







The following is the established format for referencing this article:
Lewis, E. A., and C. A. Barber. 2023. Cooperative nest defense by European Starlings (Sturnus vulgaris) during a predatory threat. Journal of Field Ornithology 94(3):13.ABSTRACT
Cooperative nest defense has been documented in various passerine species. Parents typically swoop and alarm-call at any predator near their nest, often attracting predominantly male conspecifics to help with nest defense. Potential reasons for males to engage in communal nest defense include direct benefits such as deterring a predator from their own nest area, by-product mutualism (paternity uncertainty in nearby nests), reciprocity, kin selection, and quality advertisement. European Starlings (Sturnus vulgaris) are a semicolonial and gregarious cavity-nesting passerine with biparental care. They have a mixed reproductive strategy that includes both extra-pair paternity and intraspecific brood parasitism. Therefore, both paternity and maternity uncertainty could occur in conspecific nests. Our objective was to examine whether conspecific nest defense occurred in this species, and if it did, whether both males and females participated. We exposed adult European Starlings breeding in 16 nest boxes to a taxidermy mount of an American red squirrel (Tamiasciurus hudsonicus; experimental treatment) and a similarly sized and shaped rock (control) mid-way through the nestling period when nestlings were 11 or 12 days old. Significantly more starlings (parents and conspecifics) responded in the experimental than control trials, and they responded with a significantly higher aggregate score of defensive responses, demonstrating both the effectiveness of the taxidermy mount in eliciting defensive responses and the presence of cooperative nest defense in this species. Both males and females participated in mobbing at conspecific nests during the experimental trials. This study is the first to determine that male and female European Starlings engage in cooperative defense of conspecific nests.
RESUMEN
La defensa cooperativa del nido ha sido documentada en varias especies de aves paseriformes. Los parentales, típicamente arremeten y generan alarmas ante un depredador cerca de sus nidos, con frecuencia atrayendo predominantemente individuos machos de la misma especie para ayudar con la defensa del nido. Las razones potenciales para que los machos se involucren en la defensa comunal de los nidos incluyen beneficios como, ahuyentar al depredador del área de sus propios nidos, un subproducto de mutualismos (paternidad incierta de los nidos cercanos), reciprocidad, selección de parentesco y calidad de las advertencias. Sturnus vulgaris es una especie semicolonial y gregaria que anida en cavidades con cuidado biparental. Tienen una estrategia de reproducción mixta que incluye paternidad extra parental y parasitismo intraespecífico de nido. Por lo tanto, la incertidumbre en la paternidad y la maternidad puede ocurrir en nidos conespecíficos. Nuestro objetivo fue el de examinar si la defensa de nido conespecífica ocurre en esta especie, y en caso de ocurrir, si ambos, el macho y la hembra participan. Expusimos a individuos adultos de Sturnus vulgaris anidando en 16 cajas de anidación, a un montaje en taxidermia de una Ardilla Americana roja (Tamiasciurus hudsonicus; tratamiento experimental) y a una piedra de similar tamaño (control) en la mitad del periodo de anidación cuando los pichones tenían 11 o 12 días de nacidos. Significativamente un mayor número de individuos (parentales y conespecíficos) respondieron al tratamiento experimental que, al control, y respondieron con un puntaje agregado de respuestas defensivas significativamente mayor, demostrando la efectividad del montaje de taxidermia y provocando respuestas defensivas y la presencia de defensa cooperativa de los nidos en esta especie. Este estudio es el primero en determinar que los machos y las hembras de Sturnus vulgaris participan en la defensa cooperativa de nidos conespecíficos.
INTRODUCTION
Avian parents typically respond to a predator’s presence near their nest by alarm calling (Curio 1978) and swooping at the predator, which often serve to attract nearby conspecifics to help defend the nest. Cooperative nest defense has been documented in several passerine species (Beletsky and Orians 1989, Olendorf et al. 2004, Grabowska-Zhang et al. 2012a, Krams et al. 2022) and consists of increased vigilance (Lima and Dill 1990), alarm calls (Trivers 1971), and mobbing (da Cunha et al. 2017a), which can be both costly and risky to the individuals involved. Males are predominantly the ones participating in defending conspecific nests (da Cunha et al. 2017b, Krams et al. 2022) and, in doing so, may gain direct benefits (Riehl 2013) such as proactively keeping the predator away from their own nest. Other possibilities include reciprocal altruism whereby an unrelated individual helps another at a cost to themself; the individual receiving the aid returns that help at a later date (e.g., Olendorf et al. 2004), as well as kin selection models whereby an individual helps increase the fitness of a relative at a cost to themself (Colombelli- Négrel and Evans 2017). Still other possibilities include males advertising their quality to females (da Cunha et al. 2017b) or potentially having paternity in those nests (a by-product mutualism in which an individual is helping for selfish means, but the recipient also benefits; Brown 1987, Brown and Brown 1990). When females engage in extra-pair copulations, it is most often with nearby neighbors (Gibbs et al. 1990, Westneat 1993), resulting in paternity uncertainty. According to by-product mutualism, paternity uncertainty would result in males being more likely to work together in nest defense against predators. Eliassen and Jørgensen (2014) predicted that although males would cooperate toward the public good due to paternity uncertainty, females would not because they typically produce genetic offspring only in their own nest.
The European Starling, Sturnus vulgaris, is a gregarious, semicolonial-breeding passerine that nests in cavities (Feare 1984). European Starlings generally have two broods per year (Feare 1984) and are classified as socially monogamous yet facultatively polygynous breeders (Pinxten et al. 1989). Both parents care for the offspring by incubating the eggs (Pinxten et al. 1993a) and feeding the nestlings (Kessel 1957), but surprisingly little is known about their investment in nest defense. Offspring fledge when they are 21–24 days old but still depend on parental care for 1–2 weeks thereafter (Feare 1984). Extra-pair fertilizations are part of their mixed reproductive strategy (Pinxten et al. 1993b) and have been documented in our study population (Slade 2012), so males are likely uncertain of their genetic paternity both within their social nest and those of conspecifics. Maternity uncertainty also exists because of the occurrence of intraspecific brood parasitism (IBP) whereby females lay eggs in conspecific nests (Pinxten et al. 1991, 1993b, Sandell and Diemer 1999, Slade 2012). The past frequency of IBP in our study population over a 4-yr period ranged from 30–75% of nests containing IBP offspring and 11–46% of all young produced being a result of IBP (unpublished data).
The objective of our study was to examine whether conspecific nest defense occurs in European Starlings, and if so, whether both males and females participate. We first determined whether a taxidermy mount of an American red squirrel, Tamiasciurus hudsonicus, was effective in eliciting nest defense responses from European Starlings when presented in experimental trials. American red squirrels are a natural predator of songbird eggs and nestlings (Sieving and Wilson 1998, Bayne and Hobson 2002) and are common at our study site. Control trials consisted of a rock, similar in size and shape to that of the taxidermy mount. We predicted that European Starlings would cooperate in conspecific nest defense. We also predicted that more starlings would be actively involved in communal defense during experimental than control trials, and that aggregate nest defense scores would be higher in the experimental trials than the control trials. We expected that females as well as males would assist in conspecific nest defense because it would result in direct benefits (Riehl 2013) by helping deter predators from attacking their own nest, or because both sexes could potentially have genetic offspring in conspecific nests (Slade 2012). Finally, we predicted that an increased aggregate score in experimental trials would be positively correlated with both social reproductive success (number and proportion of nestlings who fledged) and nest success (fledging at least one offspring) because higher responses to potential predators would be expected to increase reproductive success.
METHODS
We conducted this study from May through July 2015 on the campus of Saint Mary’s University, Halifax, Nova Scotia, Canada (44°37’54.07” N, 63°34’47.09” W; Fig. 1). The study site consists of mature trees and shrubs interspersed among large grassy areas, parking lots, and buildings on land covering one city block. Forty-five wooden nest boxes were attached to trees, 2.4–3.7 m above the ground. Nest boxes were 41.9 cm tall in the back and 35 cm tall in the front, with a sloping roof. They were 17 cm wide, 18 cm deep, and had a 5.1 cm diameter opening in the center front of the box. They were placed ≥ 8 m apart. We conducted both control and experimental trials on 16 nest boxes. Not all nest boxes were occupied (occupied/available: 23/45 early vs. 16/45 late season). We captured adults using a simple nest box trap (Stutchbury and Robertson 1986) and determined their sex (Kessel 1951) when nestlings were ≥ 5 days old (hatch day = day 0). We banded adults with a unique color combination of two bands on the left tarsus and one band on the right tarsus above a Canadian Wildlife Service band. Of these banded birds, 27 were female and 20 were male.
On the 11th day of the nestling period, we presented one of two models to each nest box: either a taxidermy mount of an American red squirrel (experimental treatment; Fig. 2), or a rock similar in size, shape, and coloring to the squirrel (control treatment; Fig. 3). The squirrel had glass eyes, was positioned on all four legs, and was 17.8 cm long and 7.6 cm wide, whereas the rock was 22.9 cm long and 6.4 cm wide. Then 24–48 h later, we presented the second model (e.g., Naďo et al. 2018). We randomly assigned presentation order at each nest box by flipping a coin. We placed the model (rock or squirrel) on top of the focal nest box, approximately 12.7 cm vertically from the hole, when the parents were absent. Each trial was 3 min long and began when the first adult arrived at the nest box. Trials occurred between 8:30 and 13:30 and were recorded with a digital camera, with every attempt made to remain out of view of the birds. If two neighboring nest boxes (within a radius of 80 m) were tested on the same day and one was randomly assigned the control treatment, we conducted that presentation first. If two neighboring nest boxes tested on the same day were assigned the experimental treatment, the trial at one of the nests was delayed for a few hours.
Observations recorded during the experimental and control trials comprised: (1) date, time, and age of nestlings; (2) number of adults (parents and conspecifics) present; (3) band combinations and sex of participating adults; and numbers of (4) hits and (5) dives made to the model (Blancher and Robertson 1982), (6) fly-bys (within 0.5 m of the model), (7) attack (alarm) calls (Cabe 2020), and (8) chip (warning/mobbing) vocalizations (Cabe 2020). Band color combinations permitted the identification of most adults around the focal nest box, although not all participants were banded. No adults abandoned their nest after these trials, as was found in other studies on cavity-nesting species (e.g., Fisher and Wiebe 2006, Stanback et al. 2018).
Nest defense intensity was quantified by using aggregate scores of defensive strategies that occurred at least once during the 3-min trial. If any of the following behaviors were present within the 3-min trial, it was awarded a score of 0 to 4 (least to most aggressive): no reaction (0), alarm calls or chips (1), fly-bys (2), dives (3), and hits (4) (e.g., Siderius 1993, Krama et al. 2012a). Each behavior, if present, was only counted once per trial, resulting in a potential score that ranged from 0 (no response) to 10 (all defensive behaviors observed). The maximum score of 10 would have all defensive responses present (hits, dives, fly-bys, and alarm calls/chips). To ensure that time of season did not affect the findings, we did a Mann-Whitney test on aggregate scores to determine if they differed between the 11 early (15–26 May) and five late (30 June–5 July) broods for each of the control and experimental trials. They did not (Control: U = 14.5, N1 = 11, N2 = 5, P = 0.18; Experimental: U = 24.5, N1 = 11, N2 = 5, P = 0.77), so we pooled the data.
We tested data for normality using Graph Pad Prism 9 (San Diego) for analysis. A Wilcoxon matched pairs signed-rank test was used to analyze the total number of starlings flying to the focal nest box in experimental vs. control trials, as well as to analyze aggregate scores toward the American red squirrel vs. the rock. We conducted a Fisher’s Exact test to determine whether the number of conspecifics differed between experimental and control trials as well as whether parents defended their own nest more often in experimental than control trials. Fisher’s Exact tests were also used to test for differences between experimental and control trials when examining the occurrence of (1) conspecific nest defense by each sex, and (2) each of the four defensive strategies. We used Spearman correlation tests to determine whether a relationship existed between aggregate scores and social reproductive success (the number and proportion of offspring fledged). To determine if there was relationship between nest success (fledged at least one offspring) and aggregate scores, we ran a generalized linear model with binomial distribution and logit link function on the experimental trials, with fledge as the binomial response variable and aggregate score as the predictor variable using R version 4.2.3 (R Core Team 2022). All tests were two-tailed, and results were considered significant when P ≤ 0.05.
RESULTS
We found that conspecific nest defense does occur in this species, and both males and females participated in this communal nest defense. Significantly more birds (parents and conspecifics) flew to the focal nest box during the experimental than the control trials (Table 1; Wilcoxon W = 45, N = 16, P = 0.004; Fig. 4). Conspecifics (not including parents) also actively responded more often during the experimental (56.3%) than control (0%) trials (Fisher’s Exact test P = 0.0008). Up to seven conspecifics responded in experimental trials. Only parents and conspecifics responded in 15/16 of experimental trials, and two Common Grackles (Quiscalus quiscula) joined in nest defense in the other experimental trial.
Aggregate scores in response to the American red squirrel were also significantly higher than those to the rock (W = 120, N = 16, P < 0.0001; Fig. 5). Reactions by parents in control trials ranged from briefly looking at the rock as they flew over before directly entering the nest box with food for the young (6/16 nests) to being hesitant, approaching between 15 cm and 1.8 m of the nest box but then either flying to a nearby perch (7/16) or flying away with food still in their beak (3/16). Parents never entered the nest box during the experimental trials.
Females and males defended their own nest significantly more often during the experimental than the control trials (Fisher’s Exact test, P = 0.0001 and P = 0.002, respectively). They also defended their own nest in all the experimental trials in which they were present (11/16 for females and 9/16 for males). Females did not defend conspecific nests significantly more often in the experimental (3/16) than the control (0/16) trials (Fisher’s Exact test, P = 0.23), whereas males did (6/16 vs. 0/16 trials, respectively; Fisher’s Exact test, P = 0.02). Of the 20 conspecifics who responded in the experimental trials, 11 were males (three were banded), three were females (two were banded), and six were of unknown sex. In two separate instances, mobbers were confirmed to be nearby neighbors through the presence of color bands. One male defended his own nest during the experimental trial and then also defended that of his neighbor on another day.
Parents and conspecifics reacted strongly to the squirrel. Alarm calls/chips were the most common defense tactic, occurring in 16/16 experimental trials, with a range of 62–262 alarm calls and 0–45 chips in 3 min (Table 1). Fly-bys were the next most common defense tactic, occurring in 13/16 experimental trials. Finally, direct hits at the squirrel model occurred in 4/16 trials; all hits were made by the female parent (4, 7, 20, and 23 hits; Table 1). Experimental trials had significantly more occurrences of alarm calls/chips (Fisher’s Exact test, P = 0.002) and fly-bys (Fisher’s Exact test, P = 0.004) than did control trials and tended also to have more occurrences of hits (Fisher’s Exact test, P = 0.10), although no hits were made to the rock. Finally, although the occurrence of dives did not differ significantly between experimental and control trials (Fisher’s Exact test, P = 0.23), no dives were made at the rock.
Nest success was high in 2015, with only 2/16 nests failing to fledge any offspring. One of these nests failed due to predation; the other nest failed because the nestlings appeared to have starved. We found no relationship between the aggregate scores of experimental trials and either the number of offspring fledged (rs = −0.23, N = 16, P = 0.40) or the proportion fledged from the focal nest (rs = −0.20, N = 16, P = 0.45). There was also no significant effect of aggregate score on nest success in the experimental trials (odds ratio = 1.014, P = 0.97).
DISCUSSION
European Starlings participated in conspecific nest defense when confronted with a predatory threat. As predicted, significantly more starlings defended the focal nest box and did so more aggressively during experimental than control trials. Conspecifics were absent during control trials, whereas between one and seven conspecifics aided in communal nest defense during experimental trials. The American red squirrel taxidermy mount was effective in eliciting nest defense responses by adult European Starlings. Other studies have also effectively used taxidermy mounts to simulate a predatory threat in both open-cup (Neudorf and Sealy 1992, Siderius 1993, Redmond et al. 2009, Vrublevska et al. 2015) and cavity-nesting passerines (Krams et al. 2008, 2022, Krama et al. 2012a, Stanback et al. 2018). Defensive strategies displayed by parents and conspecifics consisted of alarm calls and chips, fly-bys to examine the squirrel, dives, and direct hits, which are similar to those displayed by other passerines such as Eastern Kingbirds (Tyrannus tyrannus; Siderius 1993) and Pied Flycatchers (Ficedula hypoleuca; Krama et al. 2012a). Participating adults also displayed wing flicking and wing waving behaviors. No hits were attempted during control trials, but they occurred at 4/16 focal nests during the experimental trials.
Both males and females actively defended their own nest during the experimental trials. We were unable to determine which sex defended their nest more intensely, other than observing that female parents were the only ones hitting the stuffed squirrel model, which is a risky endeavor. In other studies, males were found to have stronger responses in nest defense to a simulated predator. For example, in the cavity-nesting Eurasian Nuthatch (Sitta europaea), both parents engaged in nest defense, but males were more aggressive than females, attacking the simulated intruder significantly more often, whereas females were more actively engaged in threat displays (Naďo et al. 2018). Male Mountain Bluebirds (Sialia currucoides), another cavity nester, defended their nest more aggressively from an American red squirrel model than did females (Tkaczyk et al. 2022). A similar result was found in Eastern Screech-owls (Megascops asio) whereby males were more aggressive than females in defending their nestlings (Sproat and Ritchison 1993). Similarly, male Eastern Kingbirds, an open cup nester, defended their own nests more intensely than did females (Redmond et al. 2009), as did Northern Mockingbird (Mimus polyglottos) males (Breitwisch 1988). Future studies of European Starlings should separate male and female responses as well as parental and conspecific responses, both as to the type of response and its frequency of occurrence.
Our study is the first to document cooperative nest defense by conspecifics in European Starlings. However, other passerines engage in communal nest defense: Red-winged Blackbirds (Agelaius phoeniceus, Olendorf et al. 2004), Pied Flycatchers (Krams et al. 2008, 2022), and Great Tits (Parus major, Grabowska-Zhang et al. 2012a, Vrublevska et al. 2015). Typically, either more males than females participate in mobbing conspecifics (e.g., da Cunha et al. 2017b) or only males do so (e.g., Krams et al. 2022). We found that both males and females helped conspecifics in defending their nest against a simulated predatory threat, although females did so less frequently. Eliassen and Jørgensen (2014) suggest that females would be unlikely to cooperate in conspecific nest defense unless they had maternity uncertainty in those nests. Although our study is the first to explore this possibility with a species that potentially has maternity uncertainty within conspecific nests, we cannot make conclusions as to why they engaged in conspecific nest defense. The various hypotheses for cooperative nest defense are not necessarily mutually exclusive but could be tested separately in European Starlings, as was done for Red-winged Blackbirds (Olendorf et al. 2004). To test for by-product mutualism, conspecifics who helped in nest defense would have their parentage examined within the nests they defended. Krams et al. (2022) found support for by-product mutualism in Pied Flycatchers, another semicolonial, nest box breeding passerine, by observing that males were more likely to mob and aggressively defend conspecific nests in which they had genetic offspring during a simulated predatory attack. To test the reciprocal altruism hypothesis in which an individual helps another and then receives help from that individual in the future (Olendorf et al. 2004, Krams et al. 2008, Krama et al. 2012b), a conspecific male would be temporarily captured such that he could not help to defend a neighbor’s nest during a simulated predation attempt. He would then be released, and it would be determined if his neighbor subsequently reduced his help during a future simulated predation event. Olendorf et al. (2004) found support for reciprocity in Red-winged Blackbirds, as males decreased their conspecific nest defense after a simulated defection by their neighbor. To test the kin selection hypothesis, in which an individual helps another because they are genetically related (Colombelli-Négrel and Evans 2017), DNA analyses to determine relatedness would be conducted on each of the conspecific males who cooperated in nest defense and those who were helped. Finally, male and female European Starlings may also help defend conspecific nests to deter predators from their own nest because they breed close to each other (direct benefits; Riehl 2013). It would be expected that conspecifics who cooperate in nest defense would suffer less predation on their own nests.
We detected one case of heterospecific mobbing, with two Common Grackles joining European Starlings during an experimental trial. Krama et al. (2012a) found that mobbing was more intense when breeding density was higher in Pied Flycatchers, but only neighboring heterospecific passerines engaged in the communal nest defense; no conspecifics helped. Heterospecific communal nest defense was also seen in Chaffinches (Fringilla coelebs), of which European Starlings were one of several passerine species to mob a taxidermied Tawny Owl (Strix aluco, Krams and Krama 2002), albeit not as actively as other species. Future studies could examine the frequency and value of heterospecific nest defense in and by European Starlings. Similarly, it would be interesting to examine whether starlings are more likely to help conspecifics in nest defense during the nestling stage over the egg stage, as was seen in male Eastern Screech-owls (Sproat and Ritchison 1993) and male Mountain Bluebirds (Tkaczyk et al. 2022). Another potential study would be to examine whether starlings have greater reproductive success in higher density than lower density breeding neighborhoods. Robinson (1985) found that coloniality helped reduce predation of Yellow-rumped Cacique (Cacicus cela) nests in several ways, one of which was to enhance group defense of nests by mobbing. It would also be interesting to determine whether starlings respond more to certain types of predators over others (e.g., reptile vs. mammal vs. avian predators) who are small enough to enter a nest box. Kleindorfer et al. (2005) tested this idea in three Acrocephalus species and found that one species nesting close to the ground responded more to snakes while another species nesting high up had the strongest response to harriers.
Many banded starlings return to breed in our nest boxes every year. Advantages to breeding with familiar neighbors have been documented and include increased cooperation in predator mobbing and enhanced reproductive success (Beletsky and Orians 1989, Grabowska-Zhang et al. 2012a,b). However, contrary to our prediction, we did not find a positive relationship between aggregate nest defense scores in experimental trials and either fledging success or nest success, although our sample size was small. Redmond et al. (2009) also found no relationship between aggregate nest defense scores and nest success in a population of Eastern Kingbirds. Nest predation in our study population was only 6.3% in 2015, and nest success was high, so our findings may vary depending on the year. To better test the relationship between behavioral responses of parents to predators and fledging success, such a study should be conducted in areas having higher levels of predation.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Both authors contributed equally to this work.
ACKNOWLEDGMENTS
We are grateful to Alyssa Walthers and Celina Campbell for their help with the field work. Erin Cameron gave statistical advice, and Jennifer Foote gave constructive suggestions. We are very grateful to three reviewers for their helpful suggestions. Our research was funded by Saint Mary’s University through a grant from SMUworks as well as a research award from the Faculty of Graduate Studies and Research.
DATA AVAILABILITY
The data from this study are available at Borealis via https://doi.org/10.5683/SP3/ZGQQSK.
LITERATURE CITED
Bayne, E. M., and K. A. Hobson. 2002. Effects of red squirrel (Tamiasciurus hudsonicus) removal on survival of artificial songbird nests in boreal forest fragments. American Midland Naturalist 147(1):72-79. https://doi.org/10.1674/0003-0031(2002)147[0072:EORSTH]2.0.CO;2
Beletsky, L. D., and G. H. Orians. 1989. Familiar neighbors enhance breeding success in birds. Proceedings of the National Academy of Sciences 86(20):7933-7936. https://doi.org/10.1073/pnas.86.20.7933
Blancher, P. J., and R. J. Robertson. 1982. Kingbird aggression: Does it deter predation? Animal Behaviour 30(3):929-930. https://doi.org/10.1016/S0003-3472(82)80167-X
Breitwisch, R. 1988. Sex differences in defence of eggs and nestlings by northern mockingbirds, Mimus polyglottos. Animal Behaviour 36(1):62-72. https://doi.org/10.1016/S0003-3472(88)80250-1
Brown, J. L. 1987. Helping communal breeding in birds: ecology and evolution. Princeton University Press, Princeton, New Jersey, USA. https://doi.org/10.1515/9781400858569
Brown, J. L., and E. R. Brown. 1990. Mexican Jays: uncooperative breeding. Pages 267-288 in P. B. Stacey, and W. D. Koenig, editors. Cooperative breeding in birds: long-term studies of ecology and behaviour. Cambridge University Press, Cambridge, UK. https://doi.org/10.1017/CBO9780511752452.010
Cabe, P. R. 2020. European Starling (Sturnus vulgaris). Version 1.0. In S. M. Billerman, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.eursta.01
Colombelli-Négrel, D., and C. Evans. 2017. Superb fairy-wrens respond more to alarm calls from mate and kin compared to unrelated individuals. Behavioral Ecology 28(4):1101-1112. https://doi.org/10.1093/beheco/arx071
Curio, E. 1978. The adaptive significance of avian mobbing: I. Teleonomic hypotheses and predictions. Zeitschrift für Tierpsychologie 48(2):175-183. https://doi.org/10.1111/j.1439-0310.1978.tb00254.x
da Cunha, F. C. R., J. C. R. Fontenelle, and M. Griesser. 2017a. Predation risk drives the expression of mobbing across bird species. Behavioral Ecology 28(6):1517-1523. https://doi.org/10.1093/beheco/arx111
da Cunha, F. C. R., J. C. R. Fontenelle, and M. Griesser. 2017b. The presence of conspecific females influences male-mobbing behavior. Behavioral Ecology and Sociobiology 71:52. https://doi.org/10.1007/s00265-017-2267-7
Eliassen, S., and C. Jørgensen. 2014. Extra-pair mating and evolution of cooperative neighbourhoods. Plos One 9(7):e99878. https://doi.org/10.1371/journal.pone.0099878
Feare, C. J. 1984. The starling. Oxford University Press, Oxford, UK.
Fisher, R. J., and K. L. Wiebe. 2006. Breeding dispersal of Northern Flickers Colaptes auratus in relation to natural nest predation and experimentally increased perception of predation risk. Ibis 148(4):772-781. https://doi.org/10.1111/j.1474-919X.2006.00582.x
Gibbs, H. L., P. J. Weatherhead, P. T. Boag, B. N. White, L. M. Tabak, and D. J. Hoysak. 1990. Realized reproductive success of polygynous red-winged blackbirds revealed by DNA markers. Science 250(4986):1394-1397. https://doi.org/10.1126/science.250.4986.1394
Grabowska-Zhang, A. M., B. C. Sheldon, and C. A. Hinde. 2012a. Long-term familiarity promotes joining in neighbour nest defence. Biology Letters 8:544-546. https://doi.org/10.1098/rsbl.2012.0183
Grabowska-Zhang, A. M., T. A. Wilkin, and B. C. Sheldon. 2012b. Effects of neighbor familiarity on reproductive success in the great tit (Parus major). Behavioral Ecology 23(2):322-333. https://doi.org/10.1093/beheco/arr189
Kessel, B. 1951. Criteria for sexing and aging European Starlings (Sturnus vulgaris). Bird-Banding 22(1):16-23. https://doi.org/10.2307/4510224
Kessel, B. 1957. A study of the breeding biology of the European Starling (Sturnus vulgaris L.) in North America. American Midland Naturalist 58(2):257-331. https://doi.org/10.2307/2422615
Kleindorfer, S., B. Fessl, and H. Hoi. 2005. Avian nest defence behaviour: assessment in relation to predator distance and type, and nest height. Animal Behaviour 69(2):307-313. https://doi.org/10.1016/j.anbehav.2004.06.003
Krama, T., A. Bērziņš, S. Rytkönen, M. J. Rantala, D. Wheatcroft, and I. Krams. 2012a. Linking anti-predator behaviour and habitat quality: group effect in nest defence of a passerine bird. Acta Ethologica 15:127-134. https://doi.org/10.1007/s10211-011-0117-6
Krama, T., J. Vrublevska, T. M. Freeberg, C. Kullberg, M. J. Rantala, and I. Krams. 2012b. You mob my owl, I’ll mob yours: birds play tit-for-tat game. Scientific Reports 2:800. https://doi.org/10.1038/srep00800
Krams, I., and T. Krama. 2002. Interspecific reciprocity explains mobbing behaviour of the breeding chaffinches, Fringilla coelebs. Proceedings of the Royal Society B 269:2345-2350. https://doi.org/10.1098/rspb.2002.2155
Krams, I., T. Krama, K. Igaune, and R. Mänd. 2008. Experimental evidence of reciprocal altruism in the pied flycatcher. Behavioral Ecology and Sociobiology 62:599-605. https://doi.org/10.1007/s00265-007-0484-1
Krams, I. A., A. Mennerat, T. Krama, R. Krams, P. Jõers, D. Elferts, S. Luoto, M. J. Rantala, and S. Eliassen. 2022. Extra-pair paternity explains cooperation in a bird species. Proceedings of the National Academy of Sciences 119(5):e2112004119. https://doi.org/10.1073/pnas.2112004119
Lima, S. L., and L. M. Dill. 1990. Behavioral decisions made under the risk of predation: a review and prospectus. Canadian Journal of Zoology 68(4):619-640. https://doi.org/10.1139/z90-092
Naďo, L., M. Kašová, A. Krištín, and P. Kaňuch. 2018. Cooperative nest-defence behaviour and territory quality in a resident and socially monogamous passerine. Ethology 124(7):514-526. https://doi.org/10.1111/eth.12755
Neudorf, D. L., and S. G. Sealy. 1992. Reactions of four passerine species to threats of predation and cowbird parasitism: enemy recognition or generalized responses? Behaviour 123:84-105. https://doi.org/10.1163/156853992X00138
Olendorf, R., T. Getty, and K. Scribner. 2004. Cooperative nest defence in red-winged blackbirds: reciprocal altruism, kinship or by-product mutualism? Proceedings of the Royal Society B 271:177-182. https://doi.org/10.1098/rspb.2003.2586
Pinxten, R., M. Eens, and R. F. Verheyen. 1989. Polygyny in the European Starling. Behaviour 111:234-256. https://doi.org/10.1163/156853989X00682
Pinxten, R., M. Eens, and R. F. Verheyen. 1991. Conspecific nest parasitism in the European Starling. Ardea 79:15-30.
Pinxten, R., M. Eens, and R. F. Verheyen. 1993a. Male and female nest attendance during incubation in the facultatively polygynous European Starling. Ardea 81:125-133.
Pinxten, R., O. Hanotte, M. Eens, R. F. Verheyen, A. A. Dhondt, and T. Burke. 1993b. Extra-pair paternity and intraspecific brood parasitism in the European Starling, Sturnus vulgaris: evidence from DNA fingerprinting. Animal Behaviour 45(4):795-809. https://doi.org/10.1006/anbe.1993.1093
R Core Team. 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Redmond, L. J., M. T. Murphy, A. C. Dolan, and K. Sexton. 2009. Parental investment theory and nest defense by Eastern Kingbirds. Wilson Journal of Ornithology 121(1):1-11. https://doi.org/10.1676/07-166.1
Riehl, C. 2013. Evolutionary routes to non-kin cooperative breeding in birds. Proceedings of the Royal Society B 280:20132245. https://doi.org/10.1098/rspb.2013.2245
Robinson, S. K. 1985. Coloniality in the Yellow-rumped Cacique (Cacicus cela) as a defense against nest predators. Auk 102(3):506-519. https://doi.org/10.1093/auk/102.3.506
Sandell, M. I., and M. Diemer. 1999. Intraspecific brood parasitism: a strategy for floating females in the European Starling. Animal Behaviour 57(1):197-202. https://doi.org/10.1006/anbe.1998.0936
Siderius, J. A. 1993. Nest defense in relation to nesting stage and response of parents to repeated model presentations in the Eastern Kingbird (Tyrannus tyrannus). Auk 110(4):921-923. https://www.jstor.org/stable/4088648
Sieving, K. E., and M. F. Wilson. 1998. Nest predation and avian species diversity in northwestern forest understory. Ecology 79(7):2391-2402. https://doi.org/10.1890/0012-9658(1998)079[2391:NPAASD]2.0.CO;2
Slade, J. 2012. Hackle spectral characteristics and their role in mate choice in European Starlings (Sturnus vulgaris). Thesis. Saint Mary’s University, Halifax, Canada. https://library2.smu.ca/handle/01/25213
Sproat, T. M., and G. Ritchison. 1993. The nest defense behavior of Eastern Screech-owls: effects of nest stage, sex, nest type and predator location. Condor 95(2):288-296. https://doi.org/10.2307/1369351
Stanback, M. T., N. A. DiLuzio, A. N. Mercadante, and E. S. Diamant. 2018. Eastern Bluebirds (Sialia sialis) do not abandon their chosen nest site in response to a single visit by a nest predator. Wilson Journal of Ornithology 130(2):568-573. https://doi.org/10.1676/17-027.1
Stutchbury, B. J., and R. J. Robertson. 1986. A simple trap for catching birds in nest boxes. Journal of Field Ornithology 57(1):64-65. https://sora.unm.edu/node/51244
Tkaczyk, S. P., D. P. Chivers, and K. L. Wiebe. 2022. Nest defense by Mountain Bluebirds (Sialia currucoides) is related to their sex and brood characteristics but not morphological attributes. Canadian Journal of Zoology 100(11):747-756. https://doi.org/10.1139/cjz-2022-0064
Trivers, R. L. 1971. The evolution of reciprocal altruism. Quarterly Review of Biology 46(1):35-57. https://doi.org/10.1086/406755
Vrublevska, J., T. Krama, M. J. Rantala, P. Mierauskas, T. M. Freeberg, and I. A. Krams. 2015. Personality and density affect nest defence and nest survival in the Great Tit. Acta Ethologica 18:111-120. https://doi.org/10.1007/s10211-014-0191-7
Westneat, D. F. 1993. Polygyny and extrapair fertilizations in eastern Red-winged Blackbirds (Agelaius phoeniceus). Behavioral Ecology 4(1):49-60. https://doi.org/10.1093/beheco/4.1.49
Fig. 1
Fig. 1. Map of study site with location of focal nest boxes on the campus of Saint Mary’s University in Halifax, Nova Scotia, Canada. Yellow highlighted circles illustrate the 16 nest boxes used.
Fig. 2

Fig. 2. Photo of the taxidermy mount of the American red squirrel used in the experimental trials.
Fig. 3

Fig. 3. Photo of the rock used in the control trials.
Fig. 4
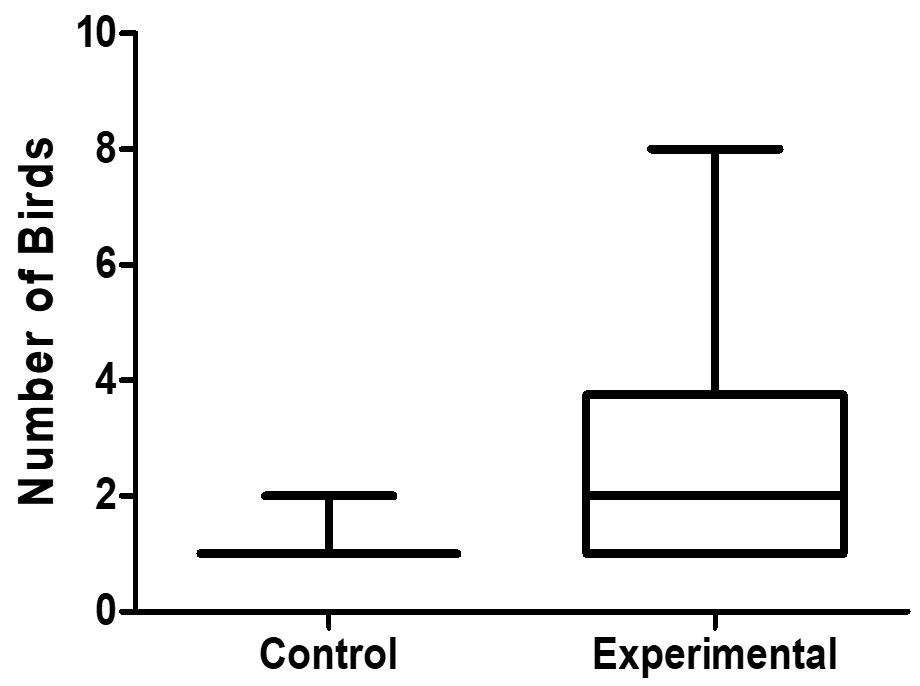
Fig. 4. Boxplots of the total number of European Starlings (parents and conspecifics) present at the focal nest box during each of the 16 control and 16 experimental trials.
Fig. 5
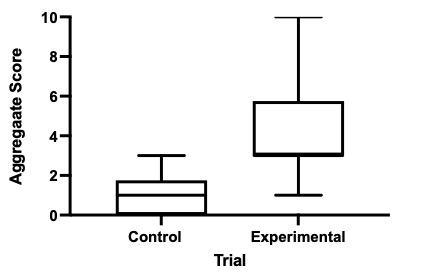
Fig. 5. Boxplots of the aggregate scores of nest defense by participating European Starlings during each of the 16 control and 16 experimental trials.
Table 1
Table 1. Range of response variables collected from European Starlings at their nest boxes when exposed to a rock (control treatment; N = 16 trials) or a taxidermied red squirrel (experimental treatment; N = 16 trials).
| Variable | Rock (control) | Red squirrel (experimental) | |||||||
| Number of starlings | 1–2 | 1–8 | |||||||
| Number of conspecifics (parents excluded) | 0 | 0–7 | |||||||
| Distance of approach (m) | 0–6 | 0–1.5 | |||||||
| Number of alarm calls | 0–86 | 62–262 | |||||||
| Number of chips | 0–72 | 0–45 | |||||||
| Number of fly-bys | 0–2 | 0–10 | |||||||
| Number of hits | 0 | 0–23 | |||||||
| Number of dives | 0 | 0–8 | |||||||