned to the same territories. One shrike banded and tagged in 2022 was reported dead about 70 km south of its previous location one year later, but we were unable to recover its geolocator. Another shrike banded in 2023 (not tagged with a geolocator) was reported dead about 65 km north of its previous location two years later. At least three color-banded shrikes were replaced in the same general territory by a new unbanded shrike in the following winter. The remaining shrikes were not detected again, although it is possible that some of these birds could have been present in the same territories but were not encountered, as individual shrike detectability is typically low.
The two shrikes that were relocated alive in subsequent seasons were trap-shy and evaded capture. We caught all of the tagged shrikes using a potter trap but attempted recapture using a bal-chatri with a noose carpet. Despite being presented with a different trap type, the birds were still suspicious. Only one shrike was recaptured throughout the course of the study, and it was an accidental same-season recapture of a non-geolocator bird banded in January 2023 and recaptured in February of the same year. We failed to recover any geolocators during the two winters following deployment, which we primarily attribute to potential low site fidelity, low detectability, and poor return rates exhibited by the shrikes in the study area.
Resource selection functions
Northern Shrikes exhibited a strong selection (preference) for four of the seven land cover variables (Table 4). The strongest selection was observed for grasslands (β = 1.72, s.e. = 0.21, P < 0.001), followed by water (β = 1.42, s.e. = 0.31, P < 0.001), wetlands (β = 1.02, s.e. = 0.14, P < 0.001), and developed areas (β = 1.00, s.e. = 0.15, P < 0.001). All four of these single-predictor models demonstrated a significantly better fit to the data than the null model (LRT P < 0.001). Conversely, Northern Shrikes showed avoidance (selection against) for two land cover types: croplands (β = -1.47, s.e. = 0.13, P < 0.001) and forests (β = -1.13, s.e. = 0.17, P < 0.001). Both models also represented a statistically significant improvement over the mull model (LRT P < 0.001). The association with shrublands was not significantly different from the null model (LRT P = 0.15). The relative abundance of land cover types within the total available habitat buffers for cropland, wetland, forest, and developed were 40%, 19%, 18%, and 12%, respectively (Fig. 3). In contrast, the most abundant cover types within the used buffers were cropland, wetland, developed, and forest (25%, 23%, 18%, and 15%, respectively; Fig. 3). Although cropland and forest were among the most abundant land cover types within used buffers (25% and 15%, respectively), their proportions were lower than expected relative to their availability, resulting in the calculated negative selection coefficients for these habitats.
Available locations had an average of 4.1 land cover types occurring within the buffers, while the used locations had an average of 4.6 land cover types within the buffers. The GLMM assessing land cover diversity showed that Northern Shrikes preferred areas with a higher number of land cover types represented (β = 0.22, s.e. = 0.03 [95% CI: 0.17, 0.28], P < 0.001).
DISCUSSION
Our study of Northern Shrikes in the western Great Lakes region provides insight into the demographics and habitat use of a poorly studied species in a crucial portion of its non-breeding range. We confirmed that the morphometric data of Northern Shrikes in Minnesota generally align with findings from nearby Wisconsin, supporting our hypothesis and suggesting regional consistency in key physical characteristics. However, our ability to investigate population dynamics and site fidelity was limited by the challenges inherent in studying a low-density species with low site fidelity. The observed low return rates and difficulties in recapturing individuals equipped with geolocators highlights the need for alternative tracking technologies or more extensive long-term studies to fully understand the movements and population connectivity of this species during the non-breeding season. Our resource selection function analysis provides valuable new information on habitat preferences, revealing that non-breeding shrikes in Minnesota actively select for open landscapes with scattered perching opportunities, a finding consistent with their known ecological niche and foraging behaviors. These results collectively contribute to a more comprehensive understanding of Northern Shrike ecology in a region that is a suspected mixing zone for different breeding populations.
Demographics and morphometrics
Differential migration, in which age or sex classes move at different times, along different routes, or to different non-breeding areas, has been documented in many bird species (Cristol et al. 1999). Members of different demographic classes may winter in different regions or in different habitats because of variation in migratory timing or dominance status (e.g., Marra and Holmes 2001). When age or sex classes are separated geographically, this can lead to disparities in survival through vulnerability of demographic groups to different environmental disturbances (Briedis and Bauer 2018).
In our study, the majority of non-breeding Northern Shrikes were adults, with juveniles composing a smaller proportion that varied between winters. This adult-biased age ratio may reflect differential migration, where adults arrive earlier and secure higher-quality non-breeding territories, potentially forcing juveniles to travel farther or occupy lower-quality habitat. Shorter migrations may benefit adults energetically and allow earlier return to breeding grounds to reestablish territories (Kokko 1999). Other demographic processes may also contribute to this pattern, including higher mortality rates for juveniles or lower reproductive output for adults, which could influence the age structure of the population and have implications for overall population declines. Because our study area is at the northern edge of the species’ non-breeding range, many juveniles may overwinter farther south, potentially in southern Minnesota or northern Iowa, while adults remain farther north. Evidence from Loggerhead Shrikes indicates that differential habitat selection or use varies based on migratory status or demographic class (Pérez and Hobson 2009, Donahue et al. 2024), supporting the idea that similar age- or sex-related habitat patterns could occur in Northern Shrikes.
Sex ratios among adults were slightly male-biased, and adult males tended to have larger wing and tail measurements than females, while mass differences were less pronounced. Adults were generally larger than juveniles, consistent with age-related growth patterns. Our morphometric results align with prior studies in the western Great Lakes region (Brady et al. 2009) and are intermediate between measurements from the eastern and western extremes of the species’ range (Paruk et al. 2020). These findings support the hypothesis that individuals from both eastern and western breeding populations mix on the non-breeding grounds and suggest that differential migration, territorial dominance, and habitat quality together shape age- and sex-specific patterns of distribution and survival during the non-breeding season.
Return rates and site fidelity
We believed Northern Shrikes to be good candidates for geolocator technology because they are known to occupy discrete non-breeding territories on the wintering grounds (Atkinson 1993), and there is some prior evidence of reasonably high non-breeding site fidelity in both this species (Rimmer and Darmstadt 1996, Small 2017) and in the closely related Loggerhead Shrike (Donahue et al. 2024). However, we were unable to retrieve any geolocators over the two winters following deployment, primarily because the shrikes did not show high fidelity to previously held non-breeding territories. Site fidelity is often poorly or vaguely defined, sometimes referring to fidelity to a broader region, and other times referring to fidelity to a particular territory. Additionally, most studies of site fidelity focus on breeding sites because fewer species maintain discrete non-breeding territories. Unlike breeding site fidelity, which appears to be influenced in some species by previous breeding success at a given site (e.g., Hoover 2003), it is more difficult to determine which factors drive an individual to choose to return or not return to a non-breeding site. The presence of high-quality habitat and abundant prey are likely important considerations as animals make movement decisions during the non-breeding season. Rimmer and Darmstadt (1996) examined non-breeding site fidelity of Northern Shrikes using band recovery data; however, because of the 10-minute block precision level of banding locations submitted to the USGS Bird Banding Lab, they were unable to determine whether shrikes encountered within the same 10-minute block were occupying the same non-breeding territory or were up to 38 km away. Thus, they ascribed high site fidelity to shrikes up to 60 km from their original banding locations. Although this type of fidelity to a general region is notable, we suggest defining a distinction between territory fidelity (returning to the same or very nearly the same territory) and regional fidelity (returning to a general area, but not the same territory).
Our results suggest that non-breeding territory fidelity in Northern Shrikes does occur in northeastern Minnesota but may not be particularly common. Other individuals may choose different non-breeding territories in the same general region, as was the case with two of our banded birds that wintered in territories > 65 km away in following winters. We found high re-occupancy rates by new shrikes in locations where we had previously banded others, suggesting that even areas of high-quality habitat with sufficient prey availability are not sought out by the same individual across multiple non-breeding seasons. However, because Northern Shrikes can be difficult to detect even across multiple attempts and color bands can be difficult to resight, there is potential for previously banded birds to return to the same territory but go undetected. Additional study is needed to determine the prevalence of both territory and regional non-breeding fidelity in this species.
Habitat use
We found that Northern Shrikes preferred grassland, water, wetland, and developed land but selected against forest and cropland. Overall, they selected areas with a larger number of different land cover types present on the landscape, indicating a preference for a heterogeneous mosaic of land cover. Although the effect size for this preference was relatively small, it aligns with patterns observed in closely related Loggerhead Shrikes, which select for vegetative heterogeneity (Prescott and Collister 1993, Michaels and Cully 1998) and for developed areas, water, and rights-of-way for hunting (Collister and Wilson 2007, Donahue et al. 2021, 2024, Maddox and Hill 2024). This comparison supports the idea that predatory shrikes may benefit from accessing a range of land cover types that provide both hunting perches and cover.
The preference for grassland and wetland is intuitive given Northern Shrike hunting behavior. They are primarily sit-and-wait predators, using elevated perches to watch for prey moving on the ground, though they will occasionally pursue prey into dense cover or hover systematically over open areas (Paruk et al. 2020). Open habitats like grasslands and herbaceous wetlands interspersed with trees or snags allow shrikes to survey large areas efficiently. Shrubs and woody wetlands provide necessary cover for roosting, shelter from harsh winter weather and predators, prey habitat, and facilitate behaviors such as acoustical luring (Atkinson 1993, 1997). These patterns are consistent with previous studies in Idaho, where Northern Shrikes were strongly associated with mesic and riparian habitats that offered hunting perches and dense cover for shelter and foraging (Atkinson 1993).
Northern Shrikes appear to prefer heterogeneous landscapes with multiple, varied land cover types rather than more homogeneous areas such as monoculture agriculture or extensive forest tracts. Notably, they also showed a preference for developed land, consistent with our observations of shrikes hunting along roadsides, near small farms, and within urban patches. Small mammals often occur at higher densities along road rights-of-way compared to adjacent habitats (Adams and Geis 1983), and raptor species are known to exploit these areas despite associated risks (Meunier et al. 2000, Planillo et al. 2015). Northern Shrikes likely use similar strategies, benefiting from concentrated prey and readily available hunting perches. Overall, these findings highlight that Northern Shrikes utilize a diverse set of habitats across the non-breeding landscape, reflecting a flexible habitat use strategy that may be important for their survival and persistence in a variable and human-modified environment. Despite the inherent limitations in our methods (e.g., estimating territory size based on single-point observations), our results align with patterns reported from other locations and for closely related species, lending credence to the usefulness of citizen-science datasets like eBird for investigating ecological questions.
CONCLUSION
Our results underscore that Northern Shrike habitat use is complex and influenced by multiple, interacting factors. Future research should continue to investigate population structure, migratory connectivity, demographics across the non-breeding range, and potential drivers of population decline, including contrasting patterns between eastern and western populations. Given the challenges of recapturing individuals and the low fidelity to non-breeding territories observed in this study, geolocators may not be well-suited for studying this species during the non-breeding season. Emerging tracking technologies, such as smaller, more accurate cellular or satellite transmitters, and tools like the Motus network for migration and non-breeding movements, may provide more precise insights into breeding origins, migration, and site fidelity. For studies that require recapture, trialing multiple trap types will likely be critical. Additionally, incorporating spatial metrics or experimentally testing which combinations of land cover types optimize for foraging efficiency, roosting safety, or thermoregulation, as well as examining landscape-level habitat heterogeneity, including spatial configuration and diversity indices, could offer valuable insight into fine-scale habitat preferences and inform effective conservation and management strategies for Northern Shrikes.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
We thank the numerous volunteers who assisted with winter fieldwork, particularly David Valine and Hannah Toutonghi. We thank Frank Nicoletti and Hawk Ridge Bird Observatory for providing banding supplies, permitting, and overall project support. We also thank Ryan Brady for sharing his shrike expertise during project conceptualization, and the three anonymous reviewers whose comments greatly improved the manuscript. Funding for the geolocators was provided by generous grants from the Minnesota Ornithologists’ Union and the Friends of Sax-Zim Bog.
DATA AVAILABILITY
The data for this study are available at the Data Repository for the University of Minnesota: https://hdl.handle.net/11299/280001.
LITERATURE CITED
Aaseng, N. E., J. C. Almendinger, R. P. Dana, D. S. Hanson, M. D. Lee, E. R. Rowe, K. A. Rusterholz, and D. S. Wovcha. 2011. Minnesota’s native plant community classification: a statewide classification of terrestrial and wetland vegetation based on numerical analysis of plot data. Biological Report No. 108. Minnesota County Biological Survey, Ecological Land Classification Program, and Natural Heritage and Nongame Research Program. Minnesota Department of Natural Resources, St. Paul, Minnesota, USA.
Adams, L. W., and A. D. Geis. 1983. Effects of roads on small mammals. Journal of Applied Ecology 20(2):403-415. https://doi.org/10.2307/2403516
Atkinson, E. C. 1993. Winter territories and night roosts of Northern Shrikes in Idaho. Condor 95(3):515-527. https://doi.org/10.2307/1369595
Atkinson, E. C. 1997. Singing for your supper: acoustical luring of avian prey by Northern Shrikes. Condor 99(1):203-206. https://doi.org/10.2307/1370239
Atkinson, E. C., and T. J. Cade. 1993. Winter foraging and diet composition of Northern Shrikes in Idaho. Condor 95(3):528-535. https://doi.org/10.2307/1369596
Barnes, R., and K. Sahr. 2024. dggridR: Discrete Global Grids. R package version 3.1.0. https://doi.org/10.32614/CRAN.package.dggridR
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1):1-48. https://doi.org/10.18637/jss.v067.i01
Brady, R. S., J. D. Paruk, and A. J. Kern. 2009. Sexing adult Northern Shrikes using DNA, morphometrics, and plumage. Journal of Field Ornithology 80(2):198-205. https://doi.org/10.1111/j.1557-9263.2009.00209.x
Briedis, M., and S. Bauer. 2018. Migratory connectivity in the context of differential migration. Biology Letters 14(12):20180679. https://doi.org/10.1098/rsbl.2018.0679
Cade, T. J., and T. Swem. 1995. Ecology of Northern Shrikes nesting in arctic Alaska. Proceedings of the Western Foundation of Vertebrate Zoology 6(1):204-214.
Collister, D. M., and S. Wilson. 2007. Territory size and foraging habitat of Loggerhead Shrikes (Lanius ludovicianus) in southeastern Alberta. Journal of Raptor Research 41(2):130-138. https://doi.org/10.3356/0892-1016(2007)41[130:TSAFHO]2.0.CO;2
Craig, S. H. 1997. What goes around-gets caught!! An improved trap for shrikes. North American Bird Bander 22:124-125.
Cristol, D. A., M. B. Baker, and C. Carbone. 1999. Differential migration revisited: latitudinal segregation by age and sex class. Pages 33-88 in V. Nolan, E. D. Ketterson, and C. F. Thompson, editors. Current Ornithology. Springer, New York, New York, USA. https://doi.org/10.1007/978-1-4757-4901-4_2
Crouch, C. G., A. J. Flores, A. Krainyk, L. A. Brennan, D. B. Wester, E. D. Grahmann, R. H. Benson, F. Hernández, and J. F. Kelly. 2019. Comparative habitat use of wintering American Kestrels and Loggerhead Shrikes along south Texas roadways. Southeastern Naturalist 18(2):240-255. https://doi.org/10.1656/058.018.0203
Davis, D. E. 1937. A cycle in Northern Shrike emigrations. Auk 54(1):43-49. https://doi.org/10.2307/4078322
Donahue, E. R., K. J. Krajcir, L. C. Bryant, R. Raibley, J. L. Wessels, J. Youtz, and T. J. Boves. 2021. Non-breeding behavior and diet of Loggerhead Shrikes in an intensive agricultural region. Southeastern Naturalist 20(3):427-447. https://doi.org/10.1656/058.020.0304
Donahue, E. R., A. J. Worm, J. L. Wessels, J. Youtz, L. C. Bryant, A. E. Matthews, K. J. Krajcir, R. Raibley, and T. J. Boves. 2024. Evaluating demographic habitat partitioning and its consequences during the non-breeding season in Loggerhead Shrikes. Journal of Field Ornithology 95(2):1. https://doi.org/10.5751/JFO-00454-950201
eBird. 2025. eBird: An online database of bird distribution and abundance [web application]. eBird, Cornell Lab of Ornithology, Ithaca, New York, USA. https://ebird.org/
Environmental Systems Research Institute. 2023. ArcGIS Pro, Version 3.2.2. Environmental Systems Research Institute, Redlands, California, USA.
Fink, D., T. Auer, A. Johnston, M. Strimas-Mackey, S. Ligocki, O. Robinson, W. Hochachka, L. Jaromczyk, C. Crowley, K. Dunham, et al. 2023. eBird Status and trends, data version: 2022; released: 2023. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/ebirdst.2022
Hess, P. 2000. Pennsylvania’s record high Northern Shrike invasion of 1999-2000. Pennsylvania Birds 14(1):2-10.
Hoover, J. P. 2003. Decision rules for site fidelity in a migratory bird, the Prothonotary Warbler. Ecology 84(2):416-430. https://doi.org/10.1890/0012-9658(2003)084[0416:DRFSFI]2.0.CO;2
Johnston, A., W. Hochachka, M. Strimas-Mackey, V. R. Gutierrez, O. Robinson, E. Miller, T. Auer, S. Kelling, and D. Fink. 2021. Analytical guidelines to increase the value of community science data: an example using eBird data to estimate species distributions. Diversity and Distributions 27(7):1265-1277. https://doi.org/10.1111/ddi.13271
Kokko, H. 1999. Competition for early arrival in migratory birds. Journal of Animal Ecology 68(5):940-950. https://doi.org/10.1046/j.1365-2656.1999.00343.x
Maddox, K. A., and C. E. Hill. 2024. Use of space by urban Loggerhead Shrikes (Lanius ludovicianus) as a window into habitat suitability. Journal of Field Ornithology 95(2):6. https://doi.org/10.5751/JFO-00458-950206
Manly, B. F. J., L. McDonald, D. L. Thomas, T. L. McDonald, and W. P. Erickson. 2002. Resource selection by animals: statistical design and analysis for field studies. Second edition. Springer Science + Business Media, Berlin, Germany.
Marra, P. P., and R. T. Holmes. 2001. Consequences of dominance-mediated habitat segregation in American Redstarts during the nonbreeding season. Auk 118(1):92-104. https://doi.org/10.1093/auk/118.1.92
Meunier, F. D., C. Verheyden, and P. Jouventin. 2000. Use of roadsides by diurnal raptors in agricultural landscapes. Biological Conservation 92(3):291-298. https://doi.org/10.1016/S0006-3207(99)00094-4
Michaels, H. L., and J. F. Cully Jr. 1998. Landscape and fine scale habitat associations of the Loggerhead Shrike. Wilson Bulletin 110(4):474-482.
Miller, A. H. 1931. Systematic revision and natural history of the American shrikes (Lanius). University of California Publications in Zoology 38(2):11-242.
Nad’o, L., and P. Kaňuch. 2018. Why sampling ratio matters: logistic regression and studies of habitat use. PLoS ONE 13(7):e0200742. https://doi.org/10.1371/journal.pone.0200742
National Audubon Society. 2025. Audubon Christmas Bird Count. https://www.audubon.org/community-science/christmas-bird-count
Niven, D. K., J. R. Sauer, G. S. Butcher, and W. A. Link. 2004. Christmas Bird Count provides insights into population change in land birds that breed in the boreal forest. American Birds 58:10-20.
O’Brien, E., and G. Ritchison. 2011. Non-breeding ecology of Loggerhead Shrikes in Kentucky. Wilson Journal of Ornithology 123(2):360-366. https://doi.org/10.1676/10-146.1
Paruk, J. D., T. J. Cade, E. C. Atkinson, P. Pyle, and M. A. Patten. 2020. Northern Shrike (Lanius borealis), version 1.0. In S. M. Billerman, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.norshr4.01
Pérez, G. E., and K. A. Hobson. 2009. Winter habitat use by Loggerhead Shrikes (Lanius ludovicianus) in Mexico: separating migrants from residents using stable isotopes. Journal of Ornithology 150(2):459-467. https://doi.org/10.1007/s10336-008-0364-0
Planillo, A., S. Kramer-Schadt, and J. E. Malo. 2015. Transport infrastructure shapes foraging habitat in a raptor community. PLoS One 10(3):e0118604. https://doi.org/10.1371/journal.pone.0118604
Prescott, D. R. C., and D. M. Collister. 1993. Characteristics of occupied and unoccupied Loggerhead Shrike territories in southeastern Alberta. Journal of Wildlife Management 57(2):346-352. https://doi.org/10.2307/3809432
Pyle, P. 1997. Identification guide to North American birds: part I: Columbidae to Ploceidae. Slate Creek Press, Point Rayes Station, California, USA.
R Core Team. 2024. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://doi.org/10.32614/R.manuals
Rappole, J. H., and A. R. Tipton. 1991. New harness design for attachment of radio transmitters to small passerines. Journal of Field Ornithology 62(3):335-337.
Rimmer, C. C., and C. H. Darmstadt. 1996. Non-breeding site fidelity in Northern Shrikes. Journal of Field Ornithology 67(3):360-366.
Sauer, J. R., K. L. Pardieck, D. J. Ziolkowski Jr, A. C. Smith, M. A. R. Hudson, V. Rodriguez, H. Berlanga, D. K. Niven, and W. A. Link. 2017. The first 50 years of the North American Breeding Bird Survey. Condor 119(3):576-593. https://doi.org/10.1650/CONDOR-17-83.1
Small, D. M. 2017. Winter site fidelity and over-wintering site persistence of a Northern Shrike, Lanius borealis, in Maryland. Maryland Birdlife 66(2):9-19.
Strimas-Mackey, M., E. Miller, and W. Hochachka. 2023. Auk: eBird data extraction and processing in R. R package version 0.7.0. https://doi.org/10.32614/CRAN.package.auk
Sullivan, B. L., C. L. Wood, M. J. Iliff, R. E. Bonney, D. Fink, and S. Kelling. 2009. eBird: a citizen-based bird observation network in the biological sciences. Biological Conservation 142(10):2282-2292. https://doi.org/10.1016/j.biocon.2009.05.006
Wolfe, J. D., T. B. Ryder, and P. Pyle. 2010. Using molt cycles to categorize the age of tropical birds: an integrative new system. Journal of Field Ornithology 81(2):186-194. https://doi.org/10.1111/j.1557-9263.2010.00276.x
Ziolkowski Jr., D. J., M. Lutmerding, W. B. English, V. I. Aponte, and M. A. R. Hudson. 2023. North American Breeding Bird Survey Dataset 1966-2022: U.S. Geological Survey data release. https://doi.org/10.5066/P9GS9K64
Fig. 1
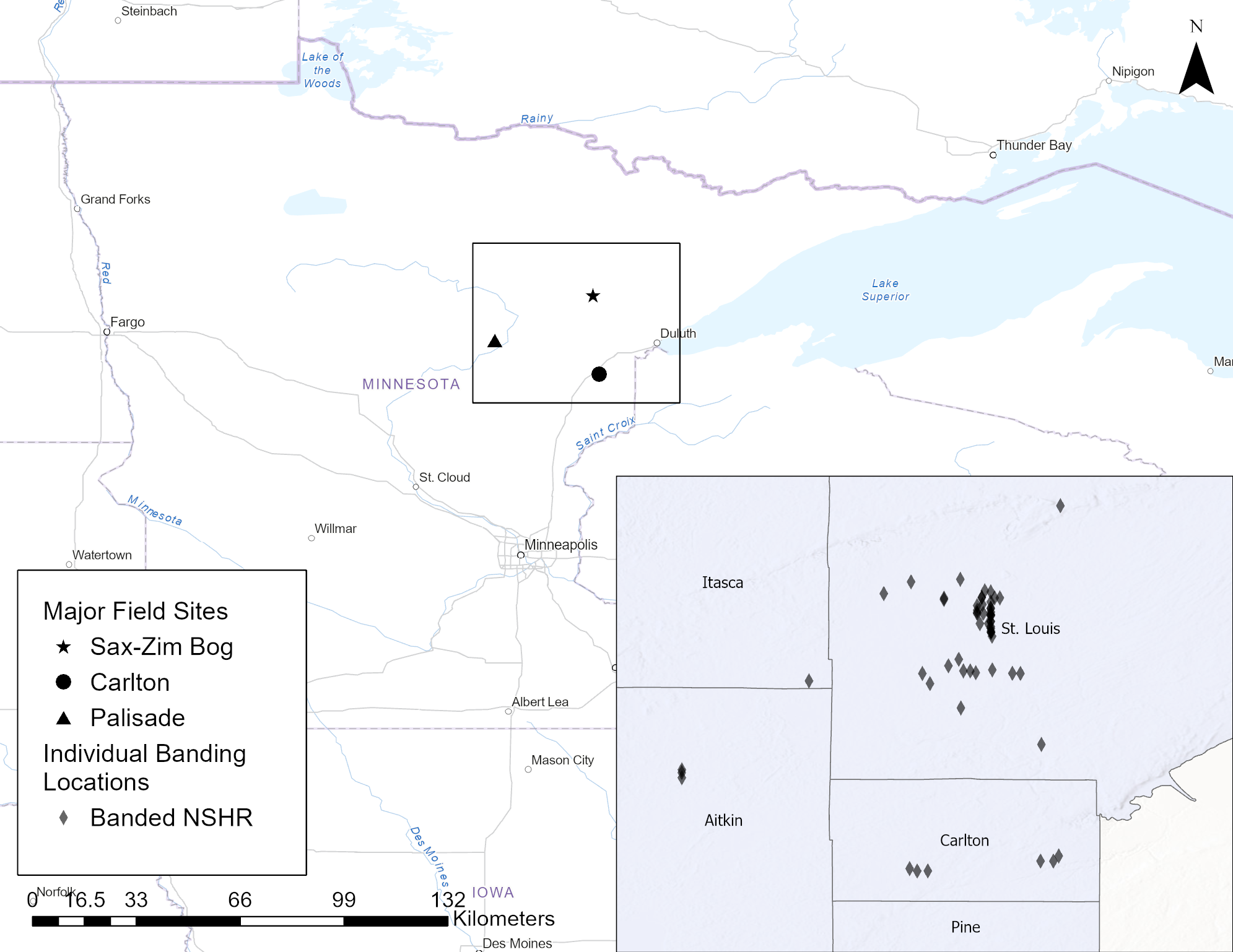
Fig. 1. Location of major field sites (Sax-Zim Bog, Carlton, and Palisade) where Northern Shrikes (Lanius borealis) were captured during the non-breeding season (December–March) in Minnesota between 2021 and 2024. Inset shows the locations where individual shrikes were captured and banded (Banded NSHR) and Minnesota county boundaries.
Fig. 2
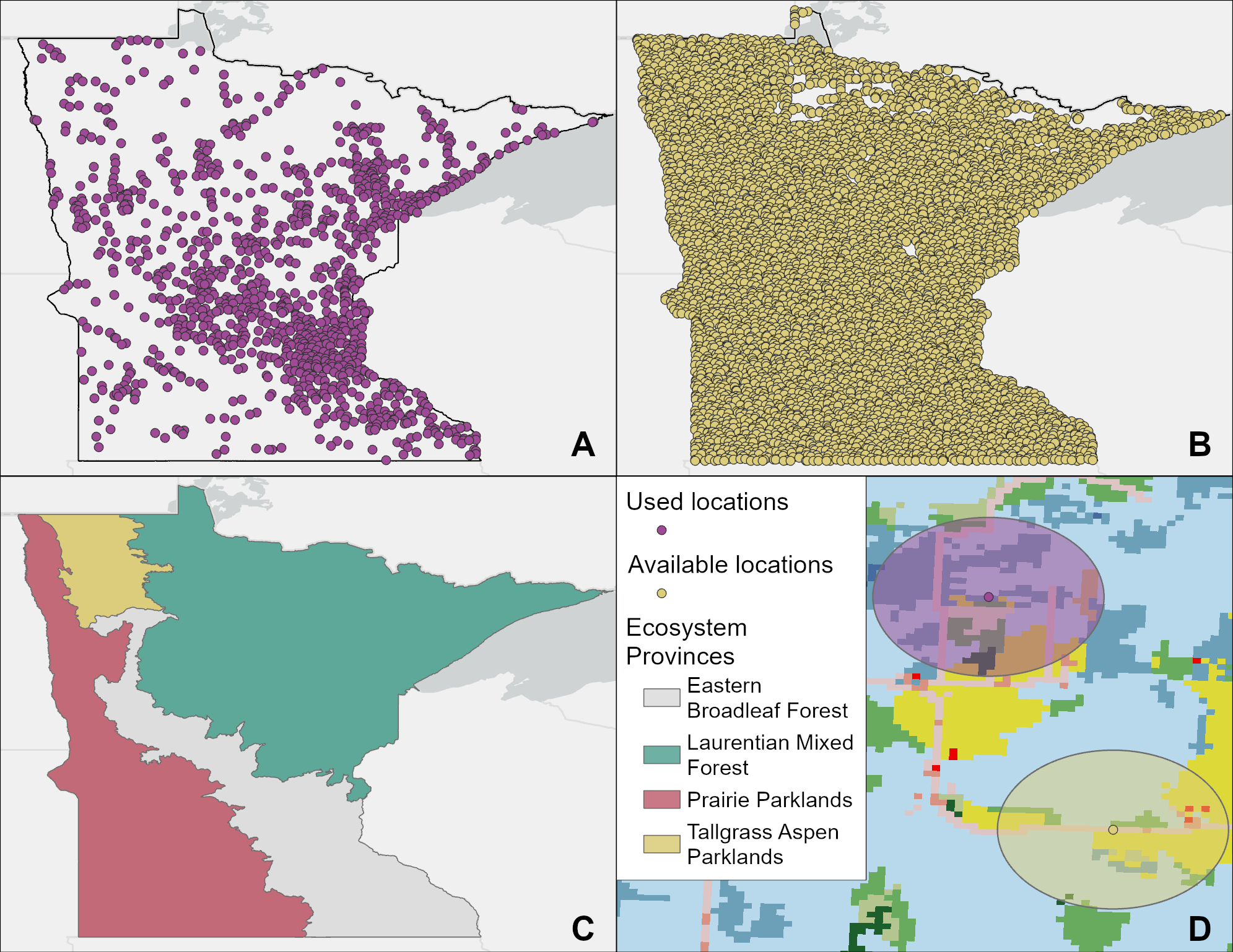
Fig. 2. Spatial data used in the study. (A) 1185 “used” Northern Shrike (Lanius borealis) locations subsampled from eBird data, (B) 6649 randomly generated “available” locations, (C) Ecosystem provinces of Minnesota, and (D) Example of used and available buffers (0.554 km²) containing National Land Cover Database (NLCD) land cover categories. Colors in panel D illustrate different land cover classes and are not listed in the legend; see Table 1 for a full list of NLCD categories and the aggregation used in the analysis.
Fig. 3
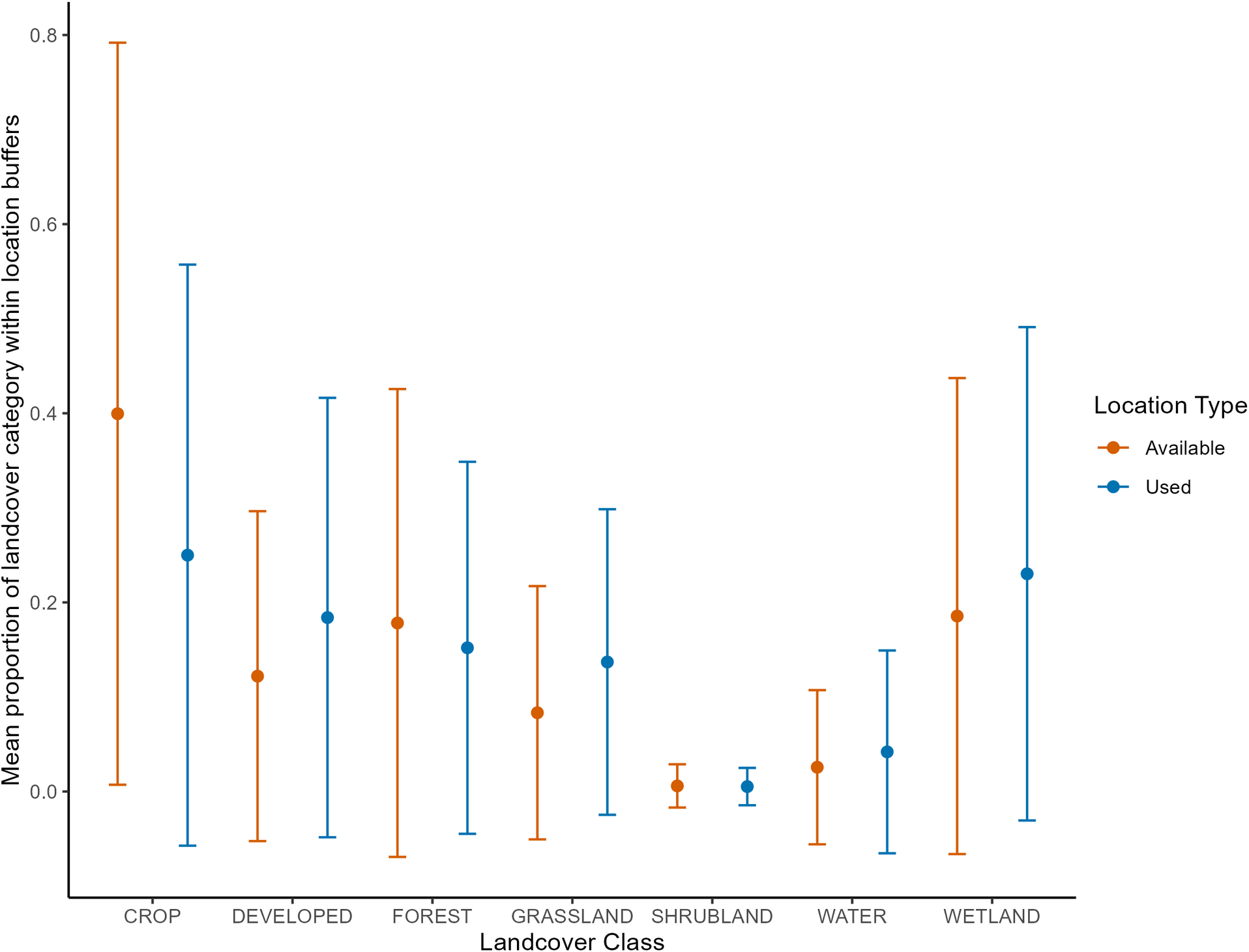
Fig. 3. Mean proportion of land cover categories found within 0.554 km² buffers around used and available locations of Northern Shrike (Lanius borealis) in Minnesota between 2015 and 2024. Error bars indicate standard deviations.
Table 1
Table 1. Hypothesized effects of land cover on Northern Shrike (Lanius borealis) habitat selection during the non-breeding season. Land cover categories were aggregated from the 15 National Land Cover Database (NLCD) classifications used in the resource selection function analysis.
| NLCD land cover variable | Hypothesized effect on Northern Shrike selection | ||||||||
| Water (Open Water) | Select. Northern Shrikes will preferentially select areas near open water because these habitats support high densities of prey such as amphibians and aquatic insects (Cade and Swem 1995, Atkinson 1997). | ||||||||
| Wetland (Woody Wetlands, Emergent Herbaceous Wetlands) | Select. Northern Shrikes will select wetland habitats, as these areas support high densities of small vertebrate and invertebrate prey, including amphibians, rodents, and insects (Cade and Swem 1995, Atkinson 1997). | ||||||||
| Shrubland (Shrub-Scrub) | Select. Northern Shrikes will select shrub-dominated areas because shrubs provide hunting perches and elevated vantage points to detect prey (Cade and Swem 1995, Atkinson 1997). | ||||||||
| Grassland (Herbaceous, Hay Pasture) | Select. Northern Shrikes will preferentially use grasslands, particularly where vegetation is relatively tall or heterogeneous, providing suitable cover for prey while maintaining visibility for hunting (Cade and Swem 1995, Paruk et al. 2020). | ||||||||
| Developed (Open Space, Low, Medium, and High Intensity, Barren) | Select. Northern Shrikes will select areas with low- to medium-intensity development (e.g., open spaces, suburban areas) because of the presence of perches for hunting and moderate prey availability (Small 2017, Donahue et al. 2024). | ||||||||
| Forest (Deciduous, Coniferous, Mixed) | Avoid. Northern Shrikes will avoid dense forested areas, as closed-canopy forests limit visibility and reduce foraging efficiency for this sit-and-wait predator (Atkinson 1993, Paruk et al. 2020). | ||||||||
| Cropland (Cultivated Crops) | Avoid. Northern Shrikes will avoid cultivated croplands because of reduced prey availability and frequent human disturbance (Atkinson and Cade 1993, Brady et al. 2009). | ||||||||
Table 2
Table 2. Comparison by sex of wing chord, tail length, and mass of adult Northern Shrikes (Lanius borealis) captured during the non-breeding season (2021–2024) in Minnesota.
| Variable | Males (n = 26) | Females (n = 19) | P-value (t-test) | ||||||
| Wing chord (mm) | 116.3 ± 1.6 | 114.4 ± 1.3 | < 0.001 | ||||||
| Range: 111–120 | Range: 111–116 | ||||||||
| Tail length (mm) | 110.4 ± 1.9 | 108.3 ± 2.4 | 0.003 | ||||||
| Range: 106–114 | Range: 101–113 | ||||||||
| Mass (g) | 71.1 ± 3.0 | 68.3 ± 5.5 | 0.056 | ||||||
| Range: 67–80 | Range: 59–80 | ||||||||
Table 3
Table 3. Comparison by age class of wing chord, tail length, and mass of adult and juvenile Northern Shrikes (Lanius borealis) captured during the non-breeding season (2021–2024) in Minnesota.
| Variable | Adults (n = 45) | Juveniles (n = 13) | P-value (t-test) | ||||||
| Wing chord (mm) | 115.5 ± 1.7 | 112.8 ± 1.6 | < 0.001 | ||||||
| Range: 111–120 | Range: 110–116 | ||||||||
| Tail length (mm) | 109.5 ± 2.4 | 106.9 ± 4.2 | 0.054 | ||||||
| Range: 101–114 | Range: 101–116 | ||||||||
| Mass (g) | 69.9 ± 4.4 | 68.4 ± 3.6 | 0.250 | ||||||
| Range: 59–80 | Range: 62–74 | ||||||||
Table 4
Table 4. Selection coefficients (β) ± standard error, 95% confidence intervals, and P-values calculated from binomial generalized linear mixed models of Northern Shrike (Lanius borealis) land cover use in Minnesota between 2015 and 2024.
| Land cover | Selection coefficient (β) | 2.5% CI | 97.5% CI | P-value | |||||
| Grassland | 1.72 ± 0.21 | 1.32 | 2.12 | < 0.001 | |||||
| Water | 1.42 ± 0.31 | 0.80 | 2.03 | < 0.001 | |||||
| Wetland | 1.02 ± 0.14 | 0.74 | 1.30 | < 0.001 | |||||
| Developed | 1.00 ± 0.15 | 0.71 | 1.28 | < 0.001 | |||||
| Shrubland | -2.29 ± 1.60 | -5.79 | 0.78 | 0.15 | |||||
| Forest | -1.13 ± 0.17 | -1.47 | -0.80 | < 0.001 | |||||
| Cropland | -1.47 ± 0.13 | -1.72 | -1.22 | < 0.001 | |||||