







The following is the established format for referencing this article:
Grinde, A. R., S. R. Kolbe, K. G. Snow, B. S. Howland, G. Niemi, R. A. Slesak, and M. A. Windmuller-Campione. 2023. Habitat use and characteristics of Connecticut Warbler during the nesting and post-fledging period. Journal of Field Ornithology 94(4):2.ABSTRACT
Widespread declines in breeding bird populations have been documented across North America since the 1970s, and concerns about loss of avian biodiversity are growing. Species with narrow habitat preferences are of particular conservation concern. Connecticut Warblers (Oporornis agilis) breed in forests in central Canada and around the western Great Lakes; throughout much of its breeding range, this species is associated with lowland black spruce (Picea mariana) and tamarack (Larix laricina) forests. Population trends of the Connecticut Warbler indicate it is one of the most rapidly declining bird species in North America, but the species is understudied because it occurs in relatively low densities across its breeding range. To better understand the breeding ecology of the Connecticut Warbler, we studied its nesting and post-fledging habitat use and survival in northern Minnesota, USA at two study areas in 2019 and 2020. We mapped territories of 49 singing males, located and monitored 11 nests, and tracked the post-fledging movements of individuals from five broods. Nest sites were located in lowland conifer stands with a semi-open canopy and dense understory. The average fledging age was 7.5 days post-hatch, and the individuals (n = 14) tracked during 0–7 days post-fledging had a mean daily distance from nests of 35.5 m and a maximum distance from nests of 104 m. Connecticut Warblers were not observed making movements of more than 100 m from the nest until 7 days post-fledging. Microsite areas with high stem density were important features for post-fledgling birds, and that the same habitats were used for breeding and the post-fledging period. The results of this study can be used by managers to develop conservation strategies that will provide critical habitat to support this species.
RESUMEN
Las disminuciones generalizadas en las poblaciones de las aves reproductivas se han documentado a través de Norte América desde 1970 y las preocupaciones sobre la pérdida de biodiversidad aviar han incrementado. Las especies con preferencias de hábitat restringidas son de particular preocupación para la conservación. Oporornis agilis, se reproduce en los bosques de Canadá central y alrededor de los Grandes Lagos; a lo largo de gran parte de su distribución reproductiva, esta especie está asociada con bosques bajos dominados por Picea mariana y Larix laricina. Las tendencias poblacionales de Oporornis agilis indican que es una de las especies de aves que ha disminuido más rápidamente en Norte América, pero la especie es poco estudiada por que ocurre en densidades relativamente bajas a través de su rango de reproducción. Para entender mejor la ecología reproductiva de Oporornis agilis, estudiamos su hábitat de anidación y post-volantones y la supervivencia en el norte de Minnesota, EE. UU. en dos áreas de estudio en 2019 y 2020. Mapeamos los territorios de 49 adultos cantores, localizamos y monitoreamos 11 nidos y rastreamos los movimientos posteriores a la salida del nido en individuos de cinco camadas. Los sitios de anidación estuvieron localizados en bosques de tierras bajas de coníferas con un dosel semi abierto y un sotobosque denso. El promedio de edad en la que los pichones abandonaron el nido fue 7.5 días después del nacimiento, y los individuos (n=14) rastreados durante 0-7 días después del abandono del nido tuvieron una distancia diaria media del nido de 35.5. m y una máxima distancia del nido de 104 m. No observamos individuos de Oporornis agilis realizando movimientos de más de 100 m del nido si no, hasta después de los 7 días posteriores al abandono del nido. Las áreas de microhábitat con alta densidad de ramas fueron características importantes para las aves después del abandono, y que los mismos hábitats fueron utilizados para la reproducción y el periodo posterior al abandono del nido. Los resultados de este estudio pueden ser usados para desarrollar estrategias que proporcionen un hábitat crítico para soportar esta especie.
INTRODUCTION
Widespread declines in breeding bird populations have been documented across North America since the 1970s, and concerns about loss of avian biodiversity are growing (Sekercioglu et al. 2004, IUCN 2019). Habitat loss and degradation associated with land management and climate change appear to be primary factors driving bird population declines (Matricardi et al. 2020, Betts et al. 2022). For example, fragmentation and simplification of forest habitat structural and floristic diversity associated with past and present forest management practices have impacted forest understory-nesting and area-sensitive birds (Alverson et al. 1988, Robinson and Wilcove 1994, Donovan et al. 1995, Betts et al. 2022). It is estimated that 98% of boreal forest bird species are either moderately or highly vulnerable to climate change (Rosenberg et al. 2019, Wilsey et al. 2019) and that ranges of some forest bird species will shift (or continue to shift), while other species may disappear entirely due to a lack of required habitat components (Hitch and Leberg 2007, Zuckerberg et al. 2009, Wilsey et al. 2019). Boreal conifer forests may be particularly vulnerable to climate change and are expected to decrease in extent as temperatures rise (LaChance et al. 2005, Handler et al. 2014, Stralberg et al. 2015).
The Connecticut Warbler (Oporornis agilis), a neotropical migrant that winters in South America and breeds in boreal forests of North America, is one of the rarest and most narrowly distributed wood warblers in eastern North America (Ridgely et al. 1989, Paynter 1995, Pitocchelli et al. 2020, Hallworth et al. 2021). Its breeding range stretches across a narrow band of the Canadian boreal forest from western Quebec to eastern British Columbia, and in the United States, is restricted to the northern Great Lakes states, specifically, northern Minnesota, northern Wisconsin, and the Upper Peninsula of Michigan (Payne 1983, Janssen 1987, Niemi et al. 2016, Pitocchelli et al. 2020, Pfannmuller et al. 2023). Long-term, widespread population declines in the breeding populations of Connecticut Warbler have been documented throughout the species’ range. For example, long-term monitoring data in Minnesota show a significant decrease in the population, averaging −8.65% per year since 1995 (Grinde et al. 2022, Pfannmuller et al. 2023). Declines in population coupled with a restricted breeding range have prompted a variety of state and federal agencies to designate the Connecticut Warbler as a species of conservation concern. For example, Partners in Flight (2021) identified the species as one of 12 Yellow Watch List species, a designation reserved for species that “require constant care.” While multiple factors are likely attributable to the species– population decline, including tower collisions and loss of wintering habitat (Rappole 1995, Arnold and Zink 2011, Longcore et al. 2013, Loss et al. 2014), quality and quantity of breeding habitat are also key factors (Hallworth et al. 2021).
Breeding Connecticut Warblers are ground nesters that are most abundant in mature, lowland coniferous forests composed of widely scattered black spruce (Picea mariana) and tamarack (Larix laricina) trees, but are also found in jack pine (Pinus banksiana) stands and in young, regenerating conifer stands, and they breed in upland aspen forests (Populus spp.) in some parts of their range (Elder 1991, Kirk et al. 1996, Kudell-Ekstrum 2002, Niemi et al. 2016). Recent research has focused on the distribution, population trends, and human impact on breeding habitat of Connecticut Warblers in the boreal forest (Sodhi and Paszkowski 1995, Lapin et al. 2013, Zlonis et al. 2017, Hallworth et al. 2021). For example, studies have indicated that Connecticut Warblers demonstrate a preference for large, unfragmented landscapes of both upland coniferous and lowland black spruce forests and breeding habitats that are characterized by a high density of ground cover and low canopy cover, features that are typical of sparsely forested peatlands (Lapin et al. 2013, Pfannmuller et al. 2023). These studies have provided important information to better understand the Connecticut Warbler’s breeding habitat associations, but characteristics associated with breeding territories, nest placement, and fledgling behavior remain largely unknown. To address these deficiencies, we evaluated nest site characteristics, nestling success, and post-fledging movements at two study areas in northern Minnesota. Our specific objectives were to describe (1) nesting locations and microhabitat site characteristics, and (2) post-fledging movements and habitat use of fledging Connecticut Warblers.
METHODS
Study area
The study areas were located in northern Minnesota, USA (Fig. 1). This region lies within a glacial moraine landscape and is dominated by aspen (Populus spp.) and pine (Pinus spp.) uplands with lowland conifer forests occupying small shallow basins and large peatland complexes. In Minnesota, forested peatlands are concentrated in the Agassiz Lowlands and Tamarack Lowlands subsections (Fig. 1) (MN DNR 2019), and are dominated by black spruce, tamarack, and northern white cedar (Thuja occidentalis). We identified two study areas that represented preferred breeding habitat in Minnesota and had known breeding territories (Pfannmuller et al. 2017, Zlonis et al. 2017, Hallworth et al. 2021). Both study areas were dominated by black spruce and tamarack; however, the proportions of each tree species differed between each of the study areas. The first was located in the Sax-Zim Bog (hereafter Sax-Zim), which is in the Tamarack Lowlands subsection and is located near the southern edge of the Connecticut Warbler breeding range in Minnesota (Fig. 1). The Sax-Zim study area was approximately 150 ha and had 20% black spruce and 80% tamarack. The second study area was located in the Red Lake Wildlife Management Area (hereafter Red Lake) in the Agassiz Lowlands subsection of the state. The Red Lake study area was approximately 60 ha and had 100% tamarack. Landscape composition surrounding the study areas was similar; each was surrounded by a matrix of black spruce, tamarack, and upland aspen.
Nest searching and monitoring
We conducted surveys throughout the two study areas to locate singing Connecticut Warblers. Surveys were completed during the first week of June 2019 in the Sax-Zim study area and the last week of May 2020 in the Red Lake study area (Svenson and Williamson 1969). When a singing male was located, its position was marked using a handheld global positioning system unit (GPS) (Garmin GPSMAP 64st). Areas where Connecticut Warblers were repeatedly singing were visited every 2–3 days to determine whether females were present. When a female was detected, nest searching efforts were focused in those areas. Nest searching was conducted daily from early June to early July in 2019 and 2020. Nests were located using systematic searches and behavioral cues (Hanski et al. 1996, Rodewald 2004, Mattsson and Niemi 2006).
Based on the results of the 2019 efforts at the Sax-Zim study area and to increase the sample size of nests in the 2020 field season, we used targeted mist netting to catch female Connecticut Warblers and attached radio transmitters to track them to nests during the incubation period (Rodewald 2004). Nets were set up approximately 10 m from an area where a female had recently been detected. We also placed a portable speaker on the base of the net and used playback recordings of a Connecticut Warbler call note (https://www.xeno-canto.org/181815). When a female was detected, a person walked toward the net in an attempt to move the female in the direction of the net. Females tended to walk primarily on the forest floor, and were rarely seen but could be heard chipping. Once the female was close to the net, the person rushed the net to flush the female into the net.
Nests were monitored every 2–4 days until the nest failed or nestlings fledged. Once the nest timing was known (i.e., lay or hatch day documented), we minimized the number of nest visits to those necessary during periods around hatching or fledging. At each visit, we noted the presence and location of adults, adult behavior, and the presence and number of eggs or nestlings (Martin and Geupel 1993, Rodewald 2004). We approached and departed the nest from a different direction for each nest check to minimize disturbance and to prevent leaving a trail to the nest (Martin and Geupel 1993, Hanski et al. 1996).
Fledgling ecology
Nestlings were banded with a USGS metal band on the left leg and a unique combination of plastic, UV-resistant, Darvic bands (Avinet, Dryden, NY) on the right leg of each individual approximately 6 days post-hatch (day of pinbreak; 2–3 days prior to fledging). We randomly chose one to three nestlings from each brood and attached VHF radio transmitters (Nanotag, NTQB2-2, Lotek Wireless, Inc.) to them using an elastic thread, figure-eight harness design modified from Rappole and Tipton (1991). Harness material was 0.5-mm diameter, white, thin, beading cord elastic (Prym Consumer, Spartanburg, SC). The transmitter and harness weighed less than 3% of the birds’ body mass, and harnesses were attached to allow for additional growth after attachment.
We tracked each radio-tagged bird once daily using a handheld receiver (SRX800, Lotek Wireless, Inc.) with a three-element Yagi antenna. We searched for individuals each morning in a random order that was developed a priori to reduce differences in locations that may be associated with time of day. Once a bird was located and resighted, we used a handheld GPS to mark its location, and we returned to the location at a later date to conduct vegetation surveys in order to reduce disturbance to the birds. Fledgling locations were recorded at the point where birds were first observed, and birds were located daily unless weather prevented tracking or until there was fledgling mortality (e.g., exposure, predation), transmitter loss (transmitter found and bird was seen alive), or we failed to detect the bird (i.e., it had potentially left the study area). In the event of mortality, we recovered the body and transmitter when possible and attempted to infer cause of mortality, including looking for evidence of predation (e.g., kinked antenna, potential bite marks). Analysis of daily movement patterns was restricted to morning tracking periods (07:00–11:00 CST). We measured distance from the nest and distance moved between observed daily locations using the R package “geodist” (Padgham and Sumner 2021) and ArcGIS Pro 2.7.0 (ESRI, Redlands, CA).
Vegetation surveys
The vegetation sampling protocol used was a modified version of the methods described by James and Shugart (1970) and the Breeding Biology Research and Monitoring Database (BBIRD) program field protocol (Martin et al. 1997). At each nest and fledgling location, we conducted detailed vegetation surveys to characterize stand and microhabitat features of the study areas. Habitat features were measured using an 11.3-m radius circular plot (400 m2) at bird locations (bird plots) and at paired randomly selected locations (random plots) to compare preferences in microsite features. Random plots were located 30 m from bird plots using a random azimuth; random azimuths were generated from spinning a compass for a random amount of time.
At each bird location and random point within a circular plot, we recorded all canopy trees to species and sorted them into a size class of small (8–23 cm diameter at breast height [DBH]), medium (23–38 cm DBH), or large (> 38 cm DBH). Canopy openness was approximated using a densiometer, and average canopy height was estimated by choosing a tree that was representative of the average canopy height and measuring its height using a clinometer. Within the 11.3-m radius plot, a 5-m radius circular plot (78.5 m2) was used to survey ground cover. In the 5-m circular plot, woody stems that were more than 0.5 m in height were counted and identified to species. We also recorded the percent cover of green vegetation, logs, bare ground, mud, leaf litter, sphagnum moss, and water. Percent green vegetation was further divided into the following categories: grass, sedge, reed, shrub, forb, fern, and (non-sphagnum) moss. We measured moss depth by inserting a ruler into the layer of moss until it met firm resistance.
Additional vegetation data were collected at each nest location. Specifically, we documented the dominant and (if applicable) subdominant nest substrate where the nest was placed. We also documented the plant species that concealed the nest; the plant was deemed to be concealing if it covered any portion of the cup of the nest. Nest cover was determined by an observer positioning themselves 1 m directly above the nest, and at eye level from each of the four cardinal directions from the nest, and estimating the percentage of the nest that was covered, to the nearest 5%. Measurements taken of each nest included the inside and outside diameter and inside and outside height.
We summarized the microhabitat features associated with fledgling dispersal by considering plots as experimental units and vegetation variables as response variables. Using the lme function in the nlme package (Pinheiro et al. 2021) in R (R Development Core Team 2022), we compared the difference between the bird plots and random plots for each pair of vegetation measurements. If the intercept was significantly different from zero, we interpreted the vegetation variable to be different between the bird plots and random plots. A unique identification number for each brood was used as a random effect in all models to account for variation.
RESULTS
Overview
During the 2019 and 2020 breeding seasons, we found 11 nests (Sax-Zim n = 2; Red Lake n = 9), attached transmitters on 13 fledglings (Sax-Zim n = 4; Red Lake n = 9) from six broods (Sax-Zim n = 2; Red Lake n = 4), and compared 48 paired (used and random) vegetation surveys (Sax-Zim n = 14; Red Lake n = 34) within fledging dispersal locations.
Nests
At the Sax-Zim study area, 24 singing male Connecticut Warblers were detected within a 122-ha area. At the Red Lake study area, 25 singing males were observed in a 47-ha area. Two nests were found during the 2019 breeding season at the Sax-Zim study area. One of the nests was found on 16 June 2019 and was in incubation stage with four eggs; three eggs hatched. The second nest was found 3–4 days after hatching; it contained four nestlings. The estimated hatch date for both nests was between 30 June 2019 and 1 July 2019. In 2019, the observed nestling period was 7 days for both nests, and all nestlings (n = 7) successfully fledged (Table 1).
Nine nests were found during the 2020 breeding season at the Red Lake study area (Table 1). Two nests (RL1 and RL2) were found early in the season (the first week of June)—one while the female was still laying and the other in the incubation stage; both nests were abandoned. Two other nests were found during the incubation stage (RL5 and RL7), and the remaining five were found during the nestling stage (Table 1). The mean estimated hatch date was June 28 (range = 14 June–11 July), the observed nestling period was approximately 9 days (range = 8–10 days), and the mean fledge date was July 5 (range = 22 June–18 July). In 2020, for the nests found in incubation or nestling stage, 30 of the 31 nestlings successfully fledged (Table 1). Overall, the mean number of nestlings hatched per nest was 4 (range = 3–5 nestlings), and the average survival rate for the nestling-to-fledgling stage was 97% (n = 9 nests). We were able to mist net and tag one female in 2020 and used telemetry to find the nest (RL2); the remaining 10 nests were found using behavioral or systematic methods.
Nest dimensions and substrate
Eleven nests were found, but two of the nests (both in the Red Lake study area) were not intact when we returned to measure the dimensions. The average inside diameter of the nest cups of the nine nests was 6.5 cm (SD = 0.9), mean inside height was 4.6 cm (SD = 1.1), and mean outside diameter was 8.3 cm (SD = 0.9). All nests were located in moss, with the exception of one (SZ1) that was in grass; subdominant substrates included grass and Labrador tea (Rhododendron groenlandicum) (Fig. 2). Concealment features also included grass, Labrador tea, bog birch (Betula pumila), tamarack, and moss. Mean percentage of nest cover (i.e., percentage of the nest that was obscured by the vegetation) in the Red Lake study area was 83% (range = 49–95% cover, n = 7), but two nests in the Sax-Zim study area had only 12 and 19% cover.
Nest microhabitats
The dominant cover type of the nest vegetation survey plots was tamarack at 10 of the 11 nests, and one nest (RL1) was in an area where black spruce was the dominant tree species. In the Sax-Zim study area, the average overstory canopy cover was 46.0% (SD = 10.3; range = 35.8–56.3) and the average height of overstory trees was 13.5 m (SD = 1.5 m; range = 12.0–15.0). The most common tree species and size class was small (8–23 cm DBH) tamarack, with an average of 60 trees (SD = 12; 1500/ha). One nest location also had five small black spruce in the 11.3-m plot (125/ha). There were no snags or medium-sized trees in the Sax-Zim nest plots. The average overstory canopy cover of 50-m plots in the Red Lake study area was 56.4% (SD = 10.9; range = 44.3–82.0), and the average height of overstory trees was 17.5 m (SD = 7.8 m; range = 6.5–32.0). The most common tree species and size class on nest sites was small (8–23 cm DBH) tamarack, with an average of 47 trees (SD = 15; 1175/ha) in the 11.3-m plot, followed by small snags (mean = 20, SD = 6.5; 500/ha), and small black spruce (mean = 43, SD = 40; 1075/ha). An average of 5 (SD = 3.7) medium-sized (23–38 cm DBH; 125/ha) snags were found on the plots in the Red Lake study area, and there were no medium or large trees on any of the nest vegetation plots.
The average woody stem count (which included all woody species over 0.5 m in height) within 5-m nest vegetation plots at the Sax-Zim study area was 400 (SD = 219, range = 181–619; 50,955 stems per hectare), and 10 different types of plants were documented in the nest plots (Appendix 1). The average woody stem count at the Red Lake study area nest plots was 473 (SD = 326, range = 97–964; 60,256 stems per hectare), and 17 different types of plants were documented in the nest plots (Appendix 1). The most common species at the Sax-Zim nest sites were leatherleaf (Chamaedaphne calyculata) and Labrador tea; the most common species at the Red Lake study area was Labrador tea, followed by bog birch (Appendix 1). The average moss depth at nest site vegetation plots was greater in the Red Lake study area (10.1 cm; SD = 2.1) than in the Sax-Zim study area (6.4 cm; SD = 2.2). Overall, the percentage of ground cover at the 5-m scale was similar between the two study areas, and the major cover types were shrub (mean = 47, SD = 31), moss (mean = 21, SD = 15), and grass (mean = 18, SD = 12.5).
Fledgling ecology
Five nestlings from the two nests in the Sax-Zim study area in 2019 were banded, and four were fitted with radio transmitters. The average weight of the nestlings at pinbreak (2–3 days before fledging) was 11.7 g (SD = 0.24; range 11.5–12.0 g), average wing chord was 34.2 mm (SD = 2.7; range = 30.0–38.0 mm), and average tarsus length was 19.8 mm (SD = 1.1; range = 18.0–21.0 mm). At the Sax-Zim study area, two of the tagged birds from nest SZ1 died the first day after fledging; the likely cause of mortality was exposure. One of the tagged birds from nest SZ2 was likely predated 7 days after leaving the nest; the other was tracked for 15 days post-fledge, after which we were unable to locate the bird and assumed it left the study area (Fig. 3a).
Fourteen nestlings were banded at the Red Lake study area in 2020; nine birds from four nests [RL3 (1 individual), RL5 (3), RL6 (2), and RL8 (3)] were tagged with radio transmitters. The average weight of the 14 nestlings at pinbreak was 10.6 g (SD = 0.9; range = 9.5–13.5 g), average wing chord was 31.1 mm (SD = 2.8; range = 25.0–37.0 mm), and average tarsus length was 20.8 mm (SD = 0.54; range = 20.0–21.9 mm). At the Red Lake study area, the fate of each individual tagged was as follows: (1) a tagged bird from nest RL3 lost its transmitter 3 days after leaving the nest (Fig. 3b); (2) one of three birds from nest RL5 lost its transmitter before any data were gathered, we removed the transmitter from a second bird from the same nest because we were concerned it was too loose, and a third bird from the same nest was tracked to day 3, after which time we lost the signal and suspected the bird had been predated (Fig. 3c); (3) we tracked one bird from nest RL6 for six days before the transmitter fell off, and the other bird from nest RL6 was tracked for 22 days before it left the study area (Fig. 3d); and (4) one bird from nest RL8 dropped the tag the second day after leaving the nest, a second bird was predated and some of its remains were found nearby, and we decided to remove the tag from the third individual (Fig. 3e).
Overall, we documented an average daily distance moved from nests of 88.5 m (range = 7.6–203.0 m, n = 39) over the entire fledgling period. The average daily distance moved from nests during the fledgling period of 0–7 days was 35.5 m (range = 7.6–104.0 m, n = 19). Connecticut Warblers were not observed making daily movements of more than 100 m from the nest until 7 days post-fledge (Figs. 3 and 4). The average distance traveled from the previous day’s observed location was 53.5 m (range = 7.6–272.4 m, n = 37) and was relatively consistent with a slight increase in distance moved from the previous day’s location as the birds aged (Fig. 4).
Fledgling microhabitats
We compared 48 paired vegetation plots during the 2019 and 2020 breeding seasons to document fledgling microhabitat use. The dominant cover type at all fledging use points was tamarack, the average canopy cover at fledgling vegetation plots was 38% (SD = 29.7, range = 0–86.3), and average overstory tree height was 16.4 m (SD = 5.5, range = 7.5–27.5). The average moss depth was 7.9 cm (SD = 3.3, range = 2.3–14.8) in the bird use sites and was similar at the random plots (mean = 8.3, SD = 4.3, range = 2.6–23.5). Overall, the percentage of ground cover at the 5-m scale was similar between the two study areas, and the major cover types included moss (mean = 35, SD = 24), shrub (mean = 32.5, SD = 28), and grass (mean = 16, SD = 18). Microhabitats where birds were located had significantly higher total stem count than paired random plots, and there were no significant differences in the other vegetation characteristics (Table 2).
DISCUSSION
The Connecticut Warbler has experienced long-term and widespread declines in breeding populations throughout North America (Sauer et al. 2020, Brady et al. 2021, Grinde et al. 2022, Pfannmuller et al. 2023). The reasons for these steep declines are likely multifaceted, but because the Connecticut Warbler is a climate-sensitive species, conservation efforts require focus on the identification and management of current threats to improve the species’ adaptive capacity and facilitate future shifts in distribution (Strahlberg et al. 2019). However, the Connecticut Warbler is one of the least studied species in North America. Data on its basic biology are lacking, and additional information will be needed before informed management recommendations can be made. Our study is the first to empirically quantify the timing of breeding activities, post-fledging behavior, and the species’ habitat use. Additionally, we have provided details on nest microsites and described sites with high densities of breeding territories.
Nesting ecology
Due to the combination of a largely inaccessible breeding range and the challenge of finding nests, there is a lack of knowledge regarding nest characteristics and fledgling ecology. Our targeted mist netting and radio tagging efforts resulted in only one female that we followed to the nest. Nests were found using behavioral cues such as calling by male or female birds, cessation of male singing when approaching near the nest, and parental “broken wing displays.” In general, the nest habitat, locations, and characteristics were similar to those previously reported (e.g., Harrison 1978, Peck and James 1987), which included breeding territories in relatively open forests, and nests on the ground in thick undergrowth and mounds of moss. Overall, high stem density and shrub cover seem to be important factors in choosing nest sites. Common Ericaceous shrub species such as Labrador tea and leatherleaf grow in canopy openings and were often associated with nesting locations, which suggests that areas with a semi-open canopy may be an important feature of breeding habitat for the Connecticut Warbler. During the nestling period, both parents were observed feeding the young 17 days after fledging. However, feeding rates and food brought to the young are difficult to observe because parents approach nests and fledglings by walking the final 10–15 m.To our knowledge, this was the first study to track fledgling Connecticut Warblers. We were able to track 14 individuals from six broods over the 2-year study. Most individuals were tracked for only a short period before their tags fell off or they died. For those that survived, our data provide an initial picture of post-fledging dispersal. Connecticut Warbler fledglings that left the nest stayed relatively close to their nests (e.g., < 100 m) for approximately 18 days post-fledge. By comparison, age-specific maximum daily movements of fledgling Ovenbirds in Minnesota were estimated to be 400 m at 18 days post-fledge (Streby and Anderson 2013). Connecticut Warblers have a short nestling stage (e.g., they fledge 6–9 days after hatch) and are unable to fly the first week out of the nest. Throughout this post-fledging period, young birds disperse from the nest by walking on moss or through thick vegetation. Brood mates were observed near each other (< 20 m away) but were not observed together after leaving the nest. When approached, the young birds stayed in place or attempted to walk away. After this initial period, on day 7–8 post-fledge, the birds attempted to fly but did not move far. They tended to stay on the ground and used the dense shrub layer for cover because, lacking flight, they are vulnerable to predation. This is supported by their preference for areas with high stem densities in the understory compared with random plots. Connecticut Warblers may select their territories based on the availability of cover for nesting and the benefits these habitats provide for successful fledging of their young. For example, the average canopy cover was approximately 55% in our study areas, and most trees counted on the vegetation plots had a DBH of 23 cm or smaller. The semi-open canopy provides sunlight, which enhances a dense understory.
Dropped tags that were recovered had no feathers or blood on them, which is commonly the case when a bird is predated. We suspected that because the birds walked through a thick shrub layer, the tags caught on vegetation and the harnesses slipped or broke off. Based on the high rate of dropped tags, we recommend that harness style attachments should not be placed on Connecticut Warbler nestlings. Alternative methods that allow the tag to fit closer to the body may perform better.
Breeding habitat
Dominant cover types used by the Connecticut Warbler during the breeding season vary throughout the species’ range; the species has been reported in many different forest types ranging from bogs to upland jack-pine barrens (Kudell-Ekstrum 2002, Niemi et al. 2016, Brady et al. 2021, Pitocchelli et al. 2020). Our study showed densities in tamarack–black spruce stands that were similar to those historically reported in northern Minnesota (Nevers et al. 1981, Hanowski and Niemi 1983, Niemi and Hanowski 1984). For instance, Niemi and Hanowski (1984) calculated average Connecticut Warbler densities of 16.8 males/40 ha in black spruce stands and 8.4 males/40 ha in a right-of-way dominated by black spruce. The approximate densities of singing males in our study areas were similar to those in previous reports: 8 males/40 ha in the Sax-Zim study area and 21 males/40 ha in the Red Lake study area. In two contemporary large-scale studies that used random sampling designs of forest cover types in northern Minnesota, Connecticut Warblers were significantly more common in black spruce–tamarack bogs and forests compared with more than 10 other cover types studied (Niemi et al. 2016, Pfannmuller et al. 2017). There are limited population density data in the United States or Canada to compare with these data; however, Erskine (1977) reported a density of 5 males/km2 in three tamarack stands in Ontario. Elder (1991) found that black spruce and tamarack bogs were the predominant breeding habitats used by the Connecticut Warbler in Ontario. A comprehensive assessment and comparison of Connecticut Warbler breeding habitat and densities across the species’ breeding range is necessary to determine commonalities in breeding habitat features. This would also provide information needed to quantify available habitat, improve species distribution models, and predict future threats to the species.A critical issue for Connecticut Warbler breeding habitat availability, especially in southern regions of in the species’ range, is the threat of climate change. Connecticut Warbler habitats are thought to be particularly vulnerable to a warming climate because bogs and lowland coniferous forests are expected to disappear in their southern range limits (Lachance et al. 2005, Handler et al. 2014, Stralberg et al. 2015). Pfannmuller et al. (2023) noted that climate modeling by the National Audubon Society (Wisely et al. 2019) predicted that a temperature increase of 1.5 °C by 2050 would reduce the acreage of bogs and lowland conifers required by the Connecticut Warbler. If these predictions and scenarios are accurate, they represent potentially dire consequences for the Connecticut Warbler in the continental United States, and potentially further loss of breeding habitat in Canada. It is imperative to understand the dynamics facing the Connecticut Warbler and other species living at the southern edge of their breeding range (Glennon et al. 2019). Importantly, developing and implementing potential conservation strategies related to protecting and increasing available habitat on the southern edge of the breeding range are essential. For example, besides strategies to reduce climate change, management and conservation of Connecticut Warblers require a focus on large tracts of lowland black spruce and tamarack bogs and forests, especially where they are adjacent to upland coniferous forests (Lapin et al. 2013, Niemi et al. 2016, Grinde et al. 2022). Understanding the minimum threshold of suitable Connecticut Warbler breeding habitat that is required for successful breeding is an important step in conservation planning.
Modifying current silvicultural approaches to enhance the quality of breeding habitat is another viable option for conservation efforts. For example, in North America, black spruce and tamarack forests are conventionally managed using even-aged silvicultural systems, often via clearcut with or without reserves, which results in relatively uniform tree sizes. Logging of these forests is often conducted during the stem exclusion stage; this does not allow stands to progress through stand reinitiation, which includes the development of gaps and open habitat required by nesting Connecticut Warblers. Shifting from even-aged silvicultural systems to two-aged systems (e.g., clumped shelterwood with reserve) or uneven-aged systems may provide the within-stand structures that are important features for the species.
CONCLUSION
The combination of future climate change in the north and low adaptive capacity of long-distance migrants may make boreal birds especially vulnerable to extinction in the future (Stralberg et al. 2019). Given that species with declining populations often experience contracting range margins (Lawton 1993, Lenoir and Svenning 2015), there is a need to invest in measures aimed at increasing local populations and preventing further declines via critical habitat protection and efforts to reduce climate change. Basic ecological information is needed before any recommendations or attempts to reverse declines can be made. Understanding the nuances of habitat requirements and how they may change with climate is critical to understanding potential population declines, particularly for species like the Connecticut Warbler. The survival of the Connecticut Warbler will depend on the collection of more data on its basic biology, changes in current projections of our global climate, and optimization of forest management to the species’ benefit.RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
ARG, RAS, and MAWC conceived and established the study design. ARG, SRK, KGS, and BSH designed the data collection methods. ARG, SRK, KGS, and BSH conducted the investigation, and ARG and SRK analyzed the data. ARG, SRK, GJN, RAS, and MAWC wrote the manuscript, and all other authors provided editorial advice and valuable insight. ARG, RAS, and MAWC were involved with funding acquisition.
ACKNOWLEDGMENTS
Thanks to Luke Hoehn, Ean Malchow, Zane Den Oden, Elizabeth Oswald, and Reid Siebers for data collection in the field. We also thank Josh Bednar, Mike Hallworth, and Sparky Stensaas for assisting in site selection. Special thanks to Gretchen Mehmel and Charlie Tucker of the Minnesota Department of Natural Resources for their strong support during all facets of this research, along with housing and logistical support in the Red Lake Wildlife Management Area. Thank you to two anonymous reviewers for their thoughtful feedback on the manuscript.
DATA AVAILABILITY
The data for this study are available at the Data Repository for the University of Minnesota (https://hdl.handle.net/11299/257421).
LITERATURE CITED
Alverson, W. S., D. M. Waller, and S. L. Solheim. 1988. Forests too deer: edge effects in northern Wisconsin. Conservation Biology 2(4):348-358. https://doi.org/10.1111/j.1523-1739.1988.tb00199.x
Arnold, T. W., and R. M. Zink. 2011. Collision mortality has no discernible effect on population trends of North American birds. PLoS One 6(9):e24708. https://doi.org/10.1371/journal.pone.0024708
Betts, M. G., Z. Yang, A. S. Hadley, A. C. Smith, J. S. Rousseau, J. M. Northrup, J. J. Nocera, N. Gorelick, and B. D. Gerber. 2022. Forest degradation drives widespread avian habitat and population declines. Nature Ecology & Evolution 6(6):709-719. https://doi.org/10.1038/s41559-022-01737-8
Brady, R. S., R. A. Staffen, N. M. Anich, and C. N. Lapin. 2021. Status and trend of the Connecticut Warbler in Wisconsin: steep decline demands urgent conservation action. Passenger Pigeon.
Donovan, T. M., R. H. Lamberson, A. Kimber, F. R. Thompson, III, and J. Faaborg. 1995. Modeling the effects of habitat fragmentation on source and sink demography of Neotropical migrant birds. Conservation Biology 9(6):1396-1407. https://doi.org/10.1046/j.1523-1739.1995.09061396.x
Elder, D. H. 1991. Breeding habitat of the Connecticut Warbler in the Rainy River District. Ontario Birds 9:84-86.
Erskine, A. J. 1977. Birds in boreal Canada: communities, densities and adaptations. Canadian Wildlife Service Report Series No. 41.
Fink, D., T. Auer, A. Johnston, V. Ruiz-Gutierrez, W. M. Hochachka, and S. Kelling. 2020. Modeling avian full annual cycle distribution and population trends with citizen science data. Ecological Applications 30(3):e02056. https://doi.org/10.1002/eap.2056
Glennon, M. J., S. F. Langdon, M. A. Rubenstein, and M. S. Cross. 2019. Temporal changes in avian community composition in lowland conifer habitats at the southern edge of the boreal zone in the Adirondack Park, NY. PloS One 14(8):e0220927. https://doi.org/10.1371/journal.pone.0220927
Grinde, A. R., R. Siebers, S. Kolbe, and J. D. Bednar. 2022. Minnesota National Forest Breeding Bird Monitoring Program annual report 1995-2022. University of Minnesota, Duluth, Minnesota, USA.
Hallworth, M. T., E. Bayne, E. McKinnon, O. Love, J. A. Tremblay, B. Drolet, J. Ibarzabal, S. Van Wilgenburg, and P. P. Marra. 2021. Habitat loss on the breeding grounds is a major contributor to population declines in a long-distance migratory songbird. Proceedings of the Royal Society B 288(1949):20203164. https://doi.org/10.1098/rspb.2020.3164
Handler, S., M. J. Duveneck, L. Iverson, E. Peters, R. M. Scheller, K. R. Wythers, L. Brandt, P. Butler, M. Janowiak, P. D. Shannon, et al. 2014. Minnesota forest ecosystem vulnerability assessment and synthesis: a report from the Northwoods Climate Change Response Framework Project. U.S. Forest Service. https://doi.org/10.2737/NRS-GTR-133
Hanowski, J. M., and G. J. Niemi. 1983. Breeding bird censuses: Red Lake Peatland, northern Minnesota. American Birds 37:100-101.
Hanski, I. K., T. J. Fenske, and G. J. Niemi. 1996. Lack of edge effect in nesting success of breeding birds in managed forest landscapes. Auk 113(3):578-585. https://doi.org/10.2307/4088978
Harrison, C. J. O. 1978. A field guide to the nests, eggs and nestlings of North American birds. Collins, Toronto, Ontario, Canada.
Hitch, A. T., and P. L. Leberg. 2007. Breeding distributions of North American bird species moving north as a result of climate change. Conservation Biology 21(2):534-539. https://doi.org/10.1111/j.1523-1739.2006.00609.x
International Union for Conservation of Nature (IUCN). 2019. The IUCN Red List of threatened species. Version 2019-1. http://www.iucnredlist.org
James, F. C., and H. H. Shugart, Jr. 1970. A quantitative method of habitat description. Audubon Field Notes 24(6):727-736.
Janssen, R. B. 1987. Birds in Minnesota. University of Minnesota Press, Minneapolis, Minnesota, USA.
Kirk, D. A., A. W. Diamond, K. A. Hobson, and A. R. Smith. 1996. Breeding bird communities of the western and northern Canadian boreal forest: relationship to forest type. Canadian Journal of Zoology 74(9):1749-1770. https://doi.org/10.1139/z96-193
Kudell-Ekstrum, J. 2002. Conservation assessment for Connecticut Warbler (Oporonis agilis). U.S. Forest Service.
Lachance, D., C. Lavoie, and A. Desrochers. 2005. The impact of peatland afforestation on plant and bird diversity in southeastern Québec. Écoscience 12(2):161-171. https://doi.org/10.2980/i1195-6860-12-2-161.1
Lapin, C. N., M. A. Etterson, and G. J. Niemi. 2013. Occurrence of the Connecticut Warbler increases with size of patches of coniferous forest: La Ocurrencia de Oporornis agilis Aumenta con el Tamaño de los Parches de Bosques de Coníferas. Condor 115(1):168-177. https://doi.org/10.1525/cond.2013.110202
Lawton, J. H. 1993. Range, population abundance and conservation. Trends in Ecology & Evolution 8(11):409-413. https://doi.org/10.1016/0169-5347(93)90043-O
Lenoir, J., and J. C. Svenning. 2015. Climate-related range shifts-a global multidimensional synthesis and new research directions. Ecography 38(1):15-28. https://doi.org/10.1111/ecog.00967
Longcore, T., C. Rich, P. Mineau, B. MacDonald, D. G Bert, L. M. Sullivan, E. Mutrie, S. A. Gauthreaux, Jr, M. L. Avery, R. L. Crawford, and A. M. Manville, II. 2013. Avian mortality at communication towers in the United States and Canada: which species, how many, and where? Biological Conservation 158:410-419. https://doi.org/10.1016/j.biocon.2012.09.019
Loss, S. R., T. Will, S. S. Loss, and P. P. Marra. 2014. Bird–building collisions in the United States: estimates of annual mortality and species vulnerability. Condor 116(1):8-23. https://doi.org/10.1650/CONDOR-13-090.1
Martin, T. E., and G. R. Geupel. 1993. Nest-monitoring plots: methods for locating nests and monitoring success (Métodos para localizar nidos y monitorear el éxito de estos). Journal of Field Ornithology 507-519.
Martin, T. E., C. R. Paine, C. J. Conway, W. M. Hochachka, P. Allen, and W. Jenkins. 1997. BBird field protocol. Montana Cooperative Wildlife Research Unit, University of Montana, Missoula, Montana, USA.
Matricardi, E. A. T., D. L. Skole, O. B. Costa, M. A. Pedlowski, J. H. Samek, and E. D. Miguel. 2020. Long-term forest degradation surpasses deforestation in the Brazilian Amazon. Science 369(6509):1378-1382. https://doi.org/10.1126/science.abb3021
Mattsson, B. J., and G. J. Niemi. 2006. Factors influencing predation on Ovenbird (Seiurus aurocapilla) nests in northern hardwoods: interactions across spatial scales. Auk 123(1):82-96. https://doi.org/10.1093/auk/123.1.82
Minnesota Department of Natural Resources (MN DNR). 2019. Minnesota’s forest resources. Division of Forestry. https://files.dnr.state.mn.us/forestry/um/forest-resources-report-2019.pdf
Nevers, M. B., J. M. Hanowski, and G. J. Niemi. 1981. Breeding bird censuses in the Red Lake Peatland, northern Minnesota. American Birds 35:99-100.
Niemi, G. J., R. W. Howe, B. R. Sturtevant, L. R. Parker, A. R. Grinde, N. P. Danz, M. D. Nelson, E. J. Zlonis, N. G. Walton, E. E. G. Giese, and S. M. Lietz. 2016. Analysis of long-term forest bird monitoring data from national forests of the western Great Lakes region. U.S. Forest Service General Technical Report NRS-159.
Niemi, G. J., and J. M. Hanowski. 1984. Effects of a transmission line on bird populations in the Red Lake Peatland, northern Minnesota. Auk 101(3):487-498. https://doi.org/10.1093/auk/101.3.487
Padgham, M., and M. D. Sumner. 2021. geodist: fast, dependency-free geodesic distance calculations. R package. Version 0.0.7. https://github.com/hypertidy/geodist
Partners in Flight. 2021. Avian conservation assessment database. Version 2021.
Payne, R. B. 1983. A distributional checklist of the birds of Michigan. University of Michigan, Ann Arbor, Michigan, USA.
Paynter, R. A., Jr. 1995. Nearctic passerine migrants in South America. Publications of the Nuttall Ornithological Club 25, Cambridge, Massachusetts, USA.
Peck, G. K., and R. D. James. 1987. Breeding birds of Ontario: nidiology and distribution. Volume 2: Passerines. Miscellaneous Publications of the Royal Ontario Museum, Toronto, Ontario, Canada.
Pfannmuller, L., G. Niemi, J. Green, and K. Rewinkel. 2023. Breeding birds of Minnesota: their history, ecology, and conservation. University of Minnesota Press, Minneapolis, Minnesota USA, in press.
Pinheiro, J., D. Bates, S. DebRoy, D. Sarkar, R. C. Team. 2021. nlme: linear and nonlinear mixed effects models.
Pitocchelli, J., J. L. Jones, D. Jones, and J. Bouchie. 2020. Connecticut Warbler (Oporornis agilis). Version 1.0. In A. F. Poole, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.conwar.01
R Core Team. 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rappole, J. H. 1995. The ecology migrant birds. A Neotropical perspective. Smithsonian Institution Press, Washington, D.C., USA.
Rappole, J. H., and A. R. Tipton. 1991. New harness design for attachment of radio transmitters to small passerines (Nuevo Diseño de Arnés para Atar Transmisores a Passeriformes Pequeños). Journal of Field Ornithology 335-337.
Ridgely, R. S., G. Tudor, and W. L. Brown. 1989. The birds of South America. Volume 1: the oscine passerines. University of Texas Press.
Robinson, S. K., and D. S. Wilcove. 1994. Forest fragmentation in the temperate zone and its effects on migratory songbirds. Bird Conservation International 4(2-3):233-249. https://doi.org/10.1017/S0959270900002793
Rodewald, A. D. 2004. Nest-searching cues and studies of nest-site selection and nesting success. Journal of Field Ornithology 75(1):31-39. https://doi.org/10.1648/0273-8570-75.1.31
Rosenberg, K. V., A. M. Dokter, P. J. Blancher, J. R. Sauer, A. C. Smith, P. A. Smith, J. C. Stanton, A. Panjabi, L. Helft, M. Parr, and P. P. Marra. 2019. Decline of the North American avifauna. Science 366(6461):120-124. https://doi.org/10.1126/science.aaw1313
Sauer, J. R., W. A. Link, and J. E. Hines. 2020. The North American Breeding Bird Survey, analysis results 1966–2021: U.S. Geological Survey. https://doi.org/10.5066/P9SC7T11
Sekercioglu, Ç.H., G. C. Daily, and P. R. Ehrlich. 2004. Ecosystem consequences of bird declines. Proceedings of the National Academy of Sciences 101(52):18042-18047. https://doi.org/10.1073/pnas.0408049101
Sodhi, N. S., and C. A. Paszkowski. 1995. Research on Wood Warblers in Canada (Estudios Sobre Parúlidos en Canadá). Journal of Field Ornithology 66:260-266.
Stralberg, D., E. M. Bayne, S. G. Cumming, P. Sólymos, S. J. Song, and F. K. Schmiegelow. 2015. Conservation of future boreal forest bird communities considering lags in vegetation response to climate change: a modified refugia approach. Diversity and Distributions 21(9):1112-1128. https://doi.org/10.1111/ddi.12356
Stralberg, D., D. Berteaux, C. R. Drever, M. Drever, I. Naujokaitis-Lewis, F. Schmiegelow, and J. Tremblay. 2019. Conservation planning for boreal birds in a changing climate: a framework for action. Avian Conservation and Ecology 14(1):13. https://doi.org/10.5751/ACE-01363-140113
Streby, H. M., and D. E. Andersen. 2013. Movements, cover-type selection, and survival of fledgling Ovenbirds in managed deciduous and mixed coniferous-deciduous forests. Forest Ecology and Management 287:9-16. https://doi.org/10.1016/j.foreco.2012.08.046
Svenson, S., and K. Williamson. 1969. Recommendations for an international standard for a mapping method in bird census work. Bird Study 16(4):249-255.
Wilsey, C., B. Bateman, L. Taylor, J. X. Wu, G. LeBaron, R. Shepherd, C. Koseff, S. Friedman, and R. Stone. 2019. Survival by degrees: 389 bird species on the brink. National Audubon Society, New York, New York, USA.
Zlonis, E., H. Panci, J. Bednar, M. Hamady, and G. Niemi. 2017. Habitats and landscapes associated with bird species in a lowland conifer-dominated ecosystem. Avian Conservation and Ecology 12(1):7. https://doi.org/10.5751/ACE-00954-120107
Zuckerberg, B., A. M. Woods, and W. F. Porter. 2009. Poleward shifts in breeding bird distributions in New York State. Global Change Biology 15(8):1866-1883.
Fig. 1
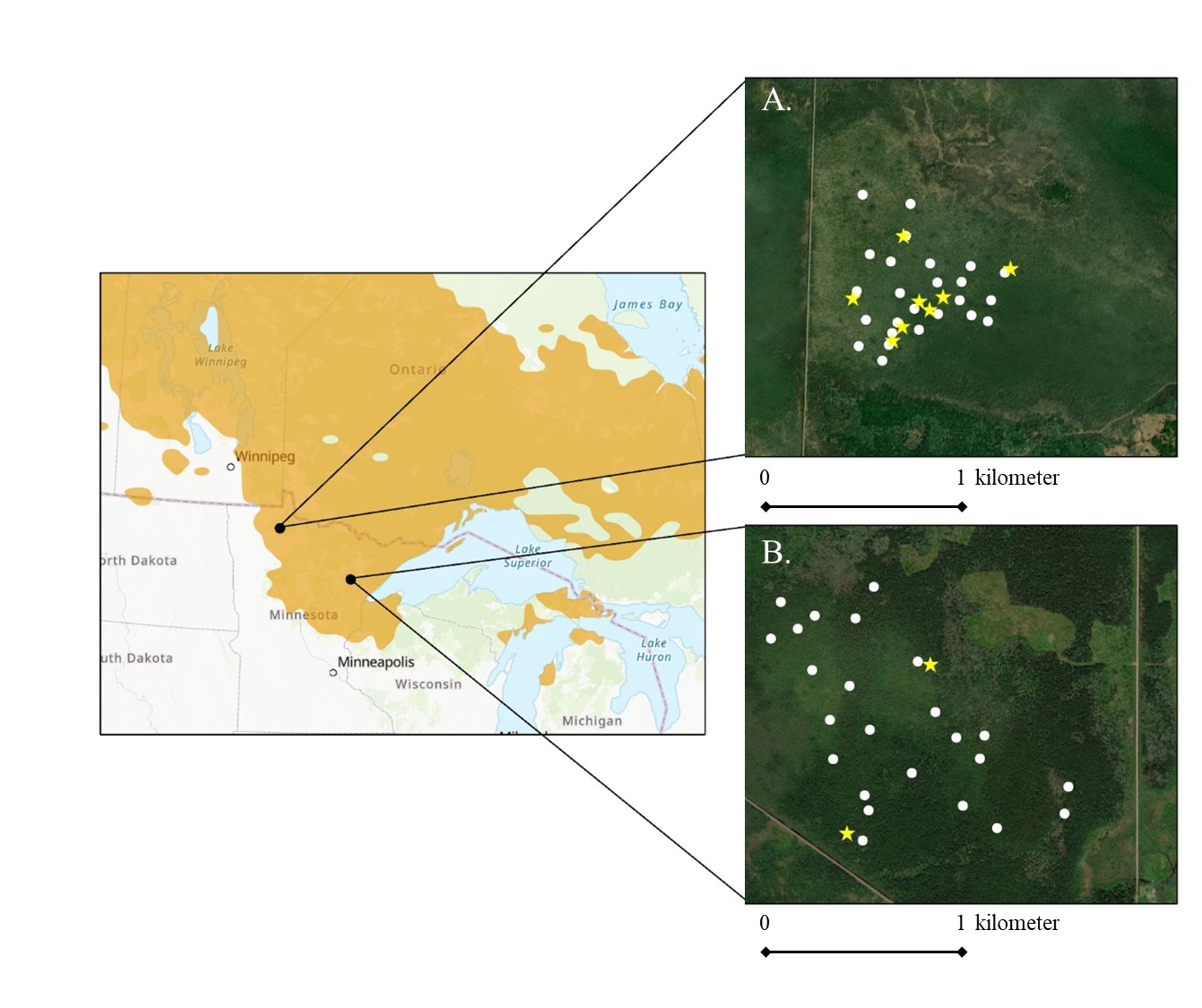
Fig. 1. Locations of the project study areas. Connecticut Warbler nest searching efforts focused on the Red Lake Wildlife Management Area (A) in 2020 and the Sax-Zim Bog (B) in 2019. White circles represent singing males; yellow stars represent nests found during systematic and behavioral nest searches. The Red Lake Wildlife Management study area is located in the Agassiz Lowlands ecological subsection, and the Sax-Zim Bog study area is located in the Tamarack Lowlands ecological subsection, both of which are shown by black dots on the overview map. Connecticut Warbler breeding range is shown in orange (from Fink et al. 2020).
Fig. 2

Fig. 2. Representative photographs of Connecticut Warbler nests during the (A) egg laying, (B) nestling, and (C) pre-fledge periods.
Fig. 3
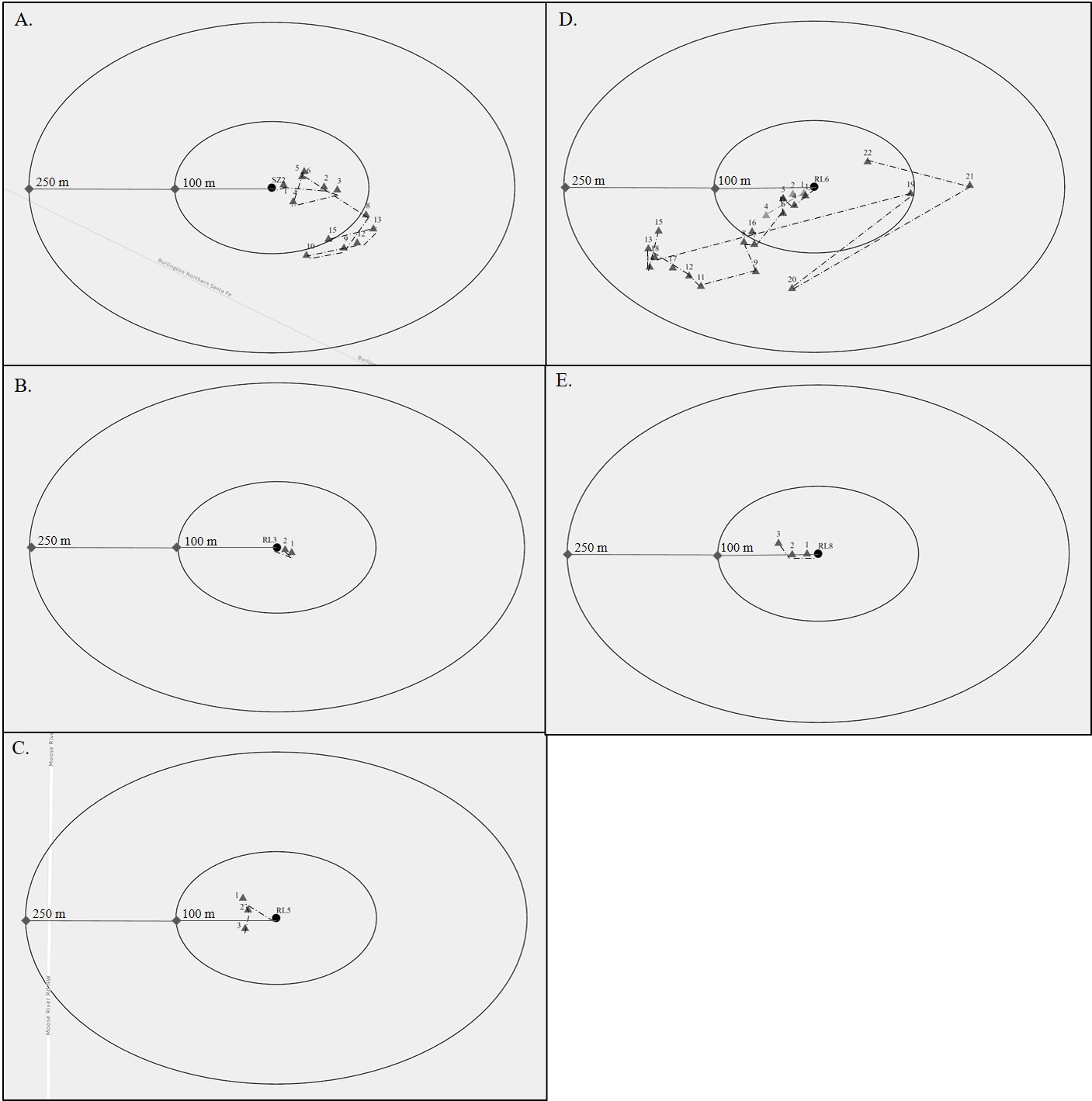
Fig. 3. Telemetry locations of Connecticut Warbler fledglings from five nests during the post-fledging period in 2019 and 2020 in northern Minnesota, USA. Nest locations are represented by black dots; daily fledgling locations are represented by triangles and are labeled with the number of days since leaving the nest; and 100-m and 250-m buffers are drawn around each nest location.
Fig. 4
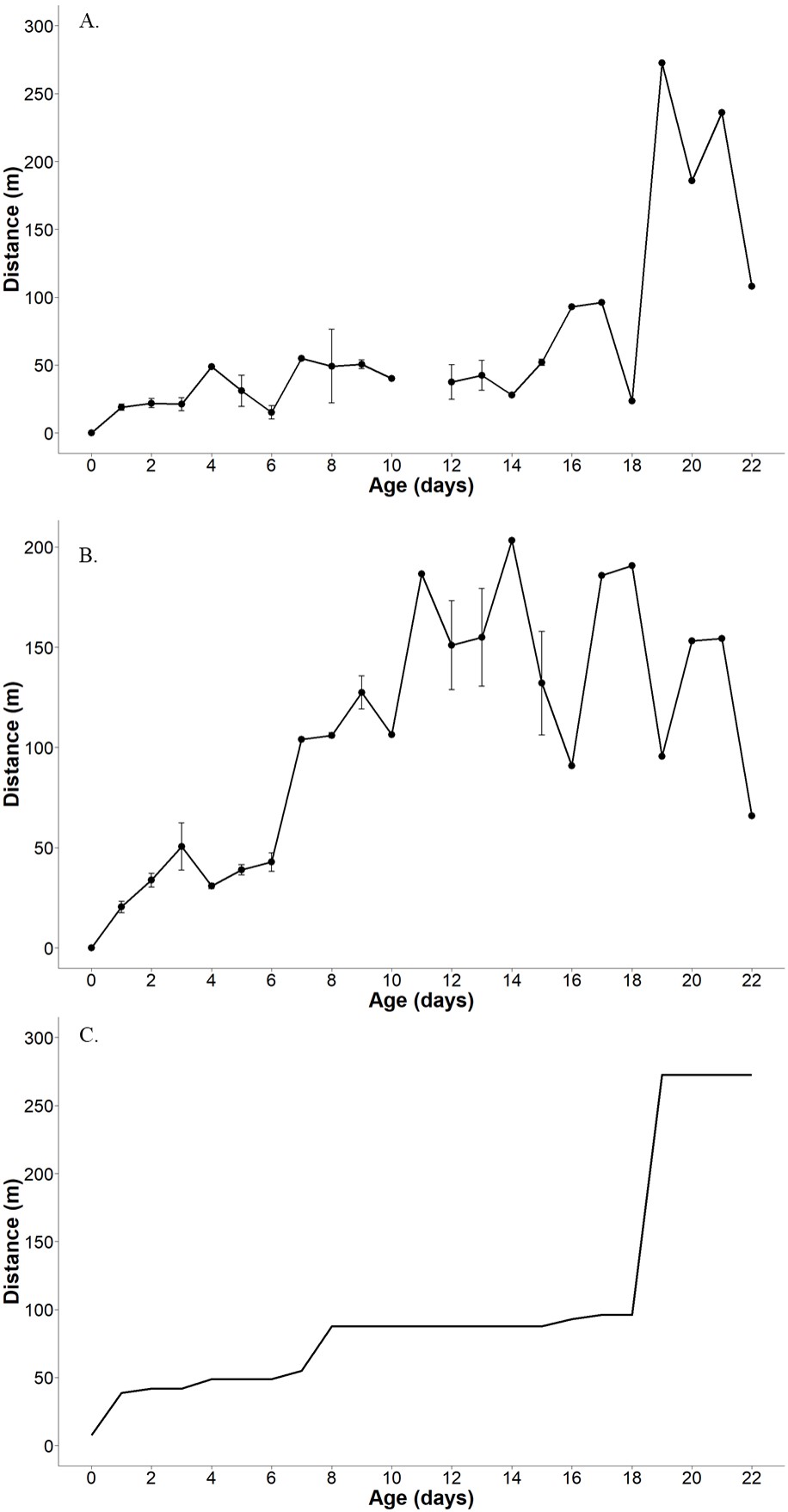
Fig. 4. Distances moved by Connecticut Warbler fledglings from five nests during the post-fledging period in 2019 and 2020 in northern Minnesota, USA (age is represented as the number of days after leaving the nest). (A) Mean daily distance moved by fledgling Connecticut Warblers from the previous day’s observed location, (B) mean daily distance from the nest, and (C) maximum observed movement for each post-fledge day. Error bars represent +/-1 standard error.
Table 1
Table 1. Nests found and monitored in the Sax-Zim and Red Lake study areas of northern Minnesota, USA in 2019 and 2020.
| Nest ID | Study area | Year | Date nest was found | Stage of nest when found | No. eggs | Est. hatch date | No. nestlings | Est. fledge date | No. fledglings | No. indiv. banded | No. transmitters deployed |
| SZ1 | Sax-Zim | 2019 | 16-Jun | Incubation | 4 | 30-Jun | 3 | 6-Jul | 3 | 2 | 2 |
| SZ2 | Sax-Zim | 2019 | 1-Jul | Nestling | – | 30-Jun | 4 | 6-Jul | 4 | 3 | 2 |
| RL1 | Red Lake | 2020 | 8-Jun | Laying | 1 | Abandoned | 0 | – | – | – | – |
| RL2 | Red Lake | 2020 | 11-Jun | Incubation | 5 | Abandoned | 0 | – | – | – | – |
| RL3 | Red Lake | 2020 | 22-Jun | Nestling | – | 14-Jun | 4 | 22-Jun | 4 | 1 | 1 |
| RL4 | Red Lake | 2020 | 25-Jun | Nestling | – | 18-Jun | 5 | 27-Jun | 5 | 0 | 0 |
| RL5 | Red Lake | 2020 | 25-Jun | Incubation | 4 | 3-Jul | 3 | 11-Jul | 3 | 3 | 3 |
| RL6 | Red Lake | 2020 | 27-Jun | Nestling | – | 20-Jun | 5 | 28-Jun | 5 | 5 | 2 |
| RL7 | Red Lake | 2020 | 2-Jul | Incubation | 4 | 11-Jul | 4 | 18-Jul | 3 | 0 | 0 |
| RL8 | Red Lake | 2020 | 2-Jul | Nestling | – | 2-Jul | 5 | 9-Jul | 5 | 5 | 3 |
| RL9 | Red Lake | 2020 | 12-Jul | Nestling | – | 4-Jul | 5 | 12-Jul | 5 | 0 | 0 |
Table 2
Table 2. Mean and standard errors for vegetation characteristics on 11.3-m (0.04-ha) plots used by fledglings, and on paired random points in the Sax-Zim and Red Lake study areas of northern Minnesota, USA in 2019 and 2020. t tests were used to determine differences between paired used and random plots across both years. Parentheses indicate conversion to “"per hectare” values.
| Vegetation variable | Bird | Random | Difference between used plot random plots | ||||||
| Mean | SE | Mean | SE | Mean | SE | t | P | ||
| Total tree count | 73 (1836) | 3 (85) | 70 (1750) | 3 (85) | 2.8 | 2.7 | 1.0 | 0.32 | |
| Small tree count | 72 (1808) | 4 (90) | 70 (1750) | 4 (90) | 2.4 | 2.8 | 0.9 | 0.39 | |
| Total stem count | 288 (7193) | 29 (735) | 241 (6000) | 23 (580) | 46.9 | 20.2 | 2.3 | 0.03 | |
| Canopy cover (%) | 39.3 | 4.6 | 40.7 | 4.2 | -1.4 | 2.8 | 0.5 | 0.62 | |
| Sphagnum depth (mm) | 7.6 | 0.5 | 8.0 | 0.6 | -0.4 | 0.7 | 0.5 | 0.61 | |
| Grass ground cover (%) | 16.1 | 2.3 | 16.7 | 2.8 | -0.5 | 5.7 | 0.1 | 0.91 | |
| Shrub ground cover (%) | 34.3 | 4.1 | 30.7 | 4.1 | 3.5 | 2.4 | 1.5 | 0.15 | |
| Sphagnum ground cover (%) | 34.2 | 3.5 | 36.1 | 3.5 | -2.0 | 2.7 | 0.7 | 0.46 | |