







The following is the established format for referencing this article:
Preston, P. J., and R. M. Brigham. 2023. Influence of localized artificial light on calling activity of Common Poorwill (Phalaenoptilus nuttallii). Journal of Field Ornithology 94(3):16.ABSTRACT
The presence of localized artificial light at night (or ALAN) drastically changes the landscape for organisms by providing areas of darkness and light within an area that was once only dark. This change can influence the behaviors of organisms in many ways. One such behavior is bird song. Many studies have examined how ALAN influences diurnal bird song, but few have examined its influence on nocturnal birds. We examined the influence of lunar illumination and localized artificial light on the calling behavior of the nocturnal Common Poorwill (Phalaenoptilus nuttallii; hereafter poorwills). To do so, we erected artificial light stations in poorwill territories and then conducted point count surveys at these stations to count poorwill calls with the lights turned on and off. We hypothesized that artificial light would have a similar effect to lunar illumination, and we would see an increase in calling when the lights were on. However, the results we obtained were mixed. We found there was no significant effect of artificial light on calling rate and, as expected, a strong positive effect of moonlight. Surprisingly, however, there was a negative effect of the interaction between moonlight and artificial light, with birds calling less when artificial light was on during nights with high lunar illumination. One possible reason for this result is increased visibility leading to increased predation risk under high levels of ambient illumination.
RESUMEN
La presencia de luz artificial localizada durante las noches (o ALAN) cambia drásticamente el paisaje para los organismos proporcionando áreas de oscuridad y luz en un área donde anteriormente solo había oscuridad. Este cambio puede influenciar los comportamientos de los organismos de muchas maneras. Un de estos comportamientos, es el canto de las aves. Muchos estudios han examinado como ALAN influencia el canto diurno de las aves, pero pocos han examinado su influencia en las aves nocturnas. Examinamos la influencia de iluminación lunar y luces artificiales localizadas en la actividad vocal de la especie nocturna Phalaenoptilus nuttallii. Con este fin, levantamos estaciones de luz artificial en territorios de P. nuttallii y luego realizamos monitoreos por medio de puntos de conteo en estas estaciones para contar los llamados de P. nuttallii con las luces encendidas y apagadas. Hipotetizamos que la luz artificial tendría un efecto similar a la iluminación lunar, y, que observaríamos un incremento en la actividad vocal cuando las luces estuvieran encendidas. Sin embargo, encontramos resultados mixtos. Encontramos que la luz artificial no tuvo un efecto significativo sobre la tasa a la que los individuos llaman, y, como lo esperábamos, un efecto fuerte positivo de la luz de la luna. Sorprendentemente, sin embargo, encontramos un efecto negative de la interacción entre la luz de la luna y la luz artificial, pues las aves llamaron menos cuando la luz artificial estaba encendida durante las noches con alta iluminación lunar. Una posible razón para este resultado es el incremento en la visibilidad lo que lleva a un incremento en el riesgo de depredación bajo altos niveles de luz ambiental.
INTRODUCTION
Increases in urbanization over the last century have been cited as one of the greatest threats to biodiversity (Marzluff 2001). Urbanization can lead to large changes in local environments, including removal of habitat, changes in local climate by increasing temperature (Oke 1973), as well as many forms of pollution (chemical, noise, and light; Grimm et al. 2008). The use of artificial lights at night (or ALAN) is a major problem for wildlife; however, the ecological consequences of ALAN are not well understood, particularly with respect to how ALAN can affect an individual’s behavior. Localized ALAN drastically changes the landscape by providing areas of darkness and light within an area that was once only dark. This alteration can lead to changes in mating, foraging, and communication behaviors in many species (Dwyer et al. 2013, Da Silva et al. 2014, de Jong et al. 2016). The effect of ALAN may be especially strong on nocturnal organisms because their behavior is strongly influenced by lunar illumination (Gaston et al. 2013). The amount of moonlight is an important cue for nocturnal organisms, particularly those that use vision to navigate at night (Brigham et al. 1999, Reino et al. 2015, Pérez-Granados et al. 2022). Because localized ALAN can be brighter than natural moonlight, organisms that normally are influenced by moonlight can behave differently in the presence of ALAN (Sierro and Erhardt 2019). Some invertebrates that do not normally forage on moonlit nights actually increase foraging activity under increased ALAN because the illumination allows them to see their predators (Tidau et al. 2022). Although ALAN can disrupt light-induced behavioral cues that are normally associated with moonlight (Bird et al. 2004, Baker and Richardson 2006, Fobert et al. 2019), few studies examine the influence of ALAN on light-associated behaviors by birds.
Bird song behavior is greatly influenced by natural light cycles, with birds typically producing most song at dawn and dusk (Cuthill and MacDonald 1990, Hardouin et al. 2008, Kacelnik and Krebs 1982). There is considerable research on how artificial light levels influence the timing and duration of diurnal bird song. In areas effected by ALAN, many diurnal birds begin singing earlier (Da Silva et al. 2014, Da Silva and Kempenaers 2017). In addition to affecting the timing of song, recent studies show that in diurnal birds that sing at night, the effects of ALAN differ depending on how much lunar illumination is present (Dickerson et al. 2022). Birds sang more on nights with bright moonlight and when birds were not near localized artificial light sources, and there was a marked decrease in singing by males found near artificial light sources (Dickerson et al. 2022). Dickerson et al. (2022) hypothesize that bird singing behavior changed because of increased predation risk. No study, to our knowledge, has examined how the competing effects of lunar illumination and ALAN influence the singing behavior of a nocturnal or crepuscular bird species.
We examined how localized ALAN influences the calling behavior of a nocturnal bird. We chose the Common Poorwill Phalaenoptilus nuttallii (hereafter poorwills) as our model species given that males call at night and the time spent calling is heavily influenced by lunar illumination (Brigham and Barclay 1992). Poorwills are visual hunters, and their diet primarily consists of night-flying beetles and moths (Csada et al. 1992). Because they are visual hunters, they are constrained by the amount of ambient light in their environment, so they forage more at dawn and dusk and during full moons than during the dark of night. Moreover, male poorwills also call significantly more often during full moons than on dark nights (Woods et al. 2020), possibly because females are more active during these periods and also because the white display patches on their wings and tail are more visible during these times (P. J. Preston, personal observation). Male poorwills are highly territorial and use calls to signify their presence on territory and to advertise for mates (Woods et al. 2020). Therefore, there should be a trade-off between time spent calling and time spent foraging. Because poorwill calling behavior is strongly influenced by lunar phase, we expected that artificial light should also affect their behavior.
Although no prior studies have been conducted using artificial lights to assess poorwill calling, some research shows that high levels of ambient artificial light can decrease the occurrence of European nightjars in otherwise suitable habitat (Sierro and Erhardt 2019). We examined how both moonlight and localized artificial light sources influence poorwill calling behavior. We placed artificial light stations in poorwill territories and then measured poorwill calling rates. We hypothesized that the artificial light sources would have a similar effect to lunar illumination on calling by poorwills and predicted that calling would increase when the lights were on, despite a potential increase in predation risk. We expected that this effect would be especially pronounced on nights with no lunar illumination because the increase in local illumination would allow poorwills to stay active longer during the night.
METHODS
Study site
Our study took place approximately 20 km west of Osoyoos, British Columbia, Canada at two sites located approximately 3.5 km apart (N 49.020846, W −119.562293). One site was partly located on properties of the Nature Conservancy of Canada called Sparrow Grasslands and South Block and partly on Provincial Crown land that is part of the South Okanagan Grasslands Protected Area. The second site was located on a property owned by the Nature Conservancy of Canada called Sagebrush Slopes. Both sites are in areas consisting of rolling hills dominated by big sagebrush (Artemisia tridentata) and bluebunch wheatgrass (Pseudoroegneria spicata) and dotted with small stands of ponderosa pine (Pinus ponderosa) and trembling aspen (Populus tremuloides). Both sites were far enough away from large cities to have little to no skyglow at night.
To create areas of artificial illumination, light stations were placed in poorwill territories found in areas with consistent calling activity. At each station, a light post was erected following the method of Woods (2002). The post comprised a 60-W compact fluorescent light bulb (Havells-Sylvania Lighting, Saint-Etienne, France) affixed to a 2 m high metal pole. These bulbs were chosen because they generate little heat, so results would not be influenced by increased temperature near the lights. Lights were powered by a 12-V motorcycle battery, inverter, and light sensor, all housed within a plastic container. When turned on, lights provided illumination with a radius of approximately 20 m. Light intensity and irradiance spectra of these bulbs were not measured in the field. However, the manufacturer reports a light intensity of 850 lumens and irradiance spectrum of between 400 and 700 nm. The lights were set to switch on at sunset and remain on for 4 h (Noma Outdoor Lighting Timer, Inliten, Northfield, Illinois, USA).
Point count surveys of poorwill calling activity
We examined the influence of both artificial and lunar light on poorwill calling activity by conducting point counts along three routes at the two study sites beginning 30 min after sunset. Counts were made on 56 nights between 29 May and 9 September 2014, coinciding with the breeding season (Woods et al. 2020). Surveys were conducted every night, with the exception of rainy or windy (> 25 km/h) nights, when poorwills are generally inactive (Woods and Brigham 2008). Each route was surveyed for three to five nights in a row, four to five times throughout the field season. Each night, two observers would conduct 20-min point counts at each of four to six stations located along a dirt road transect, with each observer walking along the transect in opposite directions. The stations along these routes were separated by at least 500 m, corresponding to what is known about territory spacing, to ensure that each count was likely in a separate territory (Csada and Brigham 1994). During the 20-min point count, observers would record the number of poorwill calls heard and estimate the number of birds calling. To estimate the minimum number of individual birds producing the calls, observers noted whether the calls seemed to be near (< 100 m away) or far (>100 m away), the general direction of the calls, and any direct overlap in calls (two or more birds heard calling at the same time). Calls were counted from all birds detected, regardless of their estimated distance from the observer. Two poorwills were briefly seen by observers within the area illuminated by artificial light.
The first point count for each survey route began 30 min after sunset to coincide with the time when poorwills typically begin calling (Woods and Brigham 2008). Point counts were then conducted approximately every 30 min as observers moved on foot along the route. Given that we surveyed each route under different lunar conditions, we also obtained estimates of percent moon face visible from the Time and Date website for Osoyoos, British Columbia (Moon Phases 2014 – Lunar Calendar for Osoyoos, British Columbia, Canada: https://www.timeanddate.com/moon/phases/@6094665?year=2014). Observers began their survey route at different stations, so that no point counts were conducted at the same station by two observers at the same time, to avoid biasing certain stations at certain times. Each time a route was surveyed, the stations that were controls and those that were experimental were alternated, allowing each station to be surveyed at least twice as a control and twice as an experimentally lit station in each round of surveys.
From the data collected during surveys, we calculated the calling rate of poorwills for each survey. Although we assumed each station corresponded to one territory, occasionally more than one bird was heard calling during a point count survey, so calling rate was calculated by dividing the number of calls heard by the number of poorwills calling. Calling rate was used to represent overall activity by an average individual poorwill at each station.
Data analysis
We used a linear mixed-effects model to determine the effect of artificial light and moonlight on the calling rate response variable. We included the presence of artificial light, percent moon illumination, and their interaction, as well as percent cloud cover as fixed effects, and station identity (representing poorwill territories), observer (to represent the four different observers), and Julian date as random effects. Statistical significance for all analyses was set at P < 0.05, and analyses were performed using R software version 4.2.1 and the lme4 package (R Core Team 2022).
RESULTS
Influences on Common Poorwill calling activity
We conducted 372 point counts on the three survey routes: 172 were at lit stations, and 200 were at control stations. Our linear mixed-effects model analysis found significant effects of both percent moon face visible (T = 4.03, P < 0.01) and the interaction between moonlight and artificial light on calling rate (T = −3.10, P < 0.05; Table 1). Calling rate increased as the percent moon face visible increased (Fig. 1). There was a negative effect of the interaction between moonlight and artificial light, with calling rate decreasing when the light was present at higher percent illuminated moon faces (Fig. 1).
DISCUSSION
Our goal was to determine whether localized artificial light sources influenced the calling rate, and therefore activity patterns, of Common Poorwills. Contrary to our prediction, we found no significant effect of artificial light alone on calling rate (Fig. 1). In contrast to the effects of artificial light, we found that the percent moon face visible had a significant positive effect on calling rate (Fig. 1). This result is not surprising given that most nocturnal bird species spend more time both foraging and calling on moonlit nights than on dark nights (Brigham and Barclay 1992, Woods and Brigham 2008, Pérez-Granados and Schuchmann 2020). What is novel about our results is that there was also a significant effect of the interaction between moonlight and artificial light, with a decrease in calling rate when the light was on during nights with higher lunar illumination (Fig. 1). One possible reason for this result is increased visibility, and therefore potentially increased risk of predation, when there is higher ambient illumination. Studies on nocturnal insects have shown that predation risk is reduced with greater levels of artificial light, possibly because of increased risk to the insectivorous predators of the insects (Eckhartt and Ruxton 2022).
Whether poorwills are more or less vulnerable to predation on moonlit night is not well understood because few studies have directly examined the nature or frequency of predation on this species. Woods and Brigham (2008) speculated that under dim conditions such as nights with little or no lunar illumination, poorwills were more vulnerable to predation because their primary predators (owls) are less constrained by the absence of light because of better vision in dark conditions. Our results contrast those from studies of Australian Owlet-nightjars (Aegotheles cristatus) in which birds were less active on moonlit nights because of increased predation risk (Brigham et al. 1999). Ambient lunar light does not lead to an increase the abundance or density of insect prey, but it does make foraging easier for nocturnal hunters such as nightjars, and therefore, they are typically more active on bright, moonlit nights (Brigham and Barclay 1992, Jetz et al. 2003). Evidence suggests that nocturnal insectivores (including poorwills) feed around streetlights and illuminated signs (Shields and Bildstein 1979, Woods and Brigham 2008, Debrot 2014); indeed, we observed poorwills feeding at our artificial lights on two occasions.
Unfortunately, we were unable to measure the light intensity or irradiance spectra of our lightbulbs and the moonlight during our surveys; therefore, we cannot speak precisely to the difference between the two types of illumination at our light stations. We were able to calculate the average lux within the radius illuminated by our bulbs (0.17 lx), which is less than the average lux of a full moon (0.32 lx; Kyba et al. 2017); however, we did not measure the light intensity of both light sources together. Even without this measure, we were able to evaluate the interaction between lunar and artificial light in influencing poorwill calling behavior. It is possible that calling near artificial lights when lunar illumination is high makes birds conspicuous to predators and thus is a high-risk activity. Recent studies suggest that any benefits of calling when lunar illumination is high are possibly reduced by increased levels of artificial light because of increased predation risk (Dickerson et al. 2022). Dickerson et al. (2022) evaluated the influence of artificial light in a diurnal bird that sings at night and found similar results to ours: there was a reduction in singing when the moon was full and the lights were on. They suggest that increased predation risk due to the loss of dark refugia due to illumination from the moon could be a possible cause of this reduction. We did not collect any data on predation, nor did we observe any predation events; thus, it is not possible to determine if poorwills are constrained by predation risk. However, evidence suggests that poorwills may be more vulnerable to predation on dark nights because their predators such as Great Horned Owls (Bubo virginianus) have better vision in the dark and therefore are less constrained than poorwills (Woods and Brigham 2008). Great Horned Owls have been shown to hunt more vigorously during nights with greater lunar illumination (Longland and Price 1991); however, no studies have been conducted to determine the effect of moonlight on predation success in these owls. The reason for the decrease in poorwill calling when the artificial lights were on and lunar illumination was high is therefore unclear.
We found that the calling activity of poorwills was clearly influenced by moonlight. The confounding effect of the artificial lights implies that a certain amount of ambient light is ideal for poorwills to call, and too much or too little light (lunar or artificial) seems to decrease their activity. However, it remains unclear what that threshold is between just enough and too much light. The extent to which predation risk affects calling activity is difficult to examine in poorwills because it is difficult to document predation events on nocturnal species. However, this topic warrants future research. Our results clarify that localized ALAN does influence the behavior of the nocturnal Common Poorwill; however, the underlying mechanism for this change in behavior is unclear. In addition to localized ALAN, what possibly has a bigger effect is the ambient light caused by skyglow in areas around large cities. We did not measure skyglow because our study site was relatively dark, but it likely does cause a large increase in the overall ambient light that a nocturnal bird experiences (Dickerson et al. 2022). By understanding how artificial light influences nocturnal species, we can find ways to mitigate their potential negative effects.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
We thank Nicole Lerminiaux, Louis Gower, Simon Tapper, and Alyssa Walterson for assistance in the field. We also acknowledge Sara Burns and Paula Hidalgo Rodriguez for help with improving the manuscript. We thank The Nature Conservancy of Canada, the Nature Trust of British Columbia, and British Columbia Parks for allowing us to conduct research on their properties. Funding was provided by the Natural Sciences and Engineering Research Council of Canada, National Geographic, and the University of Regina. We dedicate this manuscript to Des Jackson for his lifetime of outstanding contributions to the field of nightjar research.
DATA AVAILABILITY
The data and code that support the findings of this study are openly available in Open Science Framework at https://osf.io/pxn5u/ and https://osf.io/8r5af/. No ethics approval was necessary for this project because the study species was neither handled nor disturbed.
LITERATURE CITED
Baker, B. J., and J. M. L. Richardson. 2006. The effect of artificial light on male breeding-season behaviour in green frogs, Rana clamitans melanota. Canadian Journal of Zoology 84(10):1528-1532. https://doi.org/10.1139/z06-142
Bird, B. L., L. C. Branch, and D. L. Miller. 2004. Effects of coastal lighting on foraging behavior of beach mice. Conservation Biology 18(5):1435-1439. https://doi.org/10.1111/j.1523-1739.2004.00349.x
Brigham, R. M., and R. M. R. Barclay. 1992. Lunar influence on foraging and nesting activity of Common Poorwills (Phalaenoptilus nuttallii). Auk 109(2):315-320. https://doi.org/10.2307/4088200
Brigham, R. M., R. C. A. Gutsell, R. S. Wiacek, and F. Geiser. 1999. Foraging behaviour in relation to the lunar cycle by Australian Owlet-nightjars Aegotheles cristatus. Emu - Austral Ornithology 99(4):253-261. https://doi.org/10.1071/MU99031
Csada, R. D., and R. M. Brigham. 1994. Breeding biology of the Common Poorwill at the northern edge of its distribution. Journal of Field Ornithology 65(2):186-193. https://sora.unm.edu/node/51877
Csada, R. D., R. M. Brigham, and B. R. Pittendrigh. 1992. Prey selection in relation to insect availability by the common poorwill (Phalaenoptilus nuttallii). Canadian Journal of Zoology 70(7):1299-1303. https://doi.org/10.1139/z92-181
Cuthill, I. C., and W. Macdonald. 1990. Experimental manipulation of the dawn and dusk chorus in the blackbird Turdus merula. Behavioral Ecology and Sociobiology 26(3):209-216. https://doi.org/10.1007/BF00172088
Da Silva, A., and B. Kempenaers. 2017. Singing from North to South: latitudinal variation in timing of dawn singing under natural and artificial light conditions. Journal of Animal Ecology 86(6):1286-1297. https://doi.org/10.1111/1365-2656.12739
Da Silva, A., J. M. Samplonius, E. Schlicht, M. Valcu, and B. Kempenaers. 2014. Artificial night lighting rather than traffic noise affects the daily timing of dawn and dusk singing in common European songbirds. Behavioral Ecology 25(5):1037-1047. https://doi.org/10.1093/beheco/aru103
de Jong, M., J. Q. Ouyang, R. H. A. van Grunsven, M. E. Visser, and K. Spoelstra. 2016. Do wild Great Tits avoid exposure to light at night? Plos One 11(6):e0157357. https://doi.org/10.1371/journal.pone.0157357
Debrot, A. O. 2014. Nocturnal foraging by artificial light in three Caribbean bird species. Journal of Caribbean Ornithology 27:40-41. https://jco.birdscaribbean.org/index.php/jco/article/view/78
Dickerson, A. L., M. L. Hall, and T. M. Jones. 2022. The effect of natural and artificial light at night on nocturnal song in the diurnal willie wagtail. Science of the Total Environment 808:151986. https://doi.org/10.1016/j.scitotenv.2021.151986
Dwyer, R. G., S. Bearhop, H. A. Campbell, and D. M. Bryant. 2013. Shedding light on light: benefits of anthropogenic illumination to a nocturnally foraging shorebird. Journal of Animal Ecology 82(2):478-485. https://doi.org/10.1111/1365-2656.12012
Eckhartt, G. M., and G. D. Ruxton. 2022. Artificial light at night may decrease predation risk for terrestrial insects. Biology Letters 18:20220281. https://doi.org/10.1098/rsbl.2022.0281
Fobert, E. K., K. Burke da Silva, and S. E. Swearer. 2019. Artificial light at night causes reproductive failure in clownfish. Biology Letters 15:20190272. https://doi.org/10.1098/rsbl.2019.0272
Gaston, K. J., J. Bennie, T. W. Davies, and J. Hopkins. 2013. The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biological Reviews 88(4):912-927. https://doi.org/10.1111/brv.12036
Grimm, N. B., S. H. Faeth, N. E. Golubiewski, C. L. Redman, J. Wu, X. Bai, and J. M. Briggs. 2008. Global change and the ecology of cities. Science 319(5864):756-760. https://doi.org/10.1126/science.1150195
Hardouin, L. A., D. Robert, and V. Bretagnolle. 2008. A dusk chorus effect in a nocturnal bird: support for mate and rival assessment functions. Behavioral Ecology and Sociobiology 62:1909-1918. https://doi.org/10.1007/s00265-008-0621-5
Jetz, W., J. Steffen, and K. E. Linsenmair. 2003. Effects of light and prey availability on nocturnal, lunar and seasonal activity of tropical nightjars. Oikos 103(3):627-639. https://doi.org/10.1034/j.1600-0706.2003.12856.x
Kacelnik, A., and J. Krebs. 1982. The dawn chorus in the great tit (Parus major): proximate and ultimate causes. Behaviour 83(3-4):287-309. https://doi.org/10.1163/156853983X00200
Kyba, C. C. M., A. Mohar, and T. Posch. 2017. How bright is moonlight? Astronomy and Geophysics 58:1.31-1.32. https://doi.org/10.1093/astrogeo/atx025
Longland, W. S., and M. V. Price. 1991. Direct observations of owls and heteromyid rodents: Can predation risk explain microhabitat use? Ecology 72(6):2261-2273. https://doi.org/10.2307/1941576
Marzluff, J. M. 2001. Worldwide urbanization and its effects on birds. Pages 19-47 in J. M. Marzluff, R. Bowman, and R. Donnelly, editors. Avian ecology and conservation in an urbanizing world. Springer, New York, New York, USA. https://doi.org/10.1007/978-1-4615-1531-9_2
Oke, T. R. 1973. City size and the urban heat island. Atmospheric Environment (1967) 7(8):769-779. https://doi.org/10.1016/0004-6981(73)90140-6
Pérez-Granados, C., and K.-L. Schuchmann. 2020. Illuminating the nightlife of two Neotropical nightjars: vocal behavior over a year and monitoring recommendations. Ethology Ecology and Evolution 32(5):466-480. https://doi.org/10.1080/03949370.2020.1753117
Pérez-Granados, C., K.-L. Schuchmann, and M. I. Marques. 2022. Addicted to the moon: vocal output and diel pattern of vocal activity in two Neotropical nightjars is related to moon phase. Ethology Ecology and Evolution 34(1):66-81. https://doi.org/10.1080/03949370.2021.1886182
R Core Team. 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna Austria. https://www.R-project.org/
Reino, L., M. Porto, J. Santana, and T. S. Osiejuk. 2015. Influence of moonlight on nightjars’ vocal activity: a guideline for nightjar surveys in Europe. Biologia 70:968-973. https://doi.org/10.1515/biolog-2015-0099
Shields, W. M., and K. L. Bildstein. 1979. Bird versus bats: behavioral interactions at a localized food source. Ecology 60(3):468-474. https://doi.org/10.2307/1936065
Sierro, A., and A. Erhardt. 2019. Light pollution hampers recolonization of revitalised European Nightjar habitats in the Valais (Swiss Alps). Journal of Ornithology 160:749-761. https://doi.org/10.1007/s10336-019-01659-6
Tidau, S., J. Whittle, S. R. Jenkins, and T. W. Davies. 2022. Artificial light at night reverses monthly foraging pattern under simulated moonlight. Biology Letters 18:20220110. https://doi.org/10.1098/rsbl.2022.0110
Woods, C. P. 2002. Ecological aspects of torpor use and inactivity during winter by Common Poorwills. Dissertation. University of Regina, Regina, Canada. http://hdl.handle.net/10294/14088
Woods, C. P., and R. M. Brigham. 2008. Common Poorwill activity and calling behavior in relation to moonlight and predation. Wilson Journal of Ornithology 120(3):505-512. https://doi.org/10.1676/06-067.1
Woods, C. P., R. D. Csada, and R. M. Brigham. 2020. Common Poorwill (Phalaenoptilus nuttallii), version 1.0. In A. F. Poole, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.compoo.01
Fig. 1
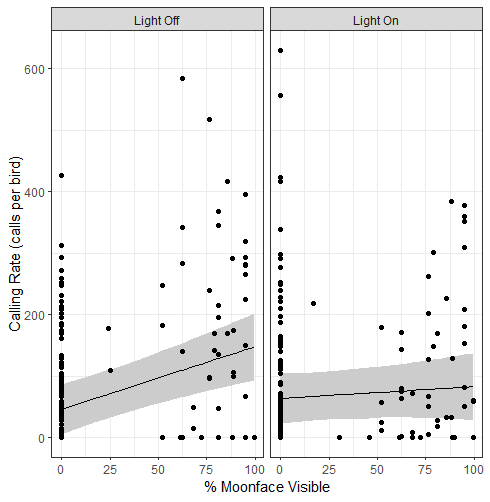
Fig. 1. Variation in Common Poorwill calling rate (calls/bird; ± 95% confidence interval) recorded during point counts conducted at stations with an artificial light on (right panel) and off (left panel) in relation to percentage of moon face visible. A linear mixed-effects model analysis showed that lunar condition positively influenced calling rate, whereas the interaction between moonlight and experimental condition negatively influenced calling rate, indicated by the comparatively lower calling rate when the light is on and the moon is nearing 100% visible.
Table 1
Table 1. Results of linear mixed-effects model of predictors of the mean calling rate of Common Poorwills. The percent moon face visible positively influenced calling rate, whereas the interaction between artificial light and moonlight negatively influenced calling rate. Neither artificial light alone nor cloud cover had a significant effect on calling rate. Station identity, survey date, and observer identity were included as random effects.
| Effect | Estimate | Standard error | T | P | Variance | Standard deviation | ||||
| Intercept | 46.56 | 18.45 | 2.233 | 0.355 | ||||||
| Artificial light | 17.59 | 10.65 | 1.651 | 0.997 | ||||||
| Moon face visible | 1.029 | 0.2552 | 4.032 | < 0.0001 | ||||||
| Artificial light & times moon face visible | −0.8316 | 0.2685 | −3.097 | 0.002 | ||||||
| Percent cloud cover | 0.1489 | 0.2244 | 0.664 | 0.507 | ||||||
| Station† | 3229.6 | 57.44 | ||||||||
| Julian date† | 2391.2 | 48.9 | ||||||||
| Observer† | 262.9 | 16.21 | ||||||||
| † Random effect. | ||||||||||