







The following is the established format for referencing this article:
Gaylord, M. A., A. Duarte, B. C. McComb, and J. Ratliff. 2023. Passive acoustic recorders increase White-headed Woodpecker detectability in the Blue Mountains. Journal of Field Ornithology 94(4):1.ABSTRACT
White-headed Woodpeckers (Dryobates albolarvatus) are strongly associated with late-successional dry forest types. Callback surveys along transects are typically used to understand their status and trends in response to forest management. However, this survey method has proven to be logistically challenging because of the number of spatial and temporal replicate surveys needed to accurately interpret surveys that yield no detections. Passive acoustic recording units (ARUs) effectively detect certain avian species and may offer a more efficient and effective survey method, but few studies have focused on detecting White-headed Woodpeckers. Our objectives were to: (1) compare detection probabilities of White-headed Woodpeckers between callback surveys and ARUs, and (2) estimate the number of surveys needed to infer White-headed Woodpeckers’ absence under different levels of certainty. We surveyed for White-headed Woodpeckers from 5 May to 15 July 2021 by conducting callback surveys along six transects, with 10 survey stations along each, and deploying ARUs at 25 survey stations across three watersheds in the Wallowa-Whitman National Forest, Oregon, USA. We developed a classifier for White-headed Woodpeckers to detect their two-, three-, and four-note calls in our ARU data. Using single-season occupancy models and Akaike Information Criterion corrected for small sample sizes, the best fit model indicated that the odds of detecting White-headed Woodpeckers were 1.28 times higher approximately every 10 days into the breeding season and 4.41 times lower when using callback surveys compared to using ARUs. The cumulative detection probability for ARUs ranged from 0.95 to 0.99 after being deployed for 5 and 8 days, respectively. The cumulative detection probability was only 0.15–0.38 after 1 and 3 replicate callback survey(s) at a survey station, respectively. Our study demonstrates that managers can gather more accurate data related to the presence/absence of White-headed Woodpeckers to inform forest management decisions when using a passive acoustic monitoring design.
RESUMEN
El pájaro carpintero de cabeza blanca (Dryobates albolarvatus) está fuertemente asociado con bosques secos de sucesión tardía. Los estudios de “playback” a lo largo de transectos se utilizan normalmente para comprender su estado y tendencias en respuesta al manejo forestal. Sin embargo, este método de encuesta ha demostrado ser un desafío logístico debido a la cantidad de replicas espaciales y temporales necesarias para interpretar con precisión las encuestas en las que no ocurrieron detecciones. Las unidades de grabación acústica pasiva (ARU) detectan eficazmente ciertas especies de aves y pueden ofrecer un método de estudio más eficiente y eficaz, pero pocos estudios se han centrado en la detección del pájaro carpintero de cabeza blanca. Nuestros objetivos fueron: (1) comparar las probabilidades de detección de pájaros carpinteros de cabeza blanca entre búsquedas con llamados a través de cantos y ARU, y (2) estimar el número de encuestas necesarias para inferir la ausencia de pájaros carpinteros de cabeza blanca bajo diferentes niveles de certeza. Realizamos estudios de pájaro carpintero de cabeza blanca del 5 de mayo al 15 de julio de 2021 mediante la búsqueda con llamados a través de cantos a lo largo de seis transectos, con 10 estaciones de estudio a lo largo de cada uno, y el despliegue de ARU en 25 estaciones de estudio en tres cuencas hidrográficas en el Bosque Nacional Wallowa-Whitman, Oregón, EE. UU. Desarrollamos un clasificador para el pájaro carpintero de cabeza blanca para detectar sus llamadas de dos, tres y cuatro notas en nuestros datos de ARU. Utilizando modelos de ocupación de una sola temporada y el Criterio de Información de Akaike corregido para tamaños de muestra pequeños, el modelo de mejor ajuste indicó que las probabilidades de detectar el pájaro carpintero de cabeza blanca eran 1,28 veces mayores aproximadamente cada 10 días en la temporada de reproducción y 4,41 veces menores cuando se usaban búsqueda con llamados a través de cantos en comparación con el uso de ARU. La probabilidad de detección acumulada de las ARU osciló entre 0,95 y 0,99 después de haber sido desplegadas durante 5 y 8 días, respectivamente. La probabilidad de detección acumulada fue solo de 0,15 a 0,38 después de 1 y 3 réplicas de búsqueda con llamados a través de cantos repetidas en una estación de encuesta, respectivamente. Nuestro estudio demuestra que los administradores pueden recopilar datos más precisos relacionados con la presencia/ausencia de pájaros carpinteros de cabeza blanca para informar las decisiones de manejo forestal cuando utilizan un diseño de monitoreo acústico pasivo.
INTRODUCTION
Dry forest types in the Blue Mountains of Oregon, USA, include warm, dry sites at low-to-middle elevations (900–1500 m) where ponderosa pine (Pinus ponderosa) was once the major forest component (Franklin and Dyrness 1973). From the 1930s to 2000s, late-successional dry forests in the Blue Mountains have declined by 90% (Buchanan et al. 2003, Powell 2011, Altman and Bresson 2017). Fire suppression, livestock grazing, and timber harvest, among other factors, have resulted in increasingly dense understories of small trees (< 30 cm diameter-at-breast height [dbh]; Harrod et al. 1998, Youngblood 2000). There has also been a significant reduction of medium (43–64 cm dbh) and large (> 64 cm dbh) trees (Harrod et al. 1998). In addition, cutting snags for fuelwood (Bate et al. 2007) and timber management practices (Wisdom and Bate 2008) have reduced the number of potential nesting sites for primary cavity nesters (Bull 2002, Wisdom and Bate 2008).
White-headed Woodpeckers (Dryobates albolarvatus) are strongly associated with late-successional dry forest types east of the Cascade Mountain Range (Dixon 1995, Latif et al. 2015, Latif et al. 2017, Latif et al. 2020). Their abundance and distribution have been proposed as a metric that can be used to assess progress toward restoring this forest type (Lambeck 1997, Lambeck 1999). White-headed Woodpeckers were identified by United States Department of Agriculture (USDA) Forest Service Region 6 (Forest Service Manual [FSM] 2670.5) as a “sensitive species”—a species that is experiencing or is threatened with a small, declining, and/or at-risk population; habitat loss; and/or is a management concern (FSM 2670.5). They have been considered a sensitive species in Oregon since 1989 and an Oregon “strategy species” for the Blue Mountain ecoregion because of degraded or lost habitat (Oregon Department of Fish and Wildlife 2021). White-headed Woodpeckers are considered “imperiled” in Idaho (Idaho Department of Fish and Game 2021) and a state candidate for listing in Washington (Washington Department of Fish and Wildlife 2021). Partners in Flight have also listed White-headed Woodpecker as a “species of concern” (Partners in Flight 2019). As such, effective White-headed Woodpecker monitoring strategies are needed to inform management decisions when restoring late-successional dry forest structure and function.
Current monitoring of White-headed Woodpeckers across dry forest landscapes relies on repeated callback surveys at survey stations along transects. However, this survey method has proven to be logistically challenging because of the number of replicate surveys needed to infer absence, the timing of the surveys (i.e., dawn callback surveys) in relation to the remote forests this species inhabits, and the relatively short season in which these surveys need to take place (i.e., May–June). Recent work indicates that several avian species can be detected by using passive acoustic recording units (ARUs; Swiston and Mennill 2009, Van Wilgenburg et al. 2017, Ruff et al. 2021), which may overcome several of the limitations in the current White-headed Woodpecker monitoring strategy. As such, our objectives were to: (1) compare detection probabilities of White-headed Woodpeckers between callback surveys at survey stations along transects and passive recordings at survey stations using ARUs, and (2) estimate the number of surveys needed to infer White-headed Woodpeckers’ absence under different levels of certainty. We hypothesized that detection probability per day for the two survey types would be equal based on previous comparisons between point-count surveys and ARU recordings (Pacifici et al. 2008, Priyadarshani et al. 2018) and the fact that both survey types largely rely on auditory detections.
METHODS
Study area
Because our objectives for this study focused on understanding White-headed Woodpecker detection probabilities using two different survey methods, rather than understanding environmental factors that are related to the occurrence of the species, we conducted our surveys within three focal watersheds (Big Creek, Ruckles Creek-Powder River, Eagle Creek) on the Wallowa-Whitman National Forest (Fig. 1) where White-headed Woodpeckers had previously been observed (J. Ratliff, personal observation). Although White-headed Woodpeckers were previously observed within the watersheds, occupancy at specific survey stations was unknown during our field season. The study area was approximately 23,987 ha, dominated by ponderosa pine and mixed conifer species. Although the northern Blue Mountains is a relatively dry landscape, the maritime air flowing through the Columbia River Gorge increases the amount of precipitation (especially in the form of snow [average snow depth 63.5–165.1 cm above 1370 m in elevation]) and reduces temperature variations compared to that which is experienced in the southern portion of the Blue Mountains (Western Regional Climate Center 2022). Sunrise and sunset times were approximately 0507–0540 and 2000–2033 hours, respectively. Other woodpecker species that co-occur with White-headed Woodpecker in the area include American Three-toed Woodpecker (Picoides dorsalis), Black-backed Woodpecker (P. arcticus), Downy Woodpecker (Dryobates pubescens), Hairy Woodpecker (D. villosus), Lewis’s Woodpecker (Melanerpes lewis), Pileated Woodpecker (Dryocopus pileatus), Northern Flicker (Colaptes auratus), Red-naped Sapsucker (Sphyrapicus nuchalis), and Williamson’s Sapsucker (S. thyroideus).
Callback surveys
We surveyed along six transects with 10 survey stations per transect and deployed 25 ARUs from 5 May to 15 July 2021. Transect placement followed the protocols typically used to monitor White-headed Woodpeckers in national forests in the region (Appendix B of Mellen-McLean et al. 2015). Transects were randomly distributed across the study area, approximately 7 km apart and accessible by road. This resulted in one transect in Big Creek watershed, three in Ruckles Creek-Powder River watershed, and two in Eagle Creek watershed. Furthermore, we ensured survey stations along transects were ≥ 300 m apart to minimize the chances of detecting the same bird at two neighboring survey stations on the same day.
We conducted callback surveys along transects following Appendix B of the Mellen-McLean et al. (2015) protocol. In general, surveyors played a White-headed Woodpecker call recording three times with a 30-s pause between recordings followed by a two-minute listening period. Typically, survey stations are surveyed two times in a season; however, we surveyed along each transect three times to improve our ability to estimate detection probabilities. We conducted these replicate surveys at least two weeks apart and always surveyed in the opposite direction along the transect from the previous survey. We spent 2.5–4.5 h surveying each transect per survey morning. We broadcast calls using a FoxPro NX4 digital game call and recorded observations using ArcGIS Survey123 (Environmental Systems Research Institute 2022). Although we recorded each White-headed Woodpecker observation along transects, we only used detections at the 60 survey stations for our analyses we present herein.
Passive acoustic surveys
To select ARU survey stations, we overlayed a 500-ha hexagon grid, where each hexagon contained six 83-ha triangles. We randomly selected 25 triangles to deploy our ARUs. We deployed the ARUs as close to the center of each randomly selected triangle as possible, which ensured that ARU survey stations were ≥ 907 m apart. Thus, it was unlikely that the same bird would be detected at two different ARUs given that the minimum spacing between ARUs approached the maximum estimated home range size (18.9 ha, SD = 10.1, range 3.1–35.7 ha) for White-headed Woodpeckers in this forest type (Kehoe 2017). Prior to initial deployment, four ARU survey stations were omitted because of inaccessibility, private land, or environmental noise issues; therefore, four replacement locations were randomly selected. The final locations of the 60 transect survey stations and 25 ARU survey stations can be seen in Fig. 1.
We used the Song Meter 4 Mini Bat with the acoustic microphone stub attachment (used to record avian species) that is manufactured by Wildlife Acoustics (Wildlife Acoustics, Maynard, Massachusetts, USA). The sample rate was set to 32,000 Hz and 16-bit pulse code modulation .wav files were recorded. We attached ARUs to trees that were < 30 cm dbh at approximately 1.5 m above ground with the microphone facing east. We removed any branches that were within 30 cm of the ARU to reduce environmental noise. We ensured each unit was deployed at least 60 m from roads to reduce the chance that they would be stolen and 100 m from large creeks to further reduce environmental noise. We programed the ARUs to record continuously from 0000 to 1100 hours Pacific Daylight Time (PDT) and for 10 min on the top of every hour for all other time periods (1100–2310 hours) for a total of 13.2 h recorded daily. This recording schedule allowed us to collect audio data when White-headed Woodpeckers are most likely to be active (30 min after sunrise to noon; Mellen-McLean et al. 2015), to collect intermittent recordings throughout the day, and passively record other species of interest. We checked and replaced batteries and memory cards as needed to ensure continuous recordings (approximately every three weeks).
Audio analyses
We analyzed audio data collected by the ARUs using Kaleidoscope Pro 5 software (Wildlife Acoustics 2020). Kaleidoscope uses classifiers to custom filter audio recordings based on individual or clusters of vocalizations (i.e., an acoustic pattern). We randomly selected ≥ 12 h of recordings from each ARU that recorded between 5 May and 15 July. We used this subset of audio recordings (457.5 h of 20,423.7 h) to develop a new classifier that could identify White-headed Woodpecker two-, three-, and four-note calls. We focused on these calls because they are easily distinguishable from co-occurring woodpecker species in the area and these calls are used to indicate species presence during traditional callback surveys. When processing these data, we set the program to scan and cluster audio recordings using the following specifications: Fast Fourier Transform (FFT) window set to 2.67 ms. (128 @ 0–24 kHz 256 @ 25–48 kHz 512 @ 49–96 kHz), number of clusters ≤ 100, frequency range of 3–4.5 kHz, detection calls of 0.1–2.0 s in length, and a maximum of 0.1 s inter-syllable gap. After the initial clusters were created (n = 100), we manually labeled a subset of the audio clips (0.1–2.0 s in length) to train the classifier to recognize White-headed Woodpecker vocalizations (138,739 of 342,357). In total, 588 audio clips out of the 138,739 audio clips were identified as White-headed Woodpeckers and used to train the classifier.
Using the classifier, we processed all audio files for each ARU. We then manually reviewed the output audio clips from the classifier to ensure no false positive White-headed Woodpecker detections (i.e., misclassified vocalizations of a White-headed Woodpecker) occurred in our data. If false positives were identified, they were manually removed until only true, verified positive White-headed Woodpecker detections remained for our use in subsequent analyses.
Although we manually verified all White-headed Woodpecker call detections for our study, we also wanted to formally evaluate how well the segmentation algorithm identified White-headed Woodpecker calls. We did this by calculating recall and precision, which quantify how likely a focal species detection was correctly identified (true positive) versus misidentified (false negative or false positive; Priyadarshani et al. 2018). In this context, a false negative is a White-headed Woodpecker call that was not identified by the classifier, and a false positive is a sound recording that is incorrectly identified as a White-headed Woodpecker. Recall is the proportion of White-headed Woodpecker calls in the audio data the classifier identified and is estimated as the ratio between the true positives identified by the classifier and the total number of White-headed Woodpecker call recordings in the data (Priyadarshani et al. 2018). Given the time required to review all audio recordings, a subset (22.5 h) of recordings with White-headed Woodpecker detections were randomly selected and reviewed to calculate recall. Precision is a measure of how well the classifier identifies White-headed Woodpecker calls and is estimated as the proportion of White-headed Woodpecker detections (both true and false positives) that were true positive detections (Priyadarshani et al. 2018). The precision was calculated by reviewing all ARU audio clips identified as White-headed Woodpecker by the classifier.
Occupancy modeling
Because it was not guaranteed our survey stations were occupied by White-headed Woodpeckers, we fit single-season occupancy models to our data to model the ecological and observation process separately (MacKenzie et al. 2002, Tyre et al. 2003). In these models, occupancy probability (ψ) is the probability that a survey station had at least one White-headed Woodpecker present, and detection probability (p) is the probability that the species was detected during a survey occasion, given that the survey station was occupied by White-headed Woodpecker. Occupancy models require a few assumptions: the occupancy state (species presence or absence) does not change at a survey station across replicate surveys, detection probabilities are independent across survey stations and replicate surveys (i.e., detecting the species does not influence the odds of detecting it at neighboring survey stations or in subsequent survey occasions), variation in occupancy and detection probability can be accounted for using covariates, and false positive detections do not occur in the detection/non-detection data. Because White-headed Woodpeckers can travel greater distances outside the nesting period (Lorenz et al. 2015), we omitted any data collected outside of the nesting period during analyses to minimize turnover in the occupancy state at a survey station. Although the exact nesting period can vary from year to year (and region to region) depending on local environmental conditions, we considered the nesting period to be from 16 May to 20 June, based on behavioral observations collected in our study area during the identical field season (M. Gaylord, personal observation). Furthermore, we only included detections that were manually verified (i.e., true positives) in this analysis to meet model assumptions. Occupancy models were fitted by using program R 4.0.5 (R Development Core Team 2021) with packages unmarked 1.1.1 (Fiske and Chandler 2011, Chandler et al. 2023) and MuMIn 1.43.17 (Bartoń 2020).
We related occupancy and detection probabilities to explanatory variables using logistic regression models. Although our objectives for this study focused on detection probability, we considered covariates on occupancy probability (ψ) to attempt to capture heterogeneity in the data associated with variation in occupancy to meet model assumptions. The occupancy probability covariates that we considered included habitat suitability (low [0.0–0.3], intermediate [0.3–0.5], and high [0.5–1.0]) at the survey station and the proximity (low [> 150 m], intermediate [< 150 to > 50 m], high [< 50 m]) to highly suitable habitat (0.5–1.0). These habitat suitability scores came from a raster layer (30 × 30 m resolution) that was developed by Latif et al. (2015), where they used White-headed Woodpecker callback surveys on the eastern side of the Cascade Mountain Range, Oregon, USA, to estimate habitat suitability across forest conditions. They found that habitat suitability at both the local and landscape scale may be important factors influencing White-headed Woodpecker occurrence, which is why we considered habitat conditions based on suitability at the survey station and distance from the survey station to the nearest area with high habitat suitability. Furthermore, we discretized the suitability values into categories (rather than leaving them as continuous values) to better align with how this variable is used within a management context. Detection probability (p) covariates that we considered included day of year, average temperature, the interaction between day of year and average temperature, and survey method (i.e., callback survey versus ARU). Daily temperature data were collected from the nearest weather station located in Union, Oregon. Day of year, average daily temperature, and the interaction between day of year and average daily temperature were included because we hypothesized that White-headed Woodpecker calling rates would decline at higher temperatures (based on previous monitoring efforts for the species), call rates would differ because of nesting phenology, and the effect of temperature on call rates would be modified on the basis of nesting phenology. We included survey method as a covariate on detection probability in a similar way as Nichols et al. (2008) because evaluating this potential effect was one of the primary objectives of this study. We did not consider models with an interaction between survey method and environmental conditions (i.e., wind and rain) because we had no expectation that the detection probability would differ across environmental gradients (e.g., a drizzle versus a downpour) based on survey method, given that they both rely so heavily on auditory detections.
We employed an exploratory approach to model selection, where we fit models with every combination of covariates. Preliminary analyses indicated there were no issues with multicollinearity. We compared candidate models using Akaike’s Information Criterion corrected for small sample size (AICc; Burnham and Anderson 2002). We considered models to have similar support if the ΔAICc was ≤ 2 and interpreted our output using predicted probabilities and odds ratios that were calculated from model coefficients (Hosmer and Lemeshow 2000). Candidate models were compared by using program R 4.0.5 (R Development Core Team 2021) and the AICmodavg 2.3-1 package (Mazerolle and Mazerolle 2017).
We used our most supported model to estimate detection probabilities for each day that each specific survey type took place. We then used a method-of-moments estimator for univariate random-effects meta-analysis (Jackson et al. 2013) to combine these estimates into an overall mean and variance for detection probabilities for each survey method. This was done in program R 4.0.5 (R Development Core Team 2021) by using the mvmeta R package (Gasparrini et al. 2012, Gasparrini and Armstrong 2013). The cumulative detection probability (p*) was calculated as p* = 1 - (1 - p)n, where n is the number of surveys and p is the detection probability for a specific survey type. We incorporated uncertainty in our estimates of p* using a parametric bootstrap simulation approach where we randomly sampled p from a beta distribution using the overall mean and variance estimates for detection probabilities for each survey method. We estimated the beta distribution shape parameters to sample from using the mean and variance estimates from our meta-analysis described above and a method-of-moments estimator. We considered scenarios where the number of surveys ranged from one to 10 in increments of one. Each of these scenarios were run for 10,000 iterations and summarized by the mean and 2.5th and 97.5th percentiles (hereafter, 95% confidence interval [CI]).
RESULTS
Most of our callback surveys started at 0830 hours and ended between 1100 and 1300 hours (2.5–4.5 h). Out of the 60 survey stations along transects, we detected a White-headed Woodpecker on at least one survey day across the breeding season at only 18% of the survey stations.
After processing all audio data with the classifier, the mean number of audio clips reviewed per ARU was 4490 (SD = 2180; Table 1). The average number of audio clips that were listened to before a verified White-headed Woodpecker call (e.g., true positive) was detected was 170 (range = 0–1389; Table 1). The estimated recall was 0.405 (SD = 0.197; 22.5 h), meaning that approximately 40.5% of the White-headed Woodpecker calls in our audio data were identified by the classifier. The precision was estimated at only 0.037 (4176 true positive/112,262 true and false positive calls), meaning that only 3.72% of the audio clips that were identified as White-headed Woodpeckers were actually White-headed Woodpecker calls and the rest were false positive detections (Table 1).
We reviewed and validated all recordings from nine ARUs with significant White-headed Woodpecker activity (> 100 detections from 5 May to 15 July; Table 1) across the recording times (0000–2310 hours). White-headed Woodpecker calls were detected as early as just before 0500 hours and no later than 2100 hours, indicating a resting period between 2100 hours to just before 0500 hours. During the continuous recording period (0000–1100 hours), we calculated the average number of calls recorded during a 10-min interval to make comparisons with the other recording events. Across the ARU recordings reviewed, we observed a spike in activity from 0500 to 0700 hours and 1700 to 1900 hours (Fig. 2).
Prior to fitting occupancy models, we removed any false positive detections from our data. Given that we only needed to validate a single White-headed Woodpecker vocalization per day per survey station for our occupancy analysis, it typically required 30–60 min for us to validate calls for each station for each recording day. After removing false positive detections, White-headed Woodpeckers were recorded and identified on 9.24 (SD 8.43) different days, on average, during the breeding season (16 May to 20 June; Table 1) at a survey station. Of the 25 ARU survey stations, we detected a White-headed Woodpecker on at least one recording day across the breeding season at 22 survey stations.
Our model selection procedure resulted in 40 candidate models. Those with an AICc model weight (wi) ≥ 0.01 are included in Table 2. The model selection process did not result in competing models (i.e., only one model had a ΔAICc ≤ 2). The top ranked model included no covariates for occupancy probability, but there was support for day of year and survey type influencing detection probability (Table 3). Occupancy probability (ψ) was estimated at 0.88 (95% CI, 0.68–0.96). The odds of detection were 4.41 times lower when using callback surveys along transects compared to using ARU surveys. The odds of detection were 1.28 times higher for every approximately 10 days that passed during the breeding season. The overall mean detection probability for an ARU survey day at a survey station was 0.46 (SD = 0.01), whereas the overall mean detection probability at a survey station along a transect survey was 0.15 (SD = 0.01). The cumulative detection probability at an ARU survey station ranged from 0.95 to 0.99 (95% CI, 0.953–0.959 to 0.993–0.994, respectively) after only recording for five to eight days, respectively (Fig. 3). On the other hand, the cumulative detection probability at a survey station along a transect was only 0.15–0.38 (95% CI, 0.126–0.181 to 0.357–0.427, respectively) after one to three callback survey(s).
DISCUSSION
We conducted the first, to our knowledge, comparative analysis between traditional callback surveys along transects and a relatively new passive acoustic monitoring design for White-headed Woodpeckers in forested landscapes. Based on our findings, callback surveys along a transect were significantly less reliable than ARU surveys at detecting White-headed Woodpecker vocalizations on a given day. Given that vocalizations are a primary data source for estimating White-headed Woodpecker habitat use, ARU surveys could be used in lieu of callback surveys along transects if the objective is to monitor White-headed Woodpecker occurrence patterns in relation to spatiotemporal variation in forest habitat conditions.
We were somewhat surprised that we found no support that occupancy was related to the habitat quality covariates we considered in our analyses; however, this could be related to several factors. First, we purposely chose to work in watersheds that had previous detections of White-headed Woodpeckers because our objectives for this study were centered on comparing detection probabilities between the two survey methods. As such, we needed White-headed Woodpeckers to be present at a sufficient number of survey stations to estimate and compare detection probabilities. This likely means our study area had higher habitat suitability, on average, than other areas of the forest and perhaps less variable forest habitat conditions. Second, there is a mismatch in the resolution of the habitat suitability raster layer developed by Latif et al. (2015; 30 × 30 m resolution) and the estimated core home range size (18.9 ha, SD = 10.1, range 3.1–35.7 ha) and use patterns of White-headed Woodpeckers during the breeding season (Koch et al. 1970, Ligon 1973, Dixon 1995, Kehoe 2017). This likely means that landscape-scale habitat suitability could be a greater influential parameter affecting White-headed Woodpecker occupancy than the local suitability at a survey station. Third, the underlying spatial data layers (gradient nearest neighbor [GNN]; Ohmann and Gregory 2002) used to estimate habitat suitability scores rely on Landsat imagery collected in 2000 and 2012, which could be outdated given the dynamic nature of managed forest landscapes (Latif et al. 2015). Last, large tree density is an important factor influencing White-headed Woodpecker habitat quality (Kozma et al. 2020), but this information was not included in the habitat suitability model used by Latif et al. (2015) because the GNN layer did not predict this forest habitat metric accurately at this scale for this region (Ohmann and Gregory 2002, Latif et al. 2015). Collectively, this suggests that the lack of a supported relationship between occupancy and habitat suitability is related to our study design and covariate data, and our findings do not indicate that White-headed Woodpeckers do not exhibit habitat preferences in forest landscapes.
Although we predicted that detection probability per day for the two survey types would be equal, we found the odds of detection were 4.41 times lower for callback surveys along transects than for ARU surveys. This meant that the cumulative detection probability when using callback surveys was only estimated at 0.81 (95% CI, 0.789–0.829) if 10 replicate surveys were conducted at the survey station (Fig. 3). Yet, callback surveys along transects that are conducted across national forests in this region to monitor White-headed Woodpeckers are typically not repeated more than two times during a breeding season and between the hours of sunrise and noon. Conversely, ARUs only needed to be recording at a survey station for five to eight days (13.2 h per day) to have a cumulative detection probability that approached one. The exact number of surveys required to infer absence at survey stations that yield no detections depends on the context of the management decision and the acceptability of risk by managers. However, our results indicate an ARU deployed in the field for a little over a week would likely detect White-headed Woodpeckers without fail. These results will only hold true when using the same recording schedule and window that we used, while reviewing every clip that was identified as a potential White-headed Woodpecker call.
Both survey types were influenced by observer bias to some extent. White-headed Woodpecker calls were likely missed by both survey types when many birds were vocalizing at the same time or when environmental factors (i.e., wind and rain) increased environmental noise. However, ARUs collected audio data for a much longer amount of time at a survey station and for each survey day (13.2 h per day) than field crews did while conducting callback surveys along transects (2.5–4.5 h per transect per day). During this study, callback surveys were often not started until 0830 hours and ended between 1100 and 1300 hours. These start and end times are typical for White-headed Woodpecker effectiveness monitoring across national forests and are the result of long drive times, rough terrain, difficulty tracking down possible White-headed Woodpecker calls, and training technicians on field protocols. Yet, peak hours of White-headed Woodpecker activity recorded on our ARUs indicate our callback surveys were started outside of peak White-headed Woodpecker calling activity. Surveyors could start callback surveys earlier to increase detection probabilities, although this is logistically challenging given the remote areas that White-headed Woodpeckers occupy. Increased detection probabilities might also be achieved by spending additional time at survey stations, but how much this would improve detection probability would require further analysis. Altogether, it makes sense that ARUs would have a significantly higher detection probability compared to callback surveys when a survey station is occupied because ARUs are surveying for a much greater number of hours per day and during peak White-headed Woodpecker activity. Perhaps a more direct comparison between these two survey methods would be to restrict our recording data each day to the same amount of time we spent at a survey station using callback surveys each morning. However, that approach would miss the fact that the use of passive ARUs affords us the opportunity to survey for longer periods each day at little to no extra investment. In practice, there does not seem to be a reason to narrow the daily recording schedule when the objective is to monitor the presence/absence of vocal species. This is why we opted to use the full recording schedule for our comparison herein. Undoubtedly, if callback surveys along transects were conducted for similar amounts of time each day, the estimated detection probabilities for each survey method would be a closer match. However, the resources required to invest in this level of effort for callback surveys is currently cost prohibitive.
Our results also indicated detection probability increased as we surveyed further into the breeding season. This is likely related to the reproductive ecology of the species. Both male and female White-headed Woodpeckers vocalize regularly during courtship and incubation, as well as pre- and post-fledging. They also both vocalize regularly during the breeding season while foraging, defending the nest, and communicating with newly fledged offspring. Parents take turns incubating eggs early in the breeding season (Audubon 2022). Once the eggs hatch later in the breeding season both parents feed the young. This means that earlier in the breeding season there is likely only one adult foraging at a time, potentially resulting in reduced detection probabilities.
We truncated our monitoring data to the nesting period to improve our ability to meet the assumption of closure for occupancy analysis. Although this is a standard approach when monitoring wildlife species at survey stations to estimate the distribution and detection of species, our random survey station placement sampling design means that some of our survey stations were likely at the edge of a White-headed Woodpecker territory. This can result in an availability issue, where the species is not always available for detection based on how they use space within a territory. Thus, our occupancy parameter should be interpreted as habitat use since we are estimating whether the species used the habitats around our survey stations at any point during the breeding season (Appel et al. 2023, Weldy et al. 2023). Although this is not a unique issue to the use of ARUs, what is unique is our ability to quantify how much of an issue this might be. Indeed, the temporal grain and extent of our sampling when using ARUs allowed us to quantify the intensity of use around survey stations based on the detection of calls (Table 1). With these data, it would be worthwhile to fit multistate occupancy models, where the different states represent the intensity of use or the call detection rates (Nichols et al. 2007). This approach provides robust inferences when using count data to track changes in population abundance (Steen et al. 2023), but to our knowledge it has not been applied to ARU monitoring data to quantify spatial and temporal patterns in the intensity of habitat use across landscapes.
One complication we had with our classifier was that White-headed Woodpecker peak time of vocalization activity (0500 to 0700 hours) corresponds with many other avian species peak time of vocalization, which resulted in many polyphonic audio clips (i.e., clips that contain multiple sound types) that were difficult to accurately identify by the classifier. By targeting vocalizations that were good examples of White-headed Woodpecker calls, the recall dropped; however, the number of false positive detections also dropped. Having increased recall is important to avoid overestimating species occurrence by confusing its call with other sound types, including other species’ vocalizations (Priyadarshani et al. 2018). We accepted this tradeoff of a slightly higher recall and lower precision for two main reasons. First, false absences are statistically accounted for when analyzing the data by using estimators such as occupancy models, but false positive detections require auxiliary data to accurately estimate occupancy (reviewed in Chambert et al. 2015). This means that detection probabilities for ARUs might increase as we reduce false absences in our classifier moving forward. Second, we only needed to detect a single White-headed Woodpecker vocalization out of the 13.2 h of audio data recorded per day to correctly classify a survey station as occupied each day. Even with relatively high recall (0.405; SD = 0.197; 22.5 h) and low precision (0.037; 4176 true positive/112,262 true and false positive calls) our classifier saved us time (i.e., it typically required 30–60 min to review 13.2 h of audio), despite needing to manually verify White-headed Woodpecker calls (i.e., remove false positives) for our analysis, because it indicated where to look for calls within the audio recordings. Nevertheless, the recall could be improved in future classifiers by increasing the number of example vocalizations in the training data as well as reducing environmental noise by increasing the distance that ARUs were deployed from large creeks. Similarly, the precision of the classifier could be improved by developing a community-based classifier that includes vocalizations from different species across the Blue Mountains (Ruff et al. 2020, Kahl et al. 2021). Last, we could also extend the classifier to include other White-headed Woodpecker sounds, such as drums. Collectively, this would increase the classifier’s ability to differentiate between species with similar vocalizations, account for changes in environmental noise, and provide a greater range of White-headed Woodpecker sound types that might change based on season or by location.
CONCLUSION
This study provides information that can be used by managers to make informed decisions about what type of survey method to use and the effort required to detect White-headed Woodpeckers. Both survey types have costs and benefits. Callback surveys along transects allow field crews to locate nest sites and then perform follow up surveys to determine nest success. However, our results indicate that quite a few repeat callback surveys are needed to infer absence and that these surveys would benefit from earlier start times (i.e., starting at sunrise instead of 0830). Extending the number of hours a surveyor is in the field may also help, especially during courtship, if additional resources are available (e.g., more personal, funding, etc.). ARUs can be used to quantify status and trend in White-headed Woodpecker occupancy across larger landscapes with reduced field crew time and increased detection probabilities compared to transect surveys. Further, our results indicate ARUs can be rotated across multiple survey stations, rather than remaining at static locations across the breeding season, and still have high cumulative detection probabilities. With an increased interest in utilizing ARUs for surveying across landscapes and the need to monitor the greater forest wildlife community, improvements in the classifier we outline above (including extending it to identify other vocal species) would be beneficial. Furthermore, the classifier we developed could be used to process audio data across other forests given similar wildlife species composition in the region (e.g., ponderosa pine–dominant forests located east of the Cascade Mountain Range).
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
Thanks to D. Smith, USDA Forest Service Wallowa-Whitman National Forest for his help in conducting surveys in the field. N. Cloud, USDA Forest Service Colville National Forest and colleagues at the USDA Forest Service Pacific Northwest Research Station Bioacoustics Laboratory including D. Lesmeister and Z. Ruff, and Wildlife Acoustics support team who provided crucial information about building an advanced classifier using Kaleidoscope Pro 5. This project was funded by the Collaborative Forest Landscape Restoration Program (CFLRP), the Wallowa-Whitman National Forest, and the USDA Forest Service Pacific Northwest Research Station. The use of trade or firm names in this publication is for reader information and does not imply endorsement by the US Government of any product or service.
DATA AVAILABILITY
The data that support the findings of this study are available from the corresponding author upon request.
LITERATURE CITED
Altman, B., and B. Bresson. 2017. Conservation of landbirds and associated habitats and ecosystems in the Northern Rocky Mountains of Oregon and Washington. Version 2.0. Oregon-Washington Partners in Flight and American Bird Conservancy and U.S. Forest Service/Bureau of Land Management.
Appel, C. A., D. B. Lesmeister, A. Duarte, R. J. Davis, M. J. Weldy, and T. Levi. 2023. Using passive acoustic monitoring to estimate Northern Spotted Owl landscape use and pair occupancy. Ecosphere 14(2):e4421. https://doi.org/10.1002/ecs2.4421
Audubon. 2022. White-headed Woodpeckers (Dryobates albolarvatus). Guide to North American birds. https://www.audubon.org/field-guide/bird/white-headed-woodpecker
Bartoń, K. 2020. MuMIn: multi-model inference. R package version 1.43.17. https://cran.r-project.org/web/packages/MuMIn/index.html
Bate, L. J., M. J. Wisdom, and B. C. Wales. 2007. Snag densities in relation to human access and associated management factors in forests of northeastern Oregon, USA. Landscape and Urban Planning 80(3):278-291. https://doi.org/10.1016/j.landurbplan.2006.10.008
Buchanan, J. B., R. E. Rogers, D. J. Pierce, and J. E. Jacobson. 2003. Nest-site habitat use by White-headed Woodpeckers in the eastern Cascade Mountains, Washington. Northwestern Naturalist 84(3):119-128. https://doi.org/10.2307/3536537
Bull, E. L. 2002. The value of coarse woody debris to vertebrates in the Pacific Northwest. Pages 171-178. Proceedings of the symposium on the ecology and management of dead wood in Western Forests. General technical report PSW-GTR-181. USDA Forest Service, Pacific Southwest Research Station, Albany, California, USA.
Burnham, K. P., and D. R. Anderson. 2002. Model selection and multi-model inference: a practical information-theoretic approach. Second edition. Springer, New York, New York, USA.
Chambert, T., D. A. W. Miller, and J. D. Nichols. 2015. Modeling false positive detections in species occurrence data under different study designs. Ecology 96(2):332-339. https://doi.org/10.1890/14-1507.1
Chandler, R., K. Kellner, I. Fiske, D. Miller, A. Royle, J. Hostetler, R. Hutchinson, A. Smith, M. Kery, M. Meredith, et al. 2023. unmarked: models for data from unmarked animals. Version 1.3.2. https://cran.r-project.org/web/packages/unmarked/index.html
Dixon, R. D. 1995. Density, nest-site and roost-site characteristics, home-range, habitat use, and behavior of White-headed Woodpeckers: Deschutes and Winema National Forests, Oregon. Technical report 93-3-01. Oregon Department of Fish and Game. Portland, Oregon, USA.
Environmental Systems Research Institute. 2022. ArcGIS Desktop 10.7.1 quick start guide. https://desktop.arcgis.com/en/quick-start-guides/10.7/arcgis-desktop-quick-start-guide.htm#ESRI_SECTION1_5BE6C58891E046E48ACD2C3E37FE359B
Fiske, I., and R. Chandler. 2011. unmarked: an R package for fitting hierarchical models of wildlife occurrences and abundance. Journal of Statistical Software 43(10):1-23. https://doi.org/10.18637/jss.v043.i10
Franklin, J. F., and C. T. Dyrness. 1973. Natural vegetation of Oregon and Washington. General technical report PNW-GTR-008. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Portland, Oregon, USA.
Gasparrini, A., B. Armstrong, and M. G. Kenward. 2012. Multivariate meta-analysis for non-linear and other multi-parameter associations. Statistics in Medicine 31(29):3821-3839. https://doi.org/10.1002/sim.5471
Gasparrini, A., and B. Armstrong. 2013. Reducing and meta-analyzing estimates from distributed lag non-linear models. BMC Medical Research Methodology 13:1. https://doi.org/10.1186/1471-2288-13-1
Harrod, R. J., W. L. Gaines, W. E. Hartl, and A. Camp. 1998. Estimating historical snag density in dry forests east of the Cascade Range. General technical report PNW-GTR-428. U.S. Forest Service, Pacific Northwest Research Station, Portland, Oregon, USA. https://doi.org/10.2737/PNW-GTR-428
Hosmer, D. W., and S. Lemeshow. 2000. Applied logistic regression. Second edition. Wiley, New York, New York, USA.
Idaho Department of Fish and Game. 2021. White-headed Woodpecker (Picoides albolarvatus). Idaho Department of Fish and Game. https://idfg.idaho.gov/species/taxa/15618
Jackson, D., I. R. White, and R. D. Riley. 2013. A matrix‐based method of moments for fitting the multivariate random effects model for meta‐analysis and meta‐regression. Biometrical Journal 55(2):231-245. https://doi.org/10.1002/bimj.201200152
Kahl, S., C. M. Wood, M. Eibl, and H. Klinck. 2021. BirdNET: A deep learning solution for avian diversity monitoring. Ecological Informatics 61:101236. https://doi.org/10.1016/j.ecoinf.2021.101236
Kehoe, A. R. 2017. Space use and foraging patterns of the White-headed Woodpecker in western Idaho. Thesis, Montana State University, Bozeman, Montana, USA.
Koch, R. F., A. E. Courchesne, and C. T. Collins. 1970. Sexual differences in foraging behavior of White-headed Woodpeckers. Bulletin of the Southern California Academy of Sciences 69(1):60-64.
Kozma, J. M., T. J. Lorenz, M. G. Raphael, K. L. Garrett, and R. D. Dixon. 2020. White-headed Woodpecker (Dryobates albolarvatus). Version 2.0. In P. G. Rodewald and B. K. Keeney, editors. Birds of the World. Cornell Lab of Ornithology, Ithaca, New York, USA.
Lambeck, R. J. 1997. Focal species: a multi‐species umbrella for nature conservation. Conservation Biology 11(4):849-856. https://doi.org/10.1046/j.1523-1739.1997.96319.x
Lambeck, R. J. 1999. Landscape planning for biodiversity conservation in agricultural regions: a case study from the wheatbelt of western Australia. Biodiversity technical paper 2. Environment Australia, Canberra, Australia.
Latif, Q. S., V. A. Saab, J. G. Dudley, A. Markus, and K. Mellen-McLean. 2020. Development and evaluation of habitat suitability models for nesting White-headed Woodpecker (Dryobates albolarvatus) in burned forest. PLoS ONE 15(5):e0233043. https://doi.org/10.1371/journal.pone.0233043
Latif, Q. S., V. A. Saab, K. Mellen-Mclean, and J. G. Dudley. 2015. Evaluating habitat suitability models for nesting White-headed Woodpeckers in unburned forest. Journal of Wildlife Management 79(2):263-273. https://doi.org/10.1002/jwmg.842
Latif, Q. S., V. A. Saab, K. Mellen-Mclean, and J. G. Dudley. 2017. White-headed Woodpecker occupancy in the Pacific Northwest Region (U.S. Forest Service R6). Final 2017 progress report. U.S. Forest Service Pacific Northwest Region; U.S. Forest Service Rocky Mountain Research Station, Bozeman, Montana and Boise, Idaho, USA.
Ligon, J. D. 1973. Foraging behavior of the White-headed Woodpecker in Idaho. Auk 90(4):862-869. https://doi.org/10.2307/4084366
Lorenz, T. J., K. T. Vierling, J. M. Kozma, J. E. Millard, and M. G. Raphael. 2015. Space use by White-headed Woodpeckers and selection for recent forest disturbances. Journal of Wildlife Management 79(8):1286-1297. https://doi.org/10.1002/jwmg.957
MacKenzie, D. I., J. D. Nichols, G. B. Lachman, S. Droege, J. A. Royle, and C. A. Langtimm. 2002. Estimating site occupancy rates when detection probabilities are less than one. Ecology 83(8):2248-2255. https://doi.org/10.1890/0012-9658(2002)083[2248:ESORWD]2.0.CO;2
Mazerolle, M. J., and M. M. J. Mazerolle. 2017. Package ‘AICcmodavg.’ R package 281.
Mellen-McLean, K., V. Saab, B. Bresson, B. Wales, A. Markus, and K. VanNorman. 2015. White-headed Woodpecker monitoring strategy and protocols. Version 1.3. U.S. Department of Agriculture, Forest Service, Region 6 & Rocky Mountain Research Station. U.S. Department of Interior, Bureau of Land Management, Oregon and Washington, USA.
Nichols, J. D., L. L. Bailey, A. F. O’Connell, Jr., N. W. Talancy, E. H. Campbell Grant, A. T. Gilbert, E. M. Annand, T. P. Husband, and J. E. Hines. 2008. Multi‐scale occupancy estimation and modelling using multiple detection methods. Journal of Applied Ecology 45(5):1321-1329. https://doi.org/10.1111/j.1365-2664.2008.01509.x
Nichols, J. D., J. E. Hines, D. I. Mackenzie, M. E. Seamans, and R. J. Gutiérrez. 2007. Occupancy estimation and modeling with multiple states and state uncertainty. Ecology 88(6):1395-1400. https://doi.org/10.1890/06-1474
Ohmann, J. L., and M. J. Gregory. 2002. Predictive mapping of forest composition and structure with direct gradient analysis and nearest-neighbor imputation in coastal Oregon, U.S.A. Canadian Journal of Forest Research 32(4):725-741. https://doi.org/10.1139/x02-011
Oregon Department of Fish and Wildlife. 2021. White-headed Woodpeckers. The Oregon Conservation Strategy. https://oregonconservationstrategy.org/strategy-species/white-headed-woodpecker/
Pacifici, K., T. R. Simons, and K. H. Pollock. 2008. Effects of vegetation and background noise on the detection process in auditory avian point-count surveys. Auk 125(3):600-607. https://doi.org/10.1525/auk.2008.07078
Partners in Flight. 2019. Population estimates database. Version 3.0. https://pif.birdconservancy.org/population-estimates-database/
Powell, D. C. 2011. Active management of dry forests in the Blue Mountains: silvicultural considerations. Supervisor’s Office. U.S. Department of Agriculture, U.S. Forest Service, Pendleton, Oregon, USA.
Priyadarshani, N., S. Marsland, and I. Castro. 2018. Automated birdsong recognition in complex acoustic environments: a review. Journal of Avian Biology 49(5):jav-01447. https://doi.org/10.1111/jav.01447
R Development Core Team. 2021. R: a language and environmental for statistical computing. R 4.0.5. R Foundation for Statistical Computing, Vienna, Austria. https://ftp.osuosl.org/pub/cran/
Ruff, Z. J., D. B. Lesmeister, C. L. Appel, and C. M. Sullivan. 2021. Workflow and convolutional neural network for automated identification of animal sounds. Ecological Indicators 124:107419. https://doi.org/10.1016/j.ecolind.2021.107419
Ruff, Z. J., D. B. Lesmeister, L. S. Duchac, B. K. Padmaraju, and C. M. Sullivan. 2020. Automated identification of avian vocalizations with deep convolutional neural networks. Remote Sensing in Ecology and Conservation 6(1):79-92. https://doi.org/10.1002/rse2.125
Steen, V. A., A. Duarte, and J. T. Peterson. 2023. An evaluation of multistate occupancy models for estimating relative abundance and population trends. Ecological Modelling 478:110303. https://doi.org/10.1016/j.ecolmodel.2023.110303
Swiston, K. A., and D. J. Mennill. 2009. Comparison of manual and automated methods for identifying target sounds in audio recordings of Pileated, Pale-billed, and putative Ivory-billed Woodpeckers. Journal of Field Ornithology 80(1):42-50. https://doi.org/10.1111/j.1557-9263.2009.00204.x
Tyre, A. J., B. Tenhumberg, S. A. Field, D. Niejalke, K. Parris, and H. P. Possingham. 2003. Improving precision and reducing bias in biological surveys: estimating false‐negative error rates. Ecological Applications 13(6):1790-1801. https://doi.org/10.1890/02-5078
United States Department of Agriculture (USDA). 2008. Updated regional forester’s sensitive animal list. 2670/1950 memo to forest supervisors. July 2004. U.S. Department of Agriculture, U.S. Forest Service, Pacific Northwest, Region 6, Portland, Oregon, USA.
Van Wilgenburg, S. L., P. Sólymos, K. J. Kardynal, and M. D. Frey. 2017. Paired sampling standardizes point count data from humans and acoustic recorders. Avian Conservation and Ecology 12(1):13. https://doi.org/10.5751/ACE-00975-120113
Washington Department of Fish and Wildlife. 2021. White-headed Woodpecker (Picoides albolarvatus). https://wdfw.wa.gov/species-habitats/species/dryobates-albolarvatus
Weldy, M. J., D. B. Lesmeister, C. B. Yackulic, C. L. Appel, C. McCafferty, and J. D. Wiens. 2023. Long-term monitoring in transition: resolving spatial mismatch and integrating multistate occupancy data. Ecological Indicators 146:109815. https://doi.org/10.1016/j.ecolind.2022.109815
Western Regional Climate Center. 2022. Climate of Oregon: topographic features. https://wrcc.dri.edu/Climate/narrative_or.php
Wildlife Acoustics. 2020. Kaleidoscope Pro 5 user guide. https://www.wildlifeacoustics.com/uploads/user-guides/Kaleidoscope-Pro-User-Guide.pdf
Wisdom, M. J., and L. J. Bate. 2008. Snag density varies with intensity of timber harvest and human access. Forest Ecology and Management 255(7):2085-2093. https://doi.org/10.1016/j.foreco.2007.12.027
Youngblood, A. 2000. Consequences of fire and fire surrogate treatments-The Hungry Bob Project, Wallowa-Whitman National Forest. U.S. Department of Agriculture, Forest Service Pacific Northwest Research Station. La Grande, Oregon, USA.
Fig. 1
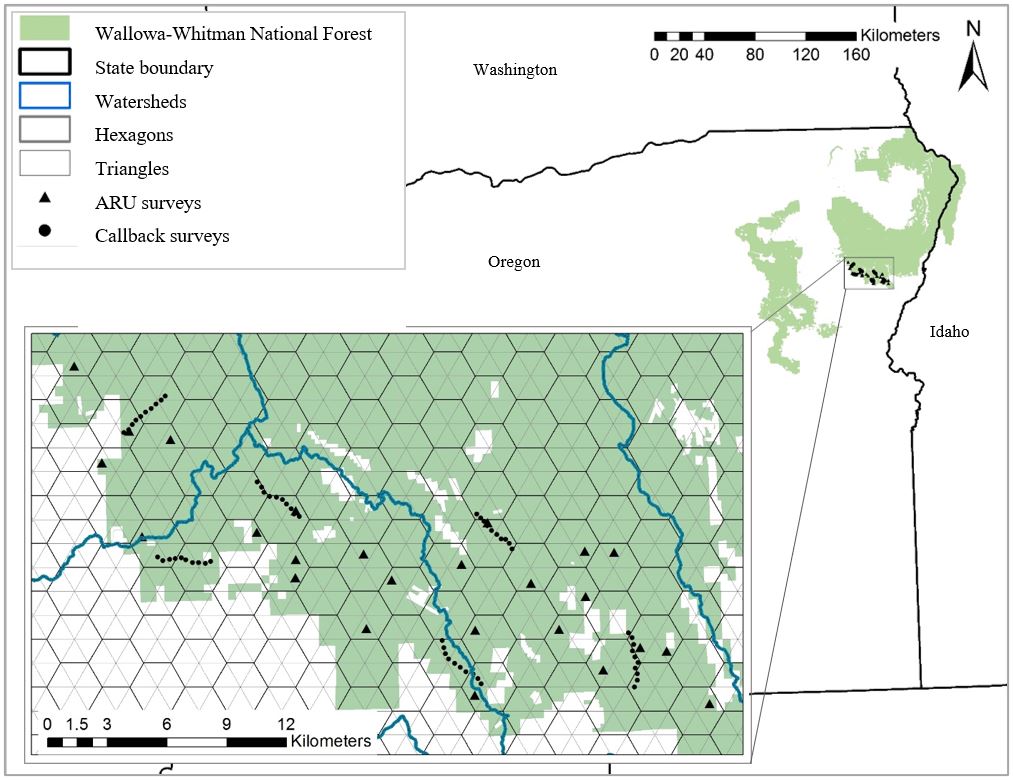
Fig. 1. Study area extent including White-headed Woodpecker (Dryobates albolarvatus) transect survey stations (n = 60) and passive acoustic recording unit (ARU) survey stations (n = 25). The overlayed 500-ha hexagon grid with six 83-ha triangles was used as a survey frame when selecting ARU survey station locations. Wallowa-Whitman National Forest, northeastern Oregon, USA, 2021.
Fig. 2
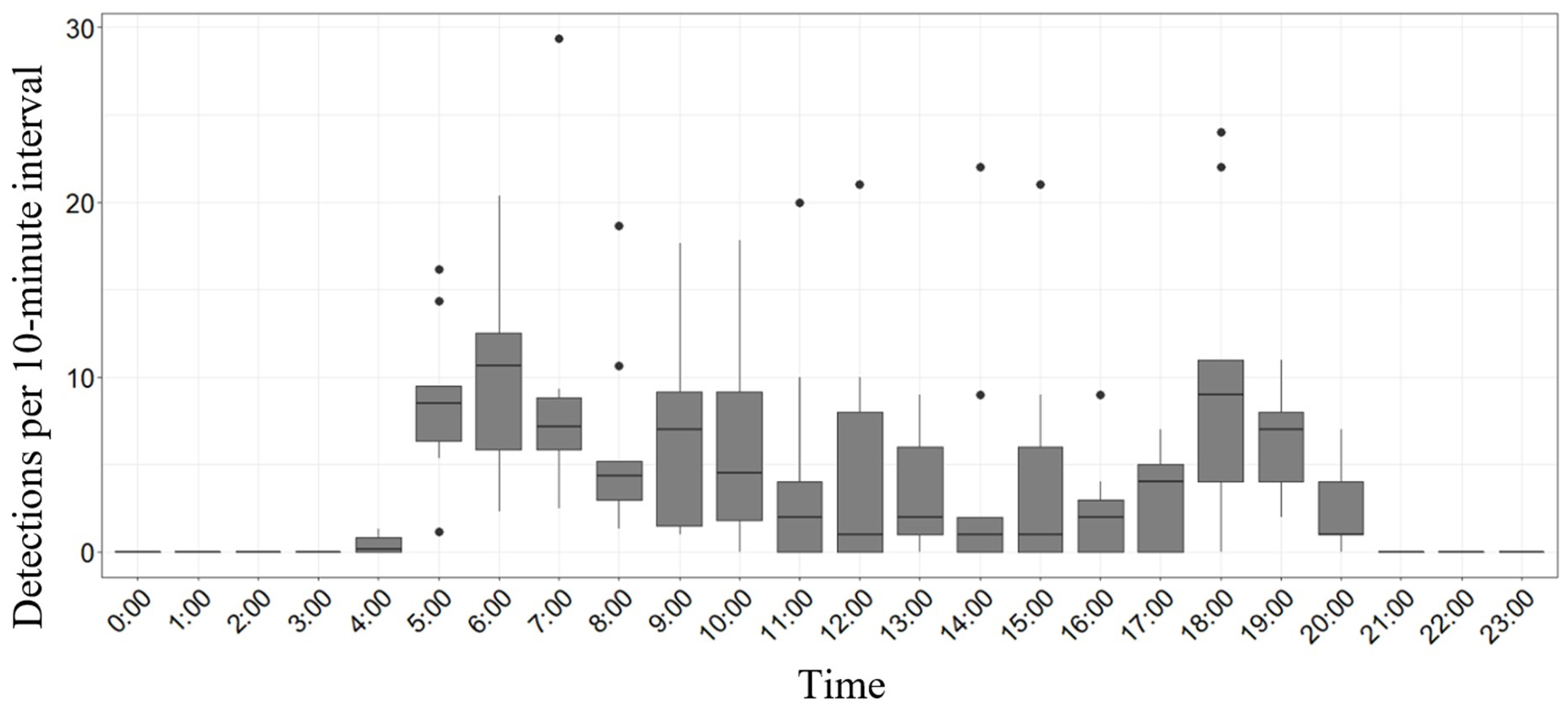
Fig. 2. The number of White-headed Woodpecker (Dryobates albolarvatus) vocalizations that were initially identified by our classifier and manually verified at nine passive acoustic recording unit stations that had at least 100 detections. The number of detections during hour-long recordings were divided by six to account for variable recording lengths throughout the day. Sunrise was approximately 0507–0540 hours and sunset was approximately 2000–2033 hours. Wallowa-Whitman National Forest, northeastern Oregon, USA, 2021.
Fig. 3
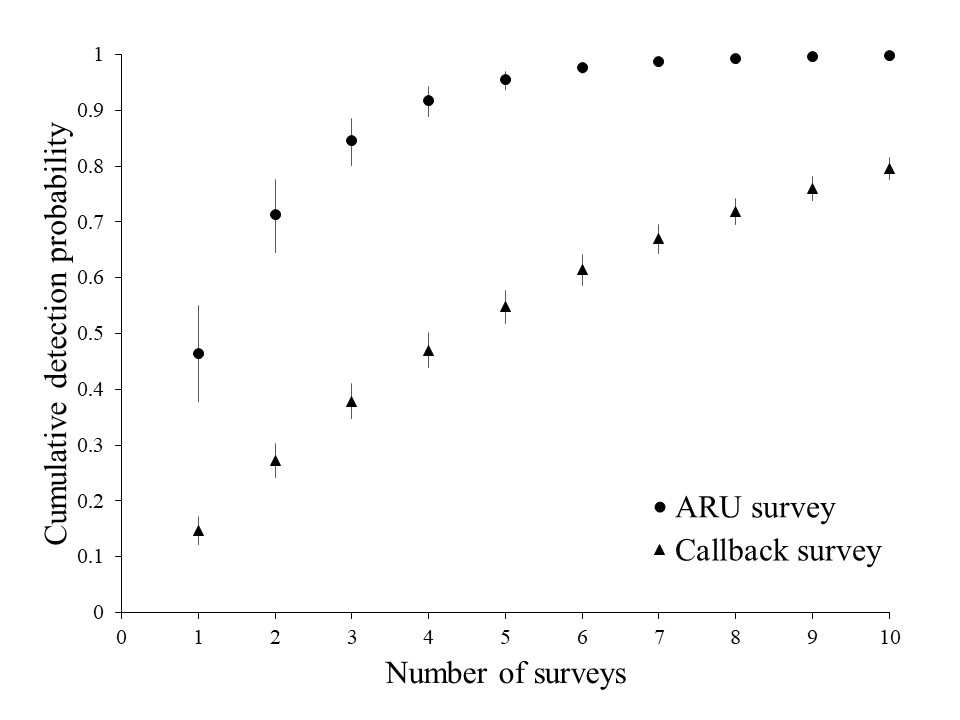
Fig. 3. Cumulative detection probability with 95% confidence interval (CI) of White-headed Woodpecker (Dryobates albolarvatus) using callback surveys at a survey station along a transect versus detecting a vocalization at a survey station using a passive acoustic recording unit. Wallowa-Whitman National Forest, northeastern Oregon, USA, 2021.
Table 1
Table 1. Detection summaries for each passive acoustic recording unit (ARU; 001–025) processed with our classifier. Notably, these detections represent only verified detections that were initially identified by our classifier. Thus, detection numbers are an underestimate of the number of clips with target class recordings at each station. Wallowa-Whitman National Forest, northeastern Oregon, USA, 2021.
| ARU | Number of audio clips reviewed | Detections | Days with detections (5 May to 15 July) | Days with detections (16 May to 20 June) | Days ARUs recorded (16 May to 20 June) | Reviewed before first verified detection | |||
| 001 | 4100 | 88 | 24 | 16 | 19 | 0 | |||
| 002 | 2086 | 155 | 24 | 24 | 27 | 4 | |||
| 003 | 1200 | 25 | 9 | 3 | 32 | 71 | |||
| 004 | 5401 | 142 | 21 | 13 | 26 | 0 | |||
| 005 | 4109 | 30 | 7 | 6 | 27 | 22 | |||
| 006 | 7417 | 464 | 48 | 31 | 33 | 0 | |||
| 007 | 4353 | 5 | 3 | 3 | 24 | 416 | |||
| 008 | 4430 | 298 | 23 | 17 | 18 | 39 | |||
| 009 | 2140 | 1 | 1 | 0 | 7 | 235 | |||
| 010 | 7412 | 38 | 8 | 5 | 26 | 284 | |||
| 011 | 2665 | 6 | 3 | 3 | 14 | 1004 | |||
| 012 | 137 | 1 | 1 | 0 | 28 | 28 | |||
| 013 | 4471 | 357 | 27 | 17 | 23 | 5 | |||
| 014 | 7821 | 11 | 3 | 3 | 27 | 4 | |||
| 015 | 3579 | 17 | 2 | 1 | 24 | 252 | |||
| 016 | 5171 | 66 | 10 | 6 | 20 | 28 | |||
| 017 | 5048 | 22 | 8 | 4 | 28 | 8 | |||
| 018 | 7986 | 1344 | 36 | 14 | 15 | 1 | |||
| 019 | 4144 | 491 | 33 | 19 | 19 | 2 | |||
| 020 | 3515 | 1 | 1 | 0 | 20 | 1389 | |||
| 021 | 6249 | 48 | 12 | 4 | 12 | 97 | |||
| 022 | 2357 | 13 | 4 | 1 | 24 | 256 | |||
| 023 | 4898 | 200 | 12 | 8 | 10 | 58 | |||
| 024 | 8868 | 288 | 29 | 21 | 34 | 4 | |||
| 025 | 2705 | 66 | 12 | 12 | 22 | 42 | |||
Table 2
Table 2. Akaike’s Information Criterion corrected for small sample size (AICc), change in AICc (ΔAICc), model weights (wi), and the number of parameters (k) for the detection probability (p) and occupancy probability (ψ) for White-headed Woodpecker (Dryobates albolarvatus). Wallowa-Whitman National Forest, northeastern Oregon, USA, 2021.
| Model | k | AICc | ΔAICc | wi | |||||
| ψ (.) p (day of year + survey) | 4 | 791.70 | 0.00 | 0.50 | |||||
| ψ (.) p (day of year + survey + temperature) | 5 | 793.94 | 2.24 | 0.16 | |||||
| ψ (habitat) p (day of year + survey) | 6 | 795.31 | 3.61 | 0.08 | |||||
| ψ (proximity) p (day of year + survey) | 6 | 795.63 | 3.93 | 0.07 | |||||
| ψ (.) p (day of year + survey + temperature + day of year:temperature) | 6 | 796.11 | 4.41 | 0.05 | |||||
| ψ (.) p (survey) | 3 | 797.24 | 5.54 | 0.03 | |||||
| ψ (habitat) p (day of year + survey + temperature) | 7 | 797.68 | 5.99 | 0.02 | |||||
| ψ (proximity) p (day of year + survey + temperature) | 7 | 798.00 | 6.30 | 0.02 | |||||
| ψ (.) p (survey + temperature) | 4 | 798.73 | 7.04 | 0.01 | |||||
| ψ (habitat + proximity) p (day of year + survey) | 8 | 799.53 | 7.83 | 0.01 | |||||
| ψ (habitat) p (day of year + survey + temperature + day of year:temperature) | 8 | 800.01 | 8.31 | 0.01 | |||||
| ψ (proximity) p (day of year + survey + temperature + day of year:temperature) | 8 | 800.30 | 8.60 | 0.01 | |||||
| ψ (habitat) p (survey) | 5 | 800.73 | 9.03 | 0.01 | |||||
Table 3
Table 3. The mean, standard error (SE), and 95% confidence intervals (CI) for occupancy probability (ψ) and detection probability (p) for White-headed Woodpecker (Dryobates albolarvatus) during the breeding season (16 May through 20 June). Wallowa-Whitman National Forest, northeastern Oregon, USA, 2021.
| Parameter | Mean | SE | Lower CI | Upper CI | |||||
| Occupancy probability (ψ) | |||||||||
| Intercept | 1.95 | 0.622 | 0.731 | 3.171 | |||||
| Detection probability (p) | |||||||||
| Intercept | -0.170 | 0.090 | -0.347 | 0.007 | |||||
| Day of year | 0.243 | 0.880 | 0.071 | 0.416 | |||||
| Survey type: transect | -1.484 | 0.334 | -2.140 | -0.829 | |||||