







The following is the established format for referencing this article:
Miller, S. M., J. Schoen, M. W. Reudink, and S. M. Mahoney. 2023. Disentangling the mechanisms of signal evolution in Tyrannidae flycatchers, part II: plumage elaboration evolved with migration behavior, but is also affected by diet, climate, and drift. Journal of Field Ornithology 94(3):7.ABSTRACT
Animal coloration is an important communication signal that varies among taxa and affects survival and reproduction. Species-specific color is influenced by a variety of factors including phylogeny, predation, sexual selection, light and resource availability, ecological context, and/or species recognition. Tyrant flycatchers are useful to study color evolution because they are the largest family of birds, occur across broad environmental gradients, and although many genera are monomorphic with drab plumage, some are very colorful. Given the complexity of factors involved, the mechanisms influencing plumage color evolution in Tyrant flycatchers likely involve multiple drivers. In the second of our two-part paper on signal evolution in this avian family, we harnessed a large plumage color database of female and male Tyrannidae species (n = 399) to test the relative importance of geography (climate, latitude), ecology (migration behavior, forest cover, diet), and heterospecific proximity on plumage color evolution. From phylogenetically controlled analyses, we found that female color was largely driven by climate and male color was more exaggerated in migratory species. Sexual dichromatism was also affected by climate and diet, and was more pronounced in migratory species, possibly as a result of color loss in females. Pairwise comparisons between heterospecific color differences and geographic distance were generally weak and consistent with expectations under drift, which contrasted our song results (presented in Part I), but analyses at finer taxonomic scales revealed color may be used for conspecific recognition in some genera. Together with our paper on Tyrannidae song evolution, the color results reflect the importance of testing multiple drivers of signal evolution at different taxonomic scales because flycatcher color variation may have arisen through genetic drift, from a loss of female color in migratory species, and for species recognition in some genera. Future work could formally test the correlation between song and color variation, and use stochastic character mapping to confirm if Tyrannidae dichromatism resulted from female color loss in migratory species.
RESUMEN
La coloración en los animales es una señal importante en la comunicación que varía entre taxones y afecta la supervivencia y la reproducción. El color específico a la especie esta influenciado por una variedad de factores incluyendo la filogenia, depredación, selección sexual, luz y disponibilidad de recursos, contexto ecológico y/o reconocimiento de especies. Los atrapamoscas son útiles para estudiar la evolución del color, porque son la familia más grande de aves, ocurren a lo largo de amplios gradientes ambientales y, a pesar que muchos géneros son monomórficos con plumajes poco vistosos, algunos son muy coloridos. Dada la complejidad en los factores involucrados, los mecanismos que influencian la evolución en la coloración del plumaje de atrapamoscas probablemente involucran múltiples determinantes. En la segunda parte de nuestro estudio enfocado en la evolución de las señales en esta familia de aves, utilizamos una gran base de datos de coloración del plumaje de machos y hembras de la familia Tyrannidae (n=399), con el fin de poner a prueba la importancia relativa de la geografía (clima, latitud), la ecología (comportamiento migratorio, cobertura boscosa, dieta) y proximidad hetero específica sobre la evolución del color del plumaje. De los análisis controlados por filogenia, encontramos que el color de las hembras es mayoritariamente determinado por el clima y el color en los machos es más exagerado en especies migratorias. El dicromatismo sexual también estuvo influenciado por el clima y la dieta y fue más pronunciado en especies migratorias, probablemente como producto de la pérdida del color en las hembras. Comparaciones pareadas entre las diferencias en el color hetero-específico y la distancia geográfica fueron generalmente débiles y consistentes con las predicciones de la deriva, lo cual es contrastante con los resultados en el canto (presentado en la Parte I), pero los análisis a una escala taxonómica más fina revelaron que el color puede ser usado en el reconocimiento conespecífico en algunos géneros. En conjunto con nuestro estudio en la evolución del canto en Tyrannidae, los resultados del color reflejan la importancia de poner a prueba múltiples determinantes de la evolución de señales a escalas taxonómicas diferentes, pues la variación en el color en atrapamoscas pudo haberse originado por deriva genética por medio de una pérdida del color de las hembras en especies migratorias y para reconocimiento específico en algunos géneros. Futuros estudios pueden formalmente poner a prueba la correlación entre el canto y la variación en el color, y utilizar el mapeo estocástico de caracteres para confirmar si el dicromatismo en la familia Tyrannidae resultó de la perdida del color de las hembras en especies migratorias.
INTRODUCTION
In birds, feather colors and patterns are important visual signals that vary within and among species (Andersson 1994). Plumage evolution can be driven by both adaptive (natural and sexual selection) and neutral processes (Bradbury and Vehrencamp 1998). Natural selection can play an important role in shaping plumage coloration because plumage characteristics of birds may be under selection to match the environment and aid in crypsis or to help with thermoregulation (Wolf and Walsberg 2000). For example, Gloger’s rule (1833), which has received empirical support among bird studies (reviewed by Zink and Remsen 1986, Delhey 2019, but see Marcondes et al. 2021), predicts that more heavily pigmented animals should be found closer to the equator where it is more humid, and less pigmented animals should be found at higher latitudes where it is more arid. Humid areas are hypothesized to have higher bacteria loads and the heavily pigmented feathers found in these areas may be less susceptible to bacterial degradation (Burtt and Ichida 2004). Lighter colored feathers may also aid in crypsis in arid environments; however, a study testing Gloger’s rule with species exhibiting carotenoid-based pigments found the opposite pattern, in which upper plumage parts became lighter in more mesic habitats (Chui and Doucet 2009).
Ornamental colors can evolve through sexual selection when individuals prefer particular colors in mates, leading to higher relative fitness of colorful individuals and, ultimately, an elaboration of those traits (Hill 2015). In birds, females are typically the choosy sex and numerous experimental and observational studies have demonstrated that females prefer males with more elaborate plumage (Hill 2006a). Plumage can act as a direct indicator of male condition or quality, but elaborate plumage can also evolve simply through female preference for specific signals (e.g., through sensory bias, Prum 1990, 2010, Tobias and Hill 1998). Males with elaborate colors may be perceived as higher quality for example, because the acquisition of pigments (e.g., carotenoids) or producing feather nanostructure for structural coloration can be costly, meaning only individuals in good physiological condition during moult can produce the most colorful displays (Hill 2006a, Hill 2015, White 2020). Regardless of the mechanism, if colorful males are more successful at obtaining mates and females prefer more colorful males, elaborate plumage can quickly evolve (Hill 2015). Additionally, sexual versus natural selection can lead to plumage divergence within species, resulting in sexual dichromatism (Badyaev and Hill 2003). Dichromatism arises when there is stronger mate choice pressure on one sex within the species, which results in more elaborate ornamentation in that sex (Bailey 1978, Simpson et al. 2015). Simultaneously, dichromatism can arise through the loss of elaborate ornaments in one sex due to relaxation of sexual selection and/or increased selection for crypsis from predation risk during migration (Simpson et al 2015). At a different level of analysis, sexual selection can drive species radiations by driving the diversification of phenotypic traits associated with mate choice (Cooney et al. 2017). In the presence of strong intra-sex competition, and a change in inter-sex mate preference, the plumage signals that increase mating success can diverge, leading to reproductive isolation (Lande 1981).
Other ecological aspects of a species’ environment may influence color evolution. The sensory drive hypothesis predicts signal transmission and reception will optimize under specific environmental contexts (Boughman 2002). In the case of plumage coloration, environmental factors like light availability may affect signal transmission (Endler 1992, 1993). For example, birds that display carotenoid-based colors absorb short and medium wavelengths of light, resulting in transmission of longer wavelengths (red, orange, and yellow; Badyaev and Hill 2003, Hill 2006a). Another way plumage color can be produced is through the interaction of light and nanostructures in the feathers. Composed of a combination of melanin, keratin, collagen, and other biomolecules, feather nanostructures interact with light and transmit blue, green, and iridescent colors (Prum 2006). Regardless of the plumage color, the light availability of an environment (e.g., open grassland vs. forest understory) will affect the conspicuousness of ornate plumage in the habitat (Endler 1992, Endler and Thery 1996).
When many species are living in sympatry and the need for species recognition is heightened, plumage signal divergence is magnified and can result in radiation events (Seddon et al. 2013). Birds that are closely related often have similar plumage color and patterns because of shared ancestry; however, overlapping ranges can cause signals to diverge (Sætre et al. 1997, Seddon 2005). This phenomenon could be explained by the species recognition hypothesis (Sætre et al. 1997, Simpson et al. 2021). In their study of Wood Warblers (Phylloscopus sibilatrix)), Simpson et al. (2021) found North American Wood Warblers with overlapping ranges exhibited greater plumage divergence. Similarly, dichromatism may evolve from a need for species recognition (Luro and Hauber 2022). Not all genera support the hypothesis because some congeners exhibit conserved plumage coloration even in sympatry, suggesting species recognition may be multi-modal (McNaught and Owens 2002).
Tyrant flycatchers are a unique group to study the evolution of visual signals because they occur across broad environmental gradients in North, South, and Central America and although there are many monomorphic species with drab plumage, some species are quite colorful (Fig. 1). The mix of conserved, drab plumage traits among many species and high elaboration in others, makes the group a promising subject to help unravel patterns of signal divergence. In our second paper of a two-part study on Tyrannidae signal evolution, we used phylogenetically controlled analyses to investigate how ecology (habitat, diet, migration behavior) and climate (precipitation and temperature) affected plumage color in Tyrant flycatchers to gain insight on how variation arose and is maintained. Additionally, we examined whether species living in closer proximity displayed greater color divergence as a test of the species recognition hypothesis. We predicted (1) plumage of both sexes would be largely driven by migration behavior, diet, climate, and habitat because these aspects have been found to influence coloration in other groups (e.g., Reudink et al. 2015, Simpson et al. 2015, Ward et al. 2021), and (2) we would find less support for the species recognition hypothesis because many Tyrant flycatchers exhibit similar plumage within genera but display pronounced vocal differences (investigated in Part I of this study). We further aimed to broaden the understanding of avian ecology and evolution in historically underrepresented regions because many of the species in our study occur exclusively in Central and South America (Nuñez et al. 2021). Our study also contributes to the growing body of literature on female signaling modalities by assessing female plumage evolution in addition to males (Riebel et al. 2019).
METHODS
Data collection
We obtained Tyrannidae plumage color data for 399 species (Appendix 1) from Dale et al. (2015), who digitally scanned images from the Handbook of the Birds of the World to score plumage color on males and females of each species from six patches (crown, forehead, nape, throat, upper breast, and lower breast), found to be consistently included in the illustrations and important for avian communication (Dale et al. 2015). Color patch values were measured from 400 randomly selected pixels to obtain red-green-blue (RGB) values (Dale et al. 2015). The RGB values were then corrected using species with similar coloration (see Dale et al. 2015) and averaged to provide single plumage color scores for males and females of each Tyrannidae species. To estimate dichromatism, we used the absolute values of the difference between the female color score from the male color score (Thibault et al. 2022). We previously used Dale et al. (2015)’s color scores to test different hypotheses about the evolutionary mechanisms of passeriform plumage coloration (Thibault et al. 2022).
Color scores provide a measurement for the general degree of color elaboration in each species and sex, where higher values indicate more colorful species, and are ideally suited for our large-scale comparative study of Tyrannidae (Dale et al. 2015). Human-biased color estimates provide useful proxies of color variation, particularly for interspecific comparative studies and for groups that are not UV sensitive (Seddon et al. 2010). Visual sensitivities for all Tyrant flycatchers are not known, however Brown-crested Flycatchers (Myiarchus tyrannulus), Eastern Phoebes (Sayornis phoebe), and Eastern Kingbirds (Tyrannus tyrannus) are violet sensitive (Ödeen and Håstad 2003, Aidala et al. 2012), suggesting UV signals are less important in Tyrannidae. This is consistent with the notion that UVS sensitive species typically communicate visually using structural blue plumage (Ödeen et al. 2012), a color rare in Tyrannidae (Gray 1996). Majority of Tyrannidae exhibit black, gray, and rufous plumage color, which typically exhibits modest amounts of UV reflectance (Eaton and Lanyon 2003). As a further test of the utility of their approach, Dale et al. (2015) verified color scores were highly correlated with UV-visual spectrometry, as measured on museum specimens (R² = 0.67, P < 0.0001; Dale et al. 2015, see their Extended Data Fig. 1 and Extended Data “Plumage scores validation analysis” section). Therefore, although our color score approach certainly underestimates color variation, we do not anticipate this has a large effect on assessing general interspecific color differences, and about the ecology and evolution of visual signals among Tyrannidae. We also endeavored to estimate carotenoid-based color in Tyrannidae by calculating chromaticity for each species (Methods and Results presented in Appendix 2, Dey et al. 2014).
Tyrannidae plumage color may be under selection to optimize transmission given the light conditions in the environment (Endler 1993). To test this hypothesis, in our models, we included estimates of forest cover from Mikula et al. (2020), a study that assessed acoustic adaptation of peak song frequency in all Passeriformes birds. Briefly, Mikula et al. (2020) estimated tree cover using data from the Copernicus Global Land Cover project (Buchhorn et al. 2020). For each geographic location of bird song recordings in their study, they extracted the percentage of tree cover in a 100 m² quadrant using the “exactextractr” package in R (Baston 2020). They then calculated species-specific tree cover percentages as the mean of all conspecific recordings in their dataset. In our analyses, we used the mean forest percentage cover for each Tyrannidae species from Mikula et al. (2020). The geographic extent of Mikula et al. (2020) broadly overlapped with that in our study, so the forest cover estimates are well suited to use here. Some species (n = 18) were missing forest cover data, so we imputed data using the mean value from congeners (n = 13) or the overall mean (n = 5).
Diet may also influence plumage color expression due to differences in resource availability (McGraw and Hill 2001), thus we included diet breadth in our models. We characterized diet breadth for each species using Wilman et al. (2014), who estimated the frequency of consumption of 10 major food types in which the frequency of use for each food type is recorded between 0 (never used) and 1 (approximately used exclusively). Data were unavailable for n = 4 species (Calyptura cristata, Piprites chloris, P. griseiceps, and P. pileata), so we gathered diet data from Birds of the World (https://birdsoftheworld.org). We then built a similarity matrix of nutritional content for each food type from Sayol et al. (2018) and calculated a breadth index using Rao’s quadratic entropy in the R package “indicspecies” (De Cáceres and Legendre 2009), in which larger diet breadth values represent broader diets (Appendix 1). The breadth values estimate the species’ diets but also account for differences in the resources, making them more accurate diet estimates (De Cáceres et al. 2011). Previous research testing diet differences among species calculated breadth using similar methodology (e.g., Sol et al. 2014).
Plumage color may be under thermoregulatory and/or feather degradation resistance pressure (Gloger 1833, Burtt and Ichida 2004; and see Marcondes et al. 2021), so we sampled maximum temperature of the warmest month (BIO5) and annual precipitation (BIO12) climate variables from WorldClim 2 (Fick and Hijmans 2017), based on the species’ range latitude and longitude centroids. We gathered range centroids and maximums from the Birdlife database (http://datazone.birdlife.org/home). Some species were lacking climate data (n = 18), so we used the mean values from congeners. In one case, a species was the sole member of the genus (Nesotriccus ridgwayi), therefore the climate variables reflect the family mean. We also used the species range centroids to assess the species recognition hypothesis (Simpson et al. 2021).
Finally, because there may be a trade-off between plumage coloration and migratory behavior due to the predation pressure, increased sexual selection, or relaxed female social selection (Badyaev and Hill 2003, Friedman et al. 2009, Simpson et al. 2015), we included in our models migration behavior from Tobias et al. (2022). We included migration as a binary variable (non-migratory and migratory) based on the classifications from Tobias et al. (2022). One species (Calyptura cristata) was missing migratory behavior, so we consulted Birds of the World and classified it as non-migratory. Our resulting dataset included 399 species for each sex.
Phylogenetic methods and statistical analysis
To control for non-independence of related species, we downloaded 1000 potential phylogenies from https://birdtree.org (Jetz et al. 2012, 2014; Hackett All Species, Hackett et al. 2008) for all Tyrannidae species. We then used “TreeAnnotator” in BEAST v.1.10.1 (Suchard et al. 2018) to construct a maximum clade credibility tree using 1% burn in and mean node heights (Pageau et al. 2020, Thibault et al. 2022).
All analyses were performed in R 3.5.3 (R Core Team 2017). To assess the evolutionary mechanism of Tyrannidae plumage color, we built Pagel’s lambda (λ) character evolution models with phylogenetic generalized least square (PGLS) statistical models, using the “nlme” package (Pinheiro et al. 2018). To determine which variables, or combination of variables, best explained color variation, we used stepwise model reduction in the “AICcmodavg” package (Mazerolle 2020) based on Akaike information criterion (AIC) and Bayesian information criterion (BIC). First, we constructed a full model, which included either female color score, male color score, or dichromatism as the response variable, and the latitude of the range centroid (absolute value), diet breadth, forest cover, annual precipitation, the maximum temperature of the warmest month, and migration as the predictor variables. Distributions of predictors varied so we scaled each term to a mean of 0 prior to analyses. Separate models were constructed for each sex and dichromatism. Next, we carried out model reduction for all possible models using the “StepAIC” function in the MASS package (Venables and Ripley 2002) and compared candidate models based on AIC and BIC (Burnham and Anderson 2003).
We also used phylogenetic path analyses using the R package “phylopath” (van der Bijl 2018) to assess the direct and indirect effects of the variables from the top PGLS models. We first built candidate path analyses informed by the PGLS models and then ranked models using an information theory approach based on C-statistics (Shipley 2009). Information theory evaluates the conditional independencies of each model and assigns a C statistic. The models are ranked based on the change in C statistic (ΔCICc) between models, where lower C scores are optimized and models within 2 ΔCICc of the top model are competitive and thus also supported. The top phylogenetic path analysis model was then selected as the model with lowest C statistic and models within 2 ΔCICc were subsequently averaged in the final model (for more details please see von Hardenberg and Gonzalez-Voyer 2013). We did not conduct a phylogenetic path analysis in cases where a single predictor variable was identified in our PGLS model selection.
To assess the relationship between color differences and geographic distance, we constructed pairwise distance matrices between all comparisons of species’ range centroids and their color scores. To calculate color distances, we used the “vegdist” function in the Vegan package for R, using Euclidean distances (1 dimension; Oksanen et al. 2013). We then calculated linear geographic distances (km) between all species comparisons from the species’ range centroids from above using the “geodist” package (van Etten 2017). We subsequently conducted a Mantel test, under the hypothesis that color distances would be greater if species ranges were closer in proximity (i.e., trait distance is negatively related to geographic distance). Although Mantel tests may suffer from low statistical power (Harmon and Glor 2010), the debate is ongoing (Hardy and Pavoine 2012). However, it is generally agreed upon that Mantel tests used to test comparative hypotheses violate assumptions of independence (Harmon and Glor 2010, Hardy and Pavoine 2012). We therefore addressed this issue using a phylogenetically controlled Mantel test in the package “evolqg” for R (Melo et al. 2016). We conducted Mantel tests for females, males, and dichromatism at two taxonomic scales: first at the family scale and then at the genus-level as a test assessing the risk of hybridization. For the genus-level analysis, we only included genera with n ≥ 10 species, restricting our analyses to n = 24 genera, and we treated genera as separate hypotheses.
RESULTS
In our dataset, female color score ranged 36.1 to 62.4 (median = 44.9), males ranged 35.1 to 72.1 (median = 45.1), and dichromatism ranged 0 to 30.7 (median = 1.4). Typically, species in our dataset with red, yellow, and black plumage had high plumage scores, gray and rufous species had low scores, and species with a mix of gray, white, and black had intermediate scores (Fig. 1). Structural blue plumage is rare in Tyrannidae species (Gray 1996) and exhibited intermediate scores (e.g., Tachuris rubrigastra female color score = 50.9, male = 56.8).
The final model for female Tyrannidae color (Table 1) included range latitude and annual precipitation (latitude: X²>1396 = 2.93, p = 0.09, ϕ = 0.07 [0-0.18]; annual precipitation: X²1396 = 2.50, p = 0.11, ϕ = 0.06 [0-0.17]; Table 2). Although the PGLS results were not statistically significant, effect size confidence intervals did not overlap 0, suggesting colorful females were from lower latitudes and in areas with less annual precipitation (Fig. 2A and 2B). Male color scores (Table 1) were best predicted by migration behavior (X21397=12.96, p=0.0003, ϕ=0.17 [0.07-0.27]; Table 2), where more colorful males were migratory (Fig. 2C). Tyrannidae dichromatism was best predicted by the reduced model (Table 1), and included latitude (X²1395 = 3.42, p = 0.06, ϕ = 0.08 [0-0.18]), diet breadth (X²1395 = 3.43, p = 0.06, ϕ = 0.08 [0-0.19]), and migration (X²1395 = 7.26, p = 0.007, ϕ = 0.13 [0-0.23], Table 2). Migration was statistically significant, but effect size confidence intervals for latitude and diet breadth did not overlap 0 (Table 2). Species with pronounced dichromatism were found at higher latitudes (Fig. 2D) and had narrower diet breadths (Fig. 2E). Similarly, to male coloration, dichromatic species were more likely to be migratory (Fig. 2F).
Our phylogenetic path analyses revealed direct and indirect effects of ecology and climate on female coloration and dichromatism (Appendix 3). In the top path model for females, annual precipitation had a direct, albeit weak, negative effect on coloration (coef. = -0.06, SE = 0.04) and latitude had a relatively strong, indirect negative effect on precipitation (coef. = -0.49, SE = 0.05, Fig. 3), suggesting more colorful females were found in dry, low latitude areas. The top model for dichromatism included a direct but weakly negative influence of range latitude (coef. = -0.01, SE = 0.03) and diet breadth (coef. = -0.004, SE = 0.03; Fig. 3). Migration positively influenced dichromatism indirectly through latitude (coef. = 0.05, SE = 0.06) and latitude positively influenced diet breadth (coef. = 0.02, SE = 0.03; Fig. 3). We did not conduct a path analysis for male Tyrannidae because our PGLS model identified migration behavior as the sole predictor.
Support for the species recognition hypothesis depended on the taxonomic analysis level (Table 3). From a phylogenetic Mantel test of all Tyrannidae, female color did not show a statistically significant relationship with geographic distance, however confidence intervals did not overlap with 0, suggesting a positive relationship (Table 3; r = 0.02, p = 0.18, CI: 0.01-0.03). Male coloration was positively related to distance (Table 3; r = 0.06, p = 0.02, CI: 0.05-0.07), but dichromatism did not vary with geographic distance (Table 3; r = 0.009, p = 0.39, CI: 0.003-0.02). From genus-level phylogenetic Mantel tests, we found Poecilotriccus female coloration was positively related to geographic distance (Table 3; r = 0.51, p = 0.002, CI: 0.30-0.67). Female Myiarchus coloration was negatively related to distance based on non-zero overlapping confidence intervals, but the result was not statistically significant (Table 3; r = -0.13, p = 0.93, CI: -0.26 - -0.002). Male coloration and geographic distance in Phylloscartes were negatively related (Table 3; r = 0.11, p = 0.04, CI: -0.01-0.23), and positively related in Poecilotriccus (Table 3; r = 0.38, p = 0.01, CI: 0.15-0.57). Male Empidonax coloration showed a positive correlation to geographic distance among congeners from non-zero overlapping CIs, however the result was not statistically significant (Table 3; r = 0.27, p = 0.07, CI: 0.09-0.44). Dichromatism distances were negatively related to geographic proximity in Elaenia because CIs did not overlap 0, but p was > 0.05 (Table 3; r = -0.17, p = 0.92, CI: -0.32 - -0.02). All other relationships between color and geographic distances among congeners were not statistically significant (Table 3).
DISCUSSION
Plumage coloration is an important visual signal that aids in mate choice, predator avoidance, thermoregulation, and species recognition (Bradbury and Vehrencamp 1998). In our study, we simultaneously tested geographic and ecological factors of plumage variation in Tyrannidae, a speciose Passeriformes family, using a large-scale dataset of the Americas. Our phylogenetic comparative analyses revealed variation in plumage coloration among female and male Tyrant flycatchers was associated most strongly with migration behavior and heterospecific differences were consistent with expectations under drift. To a lesser extent, color varied by dietary and climatic factors, both of which may influence how feathers are grown (Hill 2006a). We also incorporated a phylogenetic path analysis to disentangle the direct and indirect effects of this complex and important visual signal.
From our PGLS, latitude and climate correlated with female plumage and dichromatism, but the results were not statistically significant. This contrasts with previous findings of pronounced dichromatism at higher latitudes (Friedman et al. 2009, Simpson et al. 2015), suggesting the effect varies among families. Our results could be driven by the broad variation of dichromatism in Tyrannidae (color score range = 0-30.7). For example, the most dichromatic species in our dataset, Vermillion Flycatcher (Pyrocephalus obscurus, color score = 30.7), is found at relatively low latitudes in North and Central America. Conversely, Great-shrike Tyrant (Agriornis lividus) is more monotypic (color score = 0.7) and occurs at relatively high latitudes in Chile and Argentina. Similarly, neotropical migrants such as Empidonax, Contopus, Myiarchus, and Tyrannus flycatchers can occur at higher latitudes and are relatively monotypic. Although studies testing latitudinal effects on morphology typically use range centroids (Simpson et al. 2015, Tobias et al. 2022), maximum range latitude may reveal patterns in Tyrannidae. Indeed, a univariate test of maximum range latitude suggests a negative effect of maximum latitude in females, but no relationship in males (females: estimate = -0.07, X²1397 = 4.89, p = 0.02; males: estimate = 0.003, X²1397 = 0.006, p = 0.94). Female color is consistent with historical evidence of latitudinal gradients (Gloger 1833, Görnitz 1923), suggesting colorful plumage is more prevalent in low latitude tropical areas (i.e., “Simple Gloger’s rule,” sensu Rensch 1929), however the direct and indirect paths of climate and latitude identified from our path analyses support the hypothesis that latitudinal effects on plumage coloration is more complex than previously thought (i.e., “Complex Gloger’s rule,” sensu Rensch 1929, Delhey 2019, Marcondes et al. 2021). In our path analysis for females, we identified a direct effect of annual precipitation. Species in wetter climes may be under pressure to express bacteria- or abrasion-resistant melanin-based plumage, which are both more prevalent in mesic climates (Burtt 1986, Burtt and Ichida 2004). Dichromatism was also influenced by latitude in the path analysis, but the coefficient was relatively weak, and the stronger latitudinal influence was indirect through diet. Precipitation may mediate resource availability (Williams 1951, 1961) and can limit foraging efficiency and/or invertebrate activity (Williams 1951, Taylor 1963, Grüebler et al. 2008), thereby reducing nutrient intake required for coloration (Laczi et al. 2020). Although we did not explicitly test the “Complex Gloger’s rule” (sensu Rensch 1929), Tyrannidae exhibit eumelanin and phaeomelanin plumage and species occur across broad climatic gradients, so the group may prove useful in evaluating the generalizability of the so-called “Complex Gloger’s rule” (Rensch 1929, Delhey 2019).
Consistent with the hypothesis that migratory behavior influences plumage coloration, we found pronounced Tyrannidae male color and dichromatism in migratory species. In other avian taxa, the association between migration and sexual dichromatism results from a loss of elaboration in females (Friedman et al. 2009, Simpson et al. 2015) and there are several non-mutually exclusive hypotheses to explain this finding. First, pronounced dichromatism may arise from losses of female plumage elaboration for crypsis during migration (Caldwell 1986, Slagsvold et al. 1995, Huhta et al. 2003). Second, relaxed social selection for females at higher latitudes may reduce territoriality (West-Eberhard 1983, Irwin 1994, Tobias et al. 2011). Third, enhanced migratory male coloration may result from increased sexual selection at breeding areas (Badyaev and Hill 2003). And finally, recent evidence suggests dichromatism evolved for conspecific recognition (see Luro and Hauber 2022). Given female Tyrannidae coloration did not significantly vary with migration behavior, our results are consistent with the notion of pronounced dichromatism in migratory species due to the loss of female elaboration. To better understand the evolution of dichromatism in Tyrannidae, future studies could conduct ancestral state reconstructions to confirm whether dichromatism results from a loss or gain of elaboration and whether this pattern is associated with gains of migratory behavior (Badyaev and Hill 2003).
We did not find support for the light environment affecting tyrannid color evolution because no final models included forest cover. Similarly in Part I of our study in which we investigated the evolution of Tyrannidae song, our models failed to find support for acoustic adaptation (using forest cover), and song variation was mostly associated with morphology (body and bill size; Schoen et al. 2023). Other taxa are sensitive to light environments, such as Phylloscopus warblers, in which an individual’s plumage contrasts with the light environment (Marchetti 1993). Recent comparative studies indicate an interaction between light environments and climate affects plumage coloration (Gomez and Théry 2004, Marcondes et al. 2021) and indeed our models revealed plumage coloration was influenced by climate. We did however find positive effects of forest cover on carotenoid-based plumage coloration using chromaticity as a proxy (see Appendix 2; Dey et al. 2014), perhaps due to greater variation in carotenoid resources in forests (Grether et al. 2001, Jones et al. 2010), but spectrometry is needed to confirm if carotenoid signals are indeed associated with forest cover.
The species recognition hypothesis predicts differences among species signaling has evolved for conspecific recognition (Sætre et al. 1997, Seddon 2005). We conducted an analysis that included all Tyrannidae and found female color and dichromatism was not significantly related to geographic proximity, however confidence intervals suggested a weakly positive effect. Males were significantly positive, but the effect was also weak. The positive effects of geographic distance may be evidence of drift (Hill 2006b), and given that many flycatcher species are habitat specialists, heterospecific color differences may have arisen from geographic isolation and/or natural selection (e.g., Johnson and Cicero 2002). In contrast, Tyrannidae song (Schoen et al. 2023) at the family scale may have evolved for species recognition or to minimize signal overlap (Liou and Price 1994, Seddon 2005, Simpson et al. 2021). Genus-level tests of color differences offered some evidence for species recognition because female Myiarchus color and Elaenia dichromatism were negatively related to geographic proximity. Interestingly, Myiarchus song from Part I of our study also showed a negative relationship to congener proximity, suggesting correlated evolution of song and plumage and multi-modal signaling in species recognition (Schoen et al. 2023). Other genera showed positive relationships between color and congener proximity including Empidonax male, Phylloscartes male, and Poecilotriccus female and male coloration. Empidonax flycatchers are typically thought to be a cryptic group (e.g., Prescott 1987) and congener song differences did not vary with geographic proximity (Schoen et al. 2023), but our color results suggest differences arose from genetic drift (Hill 2006b). Poecilotriccus congener plumage differences also varied positively, as did their song difference (Schoen et al. 2023), offering evidence of similar evolutionary trajectories of song and plumage in Tyrannidae. The variable patterns of plumage and song among Tyrannidae genera suggest multiple signal modalities, possibly including other aspects of morphology, are needed to recognize conspecifics (Seddon 2005, Kirschel et al. 2009, Uy et al. 2009).
Overall, our large-scale phylogenetic analysis examines factors influencing Tyrant flycatcher plumage coloration evolution, focusing on the underlying processes of radiation in the family. We identified migration behavior, climate, and diet as the main predictors of plumage coloration and did not find strong support for a latitudinal gradient or for light environment effects (although see Appendix 2). Family-level differences in color were consistent with expectations under drift, which contrasts our findings from Part I of our study, however some genera showed support for the species recognition hypothesis, and we posit species delineations are maintained through multimodal signaling including song and other aspects of morphology. To better understand the mechanisms driving dichromatism in Tyrannidae, future studies could investigate whether dichromatism results from a loss or gain of plumage elaboration due to migratory behavior, a pattern observed in Icteridae orioles and Parulidae warblers (Friedman et al. 2009, Simpson et al. 2015), as well as formally testing the relationship between Tyrant flycatcher color and song.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Equal contributions for first author: SM and SMM. SMM conceived of the project. SMM and MWR curated data. SM and SMM conducted statistical analyses. SM wrote initial draft and SMM wrote final draft. MWR and JS assisted with writing and analyses. All authors contributed to editing the manuscript and gave final approval for the manuscript.
ACKNOWLEDGMENTS
This research was completed on the traditional lands of the Tk’emlúps te Secwépemc within Secwépemc’ulucw, the traditional and unceded territory of the Secwépemc. Funding for this project was provided by a Natural Sciences and Engineering Research Council of Canada Discovery Grant to M. W. R. We thank J. Dale for sharing color data and to iNaturalist users for the photos in Fig. 1 (iNaturalist user miscbr: Rubinus pyrocephalus Photo 215330562; Issac Rath: Sayornis nigricans Photo 97140304; Dario Sanches: Pitangus sulphuratus Photo 1766279; Frank Fogarty: Empidonax oberholseri Photo 80317978; Michael Friedman: Empidonax traillii Photo 217464498; iNaturalist milkocj: Empidonax minimus Photo 216450828; iNaturalist user avocat: Knipolegus aterrimus Photo 232272065; Ricardo Gagliardi: Arundinicola leucocephalaPhoto 206267161; Luis Trinchan: Tyrannus forficatus Photo 57471666). Comments from M. Hauber and two anonymous reviewers greatly improved the manuscript.
DATA AVAILABILITY
Data used in this manuscript are available as appendices.
LITERATURE CITED
Aidala, Z., L. Huynen, P. L. Brennan, J. Musser, A. Fidler, N. Chong, G. E. Machovsky Capuska, M. G. Anderson, A. Talaba, D. Lambert, and M. E. Hauber. 2012. Ultraviolet visual sensitivity in three avian lineages: paleognaths, parrots, and passerines. Journal of Comparative Physiology A 198:495-510. https://doi.org/10.1007/s00359-012-0724-3
Andersson, M. 1994. Sexual selection. Princeton University Press, Princeton, New Jersey, USA.
Badyaev, A. V., and G. E. Hill. 2003. Avian sexual dichromatism in relation to phylogeny and ecology. Annual Review of Ecology, Evolution, and Systematics 34:27-49. https://doi.org/10.1146/annurev.ecolsys.34.011802.132441
Bailey, S. F. 1978. Latitudinal gradients in colors and patterns of passerine birds. Condor 80:372-381. https://doi.org/10.2307/1367187
Baston, D. 2020. exactextractr: fast extraction from raster datasets using polygons. R package version 0.5.0. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/exactextractr/index.html
Boughman, J. W. 2002. How sensory drive can promote speciation. Trends in Ecology and Evolution 17:571-577. https://doi.org/10.1016/S0169-5347(02)02595-8
Bradbury, J. W., and S. L. Vehrencamp. 1998. Principles of animal communication. Second edition. Sinauer Associates, Sunderland, Massachusetts, USA.
Buchhorn, M., M. Lesiv, N.-E. Tsendbazar, M. Herold, L. Bertels, and B. Smets. 2020. Copernicus global land cover layers — collection 2. Remote Sensing 12:1044. https://doi.org/10.3390/rs12061044
Burnham, K. P., and D. R. Anderson. 2003. Model selection and multimodel inference: a practical information-theoretic approach. Springer, Berlin, Germany. https://doi.org/10.1007/b97636
Burtt, E. H. 1986. An analysis of physical, physiological, and optical aspects of avian coloration with emphasis on wood-warblers. Ornithological Monographs 38:1-126.
Burtt, E. H., and J. M. Ichida. 2004. Gloger’s rule, feather degrading bacteria, and color variation among song sparrows. Condor 106:681-686. https://doi.org/10.1093/condor/106.3.681
Caldwell, G. S. 1986. Predation as a selective force on foraging herons: effects of plumage color and flocking. Auk 103:494-505.
Chui, C. K. S., and S. M. Doucet. 2009. A test of ecological and sexual selection hypotheses for geographical variation in coloration and morphology of Golden-crowned Kinglets (Regulus satrapa). Journal of Biogeography 36:1945-1957. https://doi.org/10.1111/j.1365-2699.2009.02132.x
Cooney, C. R., J. A. Bright, E. J. R. Capp, A. M. Chira, E. C. Hughes, C. J. A. Moody, L. O. Nouri, Z. K. Varley, and G. H. Thomas. 2017. Mega-evolutionary dynamics of the adaptive radiation of birds. Nature 542:344-347. https://doi.org/10.1038/nature21074
Dale, J., C. J. Dey, K. Delhey, B. Kampenaers, and M. Valcu. 2015. The effects of life history and sexual selection on male and female plumage coloration. Nature 527:367-370. https://doi.org/10.1038/nature15509
De Cáceres, M., and P. Legendre. 2009. Associations between species and groups of sites: indices and statistical inference. Ecology 90:3566-3574. https://doi.org/10.1890/08-1823.1
De Cáceres, M., D. Sol, O. Lapiedra, and P. Legendre. 2011. A framework for estimating niche metrics using the resemblance between qualitative resources. Oikos 120:1341-1350. https://doi.org/10.1111/j.1600-0706.2011.19679.x
Delhey, K. 2019. A review of Gloger’s rule, an ecogeographical rule of colour: definitions, interpretations and evidence. Biological Reviews 94:1294-1316. https://doi.org/10.1111/brv.12503
Dey, C. J., M. Valcu, B. Kempenaers, and J. Dale. 2014. Carotenoid‐based bill coloration functions as a social, not sexual, signal in songbirds (Aves: Passeriformes). Journal of Evolutionary Biology 28:250-258. https://doi.org/10.1111/jeb.12560
Eaton, M. D., and S. M. Lanyon. 2003. The ubiquity of avian ultraviolet plumage reflectance. Proceedings of the Royal Society of London Series B: Biological Sciences 270:1721-1726. https://doi.org/10.1098/rspb.2003.2431
Endler, J. A. 1992. Signals, signal conditions, and the direction of evolution. American Naturalist 139:S125-S153. https://doi.org/10.1086/285308
Endler, J. A. 1993. The color of light in forests and its implications. Ecological Monographs 63:1-27. https://doi.org/10.2307/2937121
Endler, J. A., and M. Thery. 1996. Interacting effects of LEK placement, display behavior, ambient light, and color patterns in three neotropical forest-dwelling birds. American Naturalist 148:421-452. https://doi.org/10.1086/285934
Fick, S. E., and R. J. Hijmans. 2017. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology 37:4302-4315. https://doi.org/10.1002/joc.5086
Friedman, N. R., C. M. Hofmann, B. Kondo, and K. E. Omland. 2009. Correlated evolution of migration and sexual dichromatism in the New World Orioles (Icterus). Evolution 63:3269-3274. https://doi.org/10.1111/j.1558-5646.2009.00792.x
Gloger, C. W. L. 1833. Das Abändern der Vögel durch Einfluss des Klimas. August Schulz, Breslau, Germany.
Gomez, D., and M. Théry. 2004. Influence of ambient light on the evolution of color signals: comparative analysis of a Neotropical rainforest bird community. Ecology Letters 7:279-284. https://doi.org/10.1111/j.1461-0248.2004.00584.x
Görnitz, K. 1923. Ueber die Wirkung klimatischer Faktoren auf die Pigmentfarben der Vogelfedern. Journal Für Ornithologie 71:456-511. https://doi.org/10.1007/BF01975089
Gray, D. A. 1996. Carotenoids and sexual dichromatism in North American passerine birds. American Naturalist 148:453-480. https://doi.org/10.1086/285935
Grether, G. F., D. F. Millie, M. J. Bryant, D. N. Reznick, and W. Mayea. 2001. Rain forest canopy cover, resource availability, and life history evolution in guppies. Ecology 82:1546-1559. https://doi.org/10.1890/0012-9658(2001)082[1546:RFCCRA]2.0.CO;2
Grüebler, M. U., M. Morand, and B. Naef-Daenzer. 2008. A predictive model of the density of airborne insects in agricultural environments. Agriculture, Ecosystems and Environment 123:75-80. https://doi.org/10.1016/j.agee.2007.05.001
Hackett, S. J., R. T. Kimball, S. Reddy, R. C. K. Bowie, E. L. Braun, M. J. Braun, J. L. Chojnowski, W. A. Cox, K.-L. Han, J. Harshman, C. J. Huddleston, B. D. Marks, K. J. Miglia, W. S. Moore, F. H. Sheldon, D. W. Steadman, C. C. Witt, and T. Yuri. 2008. A phylogenomic study of birds reveals their evolutionary history. Science 320:1763–1768. https://doi.org/10.1126/science.1157704
Hardy, O. J., and S. Pavoine. 2012. Assessing phylogenetic signal with measurement error: a comparison of mantel tests, Blomberg et al.’s K, and phylogenetic distograms. Evolution 66:2614-2621. https://doi.org/10.1111/j.1558-5646.2012.01623.x
Harmon, L. J., and R. E. Glor. 2010. Poor statistical performance of the Mantel test in phylogenetic comparative analyses. Evolution 64:2173-2178. https://doi.org/10.1111/j.1558-5646.2010.00973.x
Hill, G. E. 2006. Environmental regulation of ornamental coloration. Pages 507-560 in G. E. Hill and K. J. McGraw, editors. Bird coloration, Vol. 1. Mechanisms and measurements. Harvard University Press, Cambridge, Massachusetts, USA. https://doi.org/10.2307/j.ctv22jnscm.15
Hill, G. E. 2006. Female mate choice for ornamental coloration. Pages 177-242 in G. E. Hill and K. J. McGraw, editors. Bird coloration. Vol 2. Function and evolution. Harvard University Press, Cambridge, Massachusetts, USA. https://doi.org/10.4159/9780674273818-005
Hill, G. E. 2015. Sexiness, individual condition, and species identity: the information signaled by ornaments and assessed by choosing females. Evolutionary Biology 42:251-259. https://doi.org/10.1007/s11692-015-9331-x
Huhta, E., S. Rytkönen, and T. Solonen. 2003. Plumage brightness of prey increases predation risk: an among‐species comparison. Ecology 84:1793-1799. https://doi.org/10.1890/0012-9658(2003)084[1793:PBOPIP]2.0.CO;2
Irwin, R. E. 1994. The evolution of plumage dichromatism in the New World blackbirds: social selection on female brightness. American Naturalist 144890-907. https://doi.org/10.1086/285717
Jetz, W., G. H. Thomas, J. B. Joy, K. Hartmann, and A. O. Mooers. 2012. The global diversity of birds in space and time. Nature 491: 444-448. https://doi.org/10.1038/nature11631
Jetz, W., G. H. Thomas, J. B. Joy, D. W. Redding, K. Hartmann, and A. O. Mooers. 2014. Global distribution and conservation of evolutionary distinctness in birds. Current Biology 24:919-930. https://doi.org/10.1016/j.cub.2014.03.011
Johnson, N. K., and C. Cicero. 2002. The role of ecologic diversification in sibling speciation of Empidonax flycatchers (Tyrannidae): multigene evidence from mtDNA. Molecular Ecology 11:2065-2081. https://doi.org/10.1046/j.1365-294X.2002.01588.x
Jones, T. M., A. D. Rodewald, and D. P. Shustack. 2010. Variation in plumage coloration of Northern Cardinals in urbanizing landscapes. Wilson Journal of Ornithology 122:326-333. https://doi.org/10.1676/09-082.1
Kirschel, A. N. G., D. T. Blumstein, and T. B. Smith. 2009. Character displacement of song and morphology in African Tinkerbirds. Proceedings of the National Academy of Sciences 106:8256-8261. https://doi.org/10.1073/pnas.0810124106
Laczi, M., G. Hegyi, G. Nagy, R. Pongrácz, and J. Tìrìk. 2020. Yellow plumage colour of Great Tits Parus major correlates with changing temperature and precipitation. Ibis 162:232-237. https://doi.org/10.1111/ibi.12761
Lande, R. 1981. Models of speciation by sexual selection on polygenic traits. Proceedings of the National Academy of Sciences USA 78:3721-3725. https://doi.org/10.1073/pnas.78.6.3721
Liou, L. W., and T. D. Price. 1994. Speciation by reinforcement of premating isolation. Evolution 48:1451-1459. https://doi.org/10.1111/j.1558-5646.1994.tb02187.x
Luro, A. B., and M. E. Hauber. 2022. Pressure for rapid and accurate mate recognition promotes avian‐perceived plumage sexual dichromatism in true thrushes (genus: Turdus). Journal of Evolutionary Biology 35:1558-1567. https://doi.org/10.1111/jeb.14089
Mahoney, S. M., M. D. Oud, C. Pageau, M. A. de Menezes, N. Smith, J. V. Briskie, and M. W. Reudink. Global patterns of plumage colour evolution in island-living passeriform birds. In review.
Marchetti, K. 1993. Dark habitats and bright birds illustrate the role of the environment in species divergence. Nature 362:149-152. https://doi.org/10.1038/362149a0
Marcondes, R. S., J. A. Nations, G. F. Seeholzer, and R. T. Brumfield. 2021. Rethinking Gloger’s rule: climate, light environments, and color in a large family of tropical birds (Furnariidae). American Naturalist 197:592-606. https://doi.org/10.1086/713386
Mazerolle, M. J. 2020. Model selection and multimodel inference using the AICcmodavg package. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/AICcmodavg/vignettes/AICcmodavg.pdf
McGraw, K. J., and G. E. Hill. 2001. Carotenoid access and intraspecific variation in plumage pigmentation in male American Goldfinches (Carduelis tristis) and Northern Cardinals (Cardinalis cardinalis). Functional Ecology 15:732-739. https://doi.org/10.1046/j.0269-8463.2001.00574.x
McNaught, M. K., and I. P. F. Owens. 2002. Interspecific variation in plumage colour among birds: species recognition or light environment? Journal of Evolutionary Biology 15:505-514. https://doi.org/10.1046/j.1420-9101.2002.00431.x
Melo, D., G. Garcia, A. Hubbe, A. P. Assis, and G. Marroig. 2016. EvolQG - An R package for evolutionary quantitative genetics. F1000Research 4:925. https://doi.org/10.12688/f1000research.7082.2
Mikula, P., M. Valcu, H. Brumm, M. Bulla, W. Forstmeier, T. Petrusková, B. Kempenaers, and T. Albrecht. 2020. A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. Ecology Letters 24:477-486. https://doi.org/10.1111/ele.13662
Nuñez, M. A., M. C. Chiuffo, A. Pauchard, and R. D. Zenni. 2021. Making ecology really global. Trends in Ecology and Evolution 36:766-769. https://doi.org/10.1016/j.tree.2021.06.004
Ödeen, A., and O. Håstad. 2003. Complex distribution of avian color vision systems revealed by sequencing the SWS1 opsin from total DNA. Molecular Biology and Evolution 20:855-861. https://doi.org/10.1093/molbev/msg108
Ödeen, A., S. Pruett-Jones, A. C. Driskell, J. K. Armenta, and O. Håstad. 2012. Multiple shifts between violet and ultraviolet vision in a family of passerine birds with associated changes in plumage coloration. Proceedings of the Royal Society B: Biological Sciences 279:1269-1276. https://doi.org/10.1098/rspb.2011.1777
Oksanen, J., F. G. Blanchet, R. Kindt, P. Legendre, P. R. Minchin, R. B. O’hara, G. L. Simpson, and P. Solymos, M. H. H. Stevens, H. Wagner, and M. J. Oksanen. 2013. Package ‘vegan’: community ecology package. Version 2. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/vegan/vegan.pdf
Pageau, C., C. M. Tonra, M. Shaikh, N. J. Flood, and M. W. Reudink. 2020. Evolution of moult-migration is directly linked to aridity of the breeding grounds in North American passerines. Biology Letters 16:20200155. https://doi.org/10.1098/rsbl.2020.0155
Pinheiro, J., D. Bates, S. DebRoy, and D. Sarkar. 2018. nlme: linear and nonlinear mixed effects models. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/nlme/nlme.pdf
Prescott, D. R. C. 1987. Territorial responses to song playback in allopatric and sympatric populations of alder Empidonax alnorum and willow E. traillii flycatchers. Wilson Bulletin 99611-619.
Prum, R. O. 1990. Phylogenetic analysis of the evolution of display behavior in the Neotropical manakins (Aves: Pipridae). Ethology 84:202-231. https://doi.org/10.1111/j.1439-0310.1990.tb00798.x
Prum, R. O. 2006. Anatomy, physics, and evolution of structural colors. Pages 243-294 in G. E. Hill and K. J. McGraw, editors. Bird coloration. Vol 1. Mechanisms and measurements. Harvard University Press, Cambridge, Massachusetts, USA. https://doi.org/10.2307/j.ctv22jnscm.10
Prum, R. O. 2010. The Lande-Kirkpatrick mechanism is the null model of evolution by intersexual selection: implications for meaning, honesty, and design in intersexual signals. Evolution: International Journal of Organic Evolution 64:3085-3100. https://doi.org/10.1111/j.1558-5646.2010.01054.x
R Core Team. 2017. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rensch, B. 1929. Prinzip geographischer rassenkreise und das problem der artbildung. Borntraeger, Berlin, Germany.
Reudink, M. W., A. E. McKellar, K. L. D. Marini, S. L. McArthur, P. P. Marra, and L. M. Ratcliffe. 2015. Inter-annual variation in American redstart (Setophaga ruticilla) plumage color is associated with rainfall and temperature during moult: an 11-year study. Oecologia 178:161-173. https://doi.org/10.1007/s00442-014-3167-4
Riebel, K., K. J. Odom, N. E. Langmore, and M. L. Hall. 2019. New insights from female bird song: towards an integrated approach to studying male and female communication roles. Biology Letters 15:20190059. https://doi.org/10.1098/rsbl.2019.0059
Sætre, G. P., T. Moum, S. Bureš, M. Král, M. Adamjan, and J. Moreno. 1997. A sexually selected character displacement in flycatchers reinforces premating isolation. Nature 387:589-592. https://doi.org/10.1038/42451
Sayol, F., P. A. Downing, A. N. Iwaniuk, J. Maspons, and D. Sol. 2018. Predictable evolution towards larger brains in birds colonizing oceanic islands. Nature Communications 9:2820. https://doi.org/10.1038/s41467-018-05280-8
Schoen, J. E., S. M. Miller, M. W. Reudink, Q. Jennings, and S. M. Mahoney. 2023. Disentangling the mechanisms of signal evolution in Tyrannidae flycatchers, part I: song is constrained by morphology and covaries with ecological factors. Journal of Field Ornithology 94(3):6. https://doi.org/10.5751/JFO-00260-940306
Seddon, N. 2005. Ecological adaptation and species recognition drives vocal evolution in neotropical suboscine birds. Evolution 59:200-215. https://doi.org/10.1111/j.0014-3820.2005.tb00906.x
Seddon, N., C. A. Botero, J. A. Tobias, P. O. Dunn, H. E. A. MacGregor, D. R. Rubenstein, J. A. C. Uy, J. T. Weir, L. A. Whittingham, and R. J. Safran. 2013. Sexual selection accelerates signal evolution during speciation in birds. Proceedings of the Royal Society of London B: Biological Sciences 280:20131065. https://doi.org/10.1098/rspb.2013.1065
Seddon, N., J. A. Tobias, M. Eaton, and A. Ödeen. 2010. Human vision can provide a valid proxy for avian perception of sexual dichromatism. Auk 127:283-292. https://doi.org/10.1525/auk.2009.09070
Shipley, B. 2009. Confirmatory path analysis in a generalized multilevel context. Ecology 90:363-368. https://doi.org/10.1890/08-1034.1
Simpson, R. K., M. A. Johnson, and T. G. Murphy. 2015. Migration and the evolution of sexual dichromatism: evolutionary loss of female coloration with migration among wood-warblers. Proceedings of the Royal Society of London B: Biological Sciences 282:20150375. https://doi.org/10.1098/rspb.2015.0375
Simpson, R. K., D. R. Wilson, A. F. Mistakidis, D. J. Mennill, and S. M. Doucet. 2021. Sympatry drives color and song evolution in wood-warblers (Parulidae). Proceedings of the Royal Society of London B: Biological Sciences 288:20202804. https://doi.org//10.1098/rspb.2020.2804
Slagsvold, T., S. Dale, and A. Kruszewicz. 1995. Predation favours cryptic coloration in breeding male Pied Flycatchers. Animal Behavior 50:1109-1121. https://doi.org/10.1016/0003-3472(95)80110-3
Sol, D., C. González‐Lagos, D. Moreira, J. Maspons, and O. Lapiedra. 2014. Urbanisation tolerance and the loss of avian diversity. Ecology Letters 17:942-950. https://doi.org/10.1111/ele.12297
Suchard, M. A., P. Lemey, G. Baele, D. L. Ayres, A. J. Drummond, and A. Rambaut. 2018. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evolution 4:vey016. https://doi.org/10.1093/ve/vey016
Taylor, L. R. 1963. Analysis of the effect of temperature on insects in flight. Journal of Animal Ecolology 32:99-117. https://doi.org/10.2307/2520
Thibault, E., S. M. Mahoney, J. V. Briskie, M. Shaikh, and M. W. Reudink. 2022. Extra-pair paternity drives plumage colour elaboration in male passerines. PLoS One 17:e0273347. https://doi.org/10.1371/journal.pone.0273347
Tobias, J. A., V. Gamarra‐Toledo, D. García‐Olaechea, P. C. Pulgarín, and N. Seddon. 2011. Year‐round resource defence and the evolution of male and female song in suboscine birds: social armaments are mutual ornaments. Journal of Evolutionary Biology 24:2118-2138. https://doi.org/10.1111/j.1420-9101.2011.02345.x
Tobias, J. A., C. Sheard, A. L. Pigot, A. J. Devenish, J. Yang, F. Sayol, M. H. C. Neate-Clegg, N. Alioravainen, T. L. Weeks, R. A. Barber, P. A. Walkden, H. E. A. MacGregor, S. E. I. Jones, C. Vincent, A. G. Phillips, N. M. Marples, F. A. Montaño-Centellas, V. Leandro-Silva, S. Claramunt, B. Darski, B. G. Freeman, T. P. Bregman, C. R. Cooney, E. C. Hughes, E. J. R. Capp, Z. K. Varley, N. R. Friedman, H. Korntheuer, A. Corrales-Vargas, C. H. Trisos, B. C. Weeks, D. M. Hanz, T. Töpfer, G. A. Bravo, V. Remeš, L. Nowak, L. S. Carneiro, A. J. Moncada R., B. Matysioková, D. T. Baldassarre, A. Martínez-Salinas, J. D. Wolfe, P. M. Chapman, B. G. Daly, M. C. Sorensen, A. Neu, M. A. Ford, R. J. Mayhew, L. F. Silveira, D. J. Kelly, N. N. D. Annorbah, H. S. Pollock, A. M. Grabowska-Zhang, J. P. McEntee, J. C. T. Gonzalez, C. G. Meneses, M. C. Muñoz, L. L. Powell, G. A. Jamie, T. J. Matthews, O. Johnson, G. R. R. Brito, K. Zyskowski, R. Crates, M. G. Harvey, M. J. Zevallos, P. A. Hosner, T. Bradfer-Lawrence, J. M. Maley, F. G. Stiles, H. S. Lima, K. L. Provost, M. Chibesa, M. Mashao, J. T. Howard, E. Mlamba, M. A. H. Chua, B. Li, M. I. Gómez, N. C. García, M. Päckert, J. Fuchs, J. R. Ali, E. P. Derryberry, M. L. Carlson, R. C. Urriza, K. E. Brzeski, D. M. Prawiradilaga, M. J. Rayner, E. T. Miller, R. C. K. Bowie, R.-M. Lafontaine, R. P. Scofield, Y. Lou, L. Somarathna, D. Lepage, M. Illif, E. L. Neuschulz, M. Templin, D. M. Dehling, J. C. Cooper, O. S. G. Pauwels, K. Analuddin, J. Fjeldså, N. Seddon, P. R. Sweet, F. A. J. DeClerck, L. N. Naka, J. D. Brawn, A. Aleixo, K. Böhning-Gaese, C. Rahbek, S. A. Fritz, G. H. Thomas, and M. Schleuning. 2022. AVONET: morphological, ecological and geographical data for all birds. Ecology Letters 25:581-597. https://doi.org/10.1111/ele.13898
Tobias, M. C., and G. E. Hill. 1998. A test of sensory bias for long tails in the house finch. Animal Behaviour 56:71-78. https://doi.org/10.1006/anbe.1998.0740
Uy, J. A. C., R. G. Moyle, and C. E. Filardi. 2009. Plumage and song differences mediate species recognition between incipient flycatcher species of the Solomon Islands. Evolution 63:153-164. https://doi.org/10.1111/j.1558-5646.2008.00530.x
van der Bijl, W. 2018. phylopath: easy phylogenetic path analysis in R. PeerJ 6:e4718. https://doi.org/10.7717/peerj.4718
van Etten, J. 2017. R package gdistance: distances and routes on geographical grids. Journal of Statistical Software 76:1-21. https://doi.org/10.18637/jss.v076.i13
Venables, W. N., and B. D. Ripley. 2002. Modern applied statistics with S. Fourth Edition. Springer, New York, New York, USA. https://doi.org/10.1007/978-0-387-21706-2
von Hardenberg, A., and A. Gonzalez‐Voyer. 2013. Disentangling evolutionary cause‐effect relationships with phylogenetic confirmatory path analysis. Evolution 67:378-387. https://doi.org/10.1111/j.1558-5646.2012.01790.x
Ward, G. M., S. M. Mahoney, S. Joly, and M. W. Reudink. 2021. Effects of age and weather during moult on mountain bluebird Sialia currucoides structural coloration. Journal of Avian Biology 52:e02616. https://doi.org/10.1111/jav.02616
West-Eberhard, M. J. 1983. Sexual selection, social competition, and speciation. Quarterly Review of Biology 58:155-183. https://doi.org/10.1086/413215
White, T. E. 2020. Structural colours reflect individual quality: a meta-analysis. Biology Letters 16:20200001. https://doi.org/10.1098/rsbl.2020.0001
Williams, C. B. 1951. Changes in insect populations in the field in relation to preceding weather conditions. Proceedings of the Royal Society B 138:130-156. https://doi.org/10.1098/rspb.1951.0011
Wilman, H., J. Belmaker, J. Simpson, C. de la Rosa, M. M. Rivadeneira, and W. Jetz. 2014. EltonTraits 1.0: species‐level foraging attributes of the world’s birds and mammals. Ecological Archives E095‐178. Ecology 95:2027. https://doi.org/10.1890/13-1917.1
Wolf, B. O., and G. E. Walsberg. 2000. The role of plumage in heat transfer processes of birds. American Zoologist 40:575-584. https://doi.org/10.1093/icb/40.4.575
Zink, R. M., and J. V. Remsen. 1986. Evolutionary patterns and processes of geographic variation in birds. Current Ornithology 4:1-69.
Fig. 1
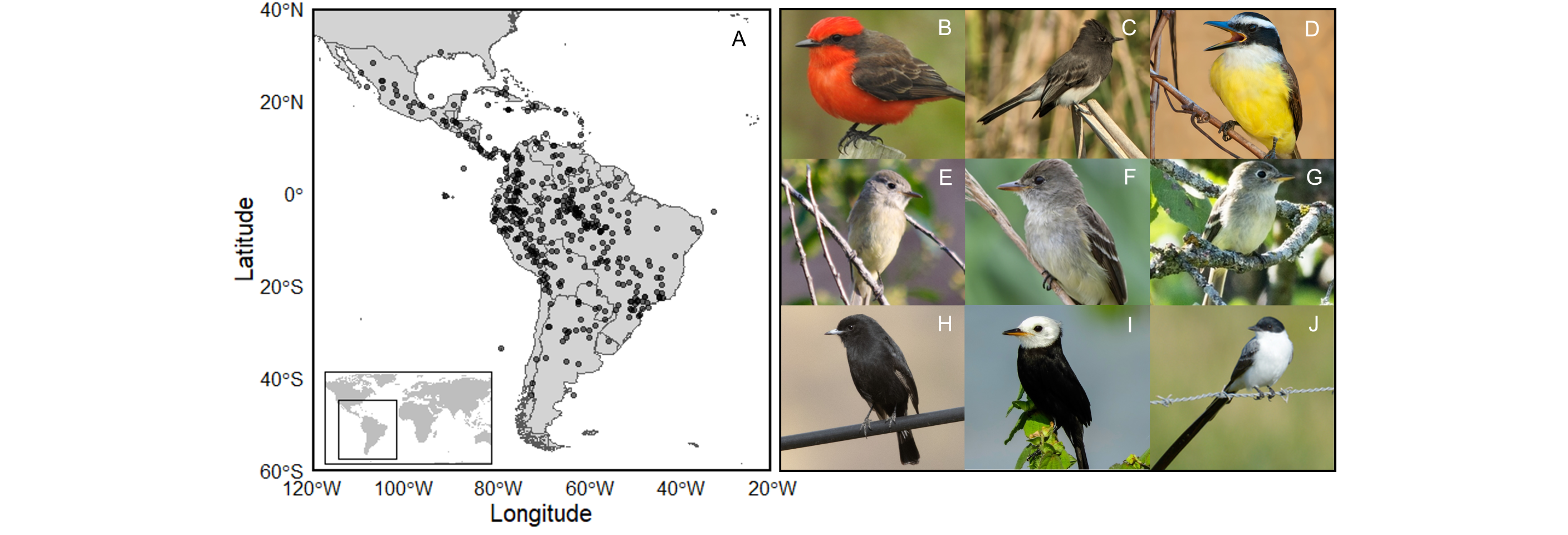
Fig. 1. A. Tyrannidae species occur across broad latitudinal, climatic, and ecological gradients (range centroids are shown by black dots). Tyrant flycatchers are a useful group to investigate plumage color evolution because although heterospecific coloration can be variable with species expressing carotenoid- and melanin-based pigments (B-D), coloration among some congeners is highly conserved (E-G). To assess color variation, we used published color scores from Dale et al. (2015) in which Tyrannidae ranged 35.1-72.1. Photos B and D represent higher scores (Pyrocephalus rubinus, 72.1, Pitangus sulphuratus, 59.0). Photos E-G exhibit low color scores (range 38-42.5). Species with primarily black coloration have higher color scores (H. Knipolegus aterrimus, 64.7; I. Arundinicola leucocephala, 63.4), and species with a combination of whites, blacks, and grays are intermediate (C. Sayornis nigricans, 55.8, J. Tyrannus forficatus, 50.4). Photo credits: B. P. rubinus (iNaturalist user miscbr), C. S. nigricans (Isaac Rath), D. P. sulphuratus (Dario Sanches), E. Empidonax oberholseri (Frank Fogarty), F. E. traillii (Michael Friedman), G. E. minimus (iNaturalist user milkocj), H. K. aterrimus (iNaturalist user avocat), I. A. leucocephala (Ricardo Gagliardi), J. T. forficatus (Luis Trinchan). Photo numbers can be found in acknowledgments.
Fig. 2
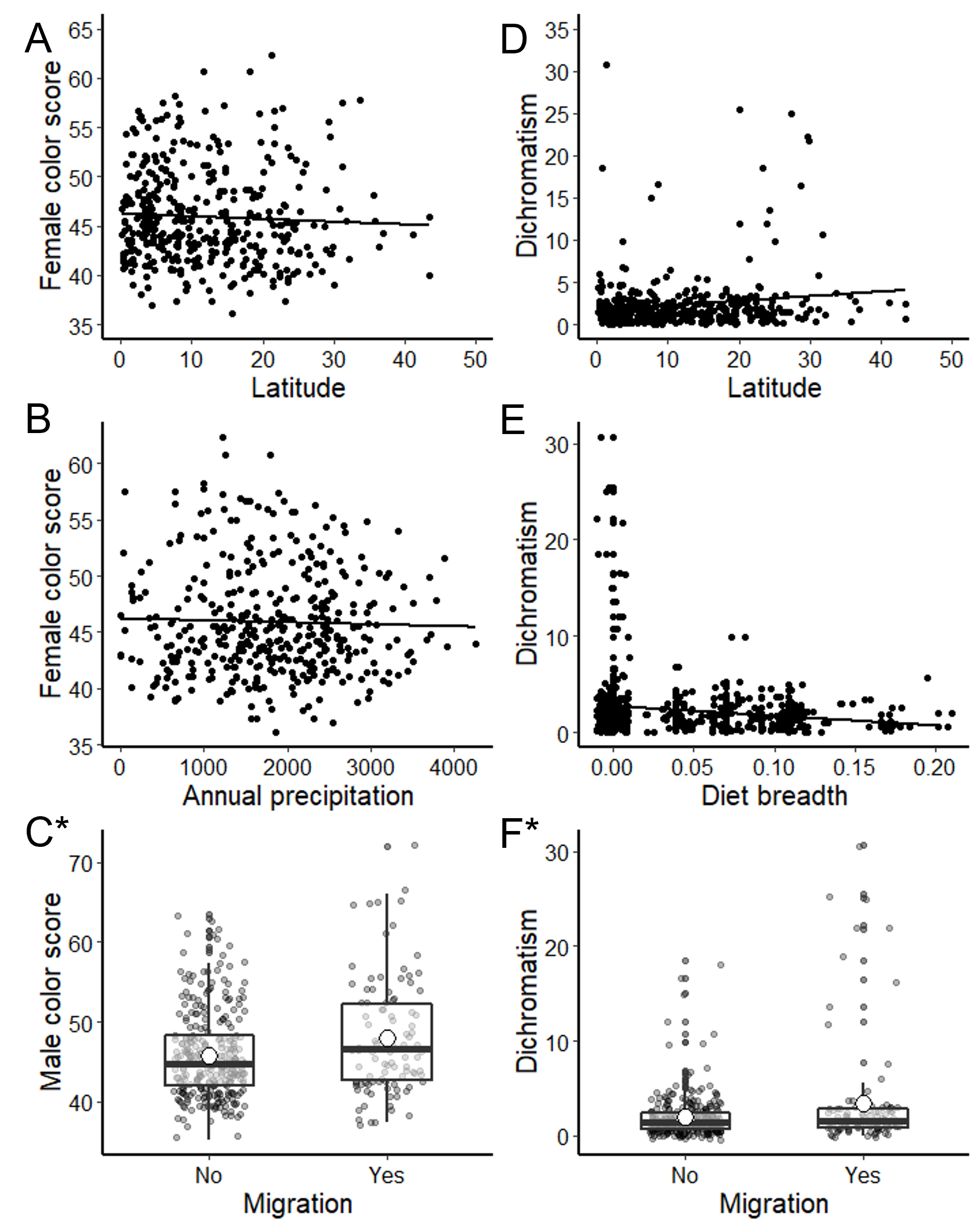
Fig. 2. Tyrannidae plumage color varied by ecology (diet breadth and migration), climate (precipitation), and geography (latitude). Figures show plumage variation for females (A, B), males (C), and sexual dichromatism (D, E, F). From phylogenetic generalized least square (PGLS) models, male color scores (C) were significantly higher and dichromatism (F) was more pronounced in migratory species (indicated by asterisks). White dots on boxplots represent mean values.
Fig. 3
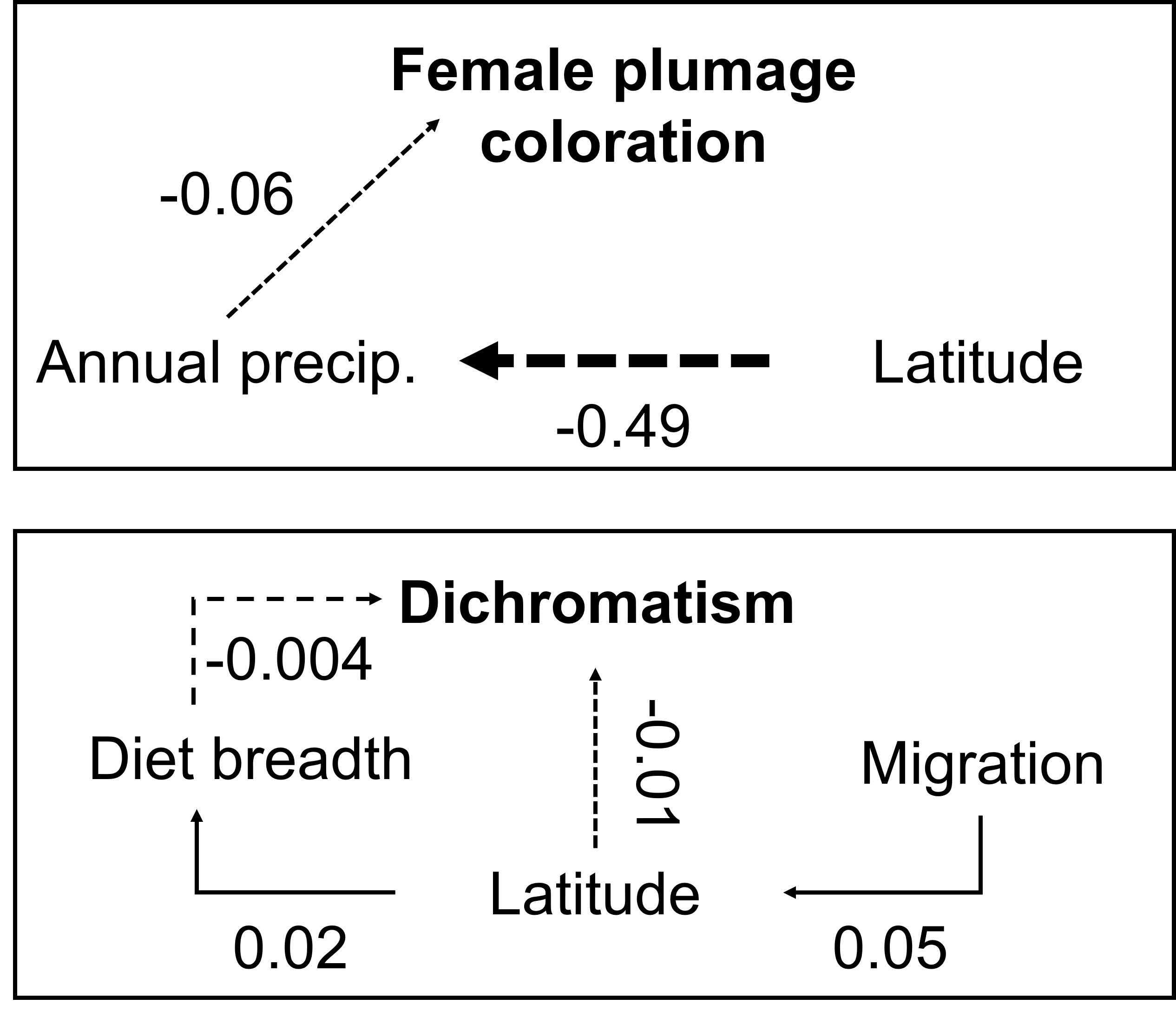
Fig. 3. Tyrannidae color evolution is directly and indirectly affected by climatic, ecological, and geographic factors. Schematic shows final phylogenetic generalized least square, i.e., PGLS-informed phylogenetic path analysis models testing the relative importance of ecological, climatic, and geographic variables on female plumage color (top panel) and sexual plumage dichromatism (bottom panel) evolution in Tyrannidae. Solid arrows represent positive effects while dashed arrows represent negative effects, and the arrows are scaled by their relative effect. Model coefficients are indicated adjacent to arrows.
Table 1
Table 1. Full and final model selection results assessing the relationships between climatic and ecological variables on Tyrannidae plumage color (female, male, dichromatism). Lambda (λ) indicates the phylogenetic signal of the response variable and was estimated using Pagel’s λ model of character evolution in a phylogenetic generalized least square analysis. Note: K: number of model parameters; BIC: Baysian information criterion score; AICc: Akaike’s information criterion score corrected for small sample sizes; ΔAIC: change between full and reduced model AICc; AICcWt: AICc weights, indicating the probability a model is the most parsimonious model; Cum.Wt: cumulative model weights; LL: log-likelihood.
| Variable | Model | Model terms | K | BIC | AICc | ΔAICc | AICcWt | Cum.Wt | LL | λ | |
| Female | Final | Latitude + Annual precipitation | 5 | 2383.7 | 2363.91 | 0 | 0.96 | 0.96 | -1176.88 | 0.37 | |
| Full | Latitude + Diet breadth + Forest cover + Annual precipitation + Maximum temperature of the warmest month + Migration | 9 | 2405.77 | 2370.33 | 6.42 | 0.04 | 1 | -1175.93 | 0.37 | ||
| Male | Final | Migration | 4 | 2542.76 | 2526.91 | 0 | 0.98 | 0.98 | -1259.4 | 0.38 | |
| Full | Latitude + Diet breadth + Forest cover + Annual precipitation + Maximum temperature of the warmest month + Migration | 9 | 2570.19 | 2534.75 | 7.84 | 0.02 | 1 | -1258.14 | 0.38 | ||
| Dichromatism | Final | Latitude + Diet breadth + Migration | 6 | 2128.27 | 2104.55 | 0 | 0.9 | 0.9 | -1046.17 | 0.29 | |
| Full | Latitude + Diet breadth + Forest cover + Annual precipitation + Maximum temperature of the warmest month + Migration | 9 | 2144.28 | 2108.84 | 4.29 | 0.1 | 1 | -1045.19 | 0.29 | ||
Table 2
Table 2. Akaike’s information criterion score (AIC) and Baysian information criterion score (BIC) selected model results from phylogenetic generalized least square models demonstrating the effects of climate and ecology on Tyrannidae color. Also included are effect sizes (ϕ, [95% CI]) for main effects. Significant results are shown in bold text.
| Response | Main Effect | Model df | Residual df | X² | p | ϕ | |||||
| Female color | Latitude | 1 | 396 | 2.93 | 0.09 | 0.07 [0, 0.18] | |||||
| Annual precipitation | 1 | 396 | 2.5 | 0.11 | 0.06 [0, 0.17] | ||||||
| Male color | Migration | 1 | 397 | 12.96 | 0.0003 | 0.17 [0.07, 0.27] | |||||
| Dichromatism | Latitude | 1 | 395 | 3.42 | 0.06 | 0.08 [0, 0.18] | |||||
| Diet breadth | 1 | 395 | 3.43 | 0.06 | 0.08 [0, 0.19] | ||||||
| Migration | 1 | 395 | 7.26 | 0.007 | 0.13 [0, 0.23] | ||||||
Table 3
Table 3. Tyrannidae coloration showed mixed support for the species recognition hypothesis among genera. Table summarizes phylogenetically controlled Mantel test results comparing Euclidean color distances and geographic proximity (km). Significant relationships based on alpha = 0.05 (p) are in bold and in italics are Mantel correlation estimates (r) whose confidence intervals (CI) did not overlap 0 but p > 0.05. n refers to the number of species in the analysis.
| Female plumage color | Male plumage color | Dichromatism | |||||||||||
| Genus | r | 95% LCI | 95% UCI | p | r | 95% LCI | 95% UCI | p | r | 95% LCI | 95% UCI | p | n |
| All genera | 0.02 | 0.01 | 0.03 | 0.18 | 0.06 | 0.05 | 0.07 | 0.02 | 0.009 | 0.003 | 0.02 | 0.39 | 399 |
| Phylloscartes | 0.08 | -0.04 | 0.20 | 0.09 | 0.11 | -0.01 | 0.23 | 0.04 | -0.02 | -0.15 | 0.10 | 0.60 | 46 |
| Myiarchus | -0.13 | -0.26 | -0.002 | 0.93 | -0.07 | -0.20 | 0.06 | 0.75 | 0.07 | -0.06 | 0.20 | 0.25 | 44 |
| Hemitriccus | 0.02 | -0.12 | 0.15 | 0.43 | 0.03 | -0.10 | 0.17 | 0.32 | -0.03 | -0.16 | 0.11 | 0.61 | 42 |
| Elaenia | -0.10 | -0.26 | 0.06 | 0.71 | -0.04 | -0.19 | 0.12 | 0.57 | -0.17 | -0.32 | -0.02 | 0.92 | 36 |
| Empidonax | -0.01 | -0.21 | 0.18 | 0.50 | 0.27 | 0.09 | 0.44 | 0.07 | 0.11 | -0.09 | 0.29 | 0.18 | 30 |
| Contopus | 0.07 | -0.14 | 0.27 | 0.28 | 0.12 | -0.09 | 0.32 | 0.19 | 0.01 | -0.20 | 0.21 | 0.35 | 28 |
| Muscisaxicola | -0.07 | -0.29 | 0.16 | 0.52 | -0.02 | -0.24 | 0.20 | 0.40 | 0.16 | -0.07 | 0.37 | 0.18 | 26 |
| Tyrannus | -0.12 | -0.34 | 0.10 | 0.88 | -0.03 | -0.25 | 0.20 | 0.52 | -0.16 | -0.37 | 0.06 | 0.99 | 26 |
| Phyllomyias | 0.15 | -0.10 | 0.38 | 0.09 | -0.005 | -0.25 | 0.24 | 0.46 | -0.09 | -0.33 | 0.15 | 0.83 | 24 |
| Poecilotriccus | 0.51 | 0.30 | 0.67 | 0.002 | 0.38 | 0.15 | 0.57 | 0.01 | -0.18 | -0.40 | 0.06 | 0.79 | 24 |
| Knipolegus | 0.11 | -0.19 | 0.39 | 0.20 | 0.04 | -0.25 | 0.33 | 0.38 | 0.03 | -0.27 | 0.32 | 0.35 | 20 |
| Ochthoeca | -0.05 | -0.34 | 0.25 | 0.58 | 0.12 | -0.18 | 0.40 | 0.27 | -0.001 | -0.29 | 0.29 | 0.42 | 20 |