







The following is the established format for referencing this article:
Mejías, M. A., and D. R. Wilson. 2023. Breeding biology and nesting behavior of the endemic subspecies of White-eyed Vireo (Vireo griseus bermudianus) on the Bermuda archipelago. Journal of Field Ornithology 94(3):1.ABSTRACT
Avian island endemics are prone to extinction, and the preservation of remaining taxa requires long-term studies of their natural history and threats to survival. Thus, we provide, for the first time, a detailed account of the breeding biology and nesting threats of an endemic subspecies of vireonid, the non-migratory Bermuda White-eyed Vireo (Vireo griseus bermudianus). From 2016–2021, we located 84 nests, collected breeding data associated with 47 breeding pairs, and conducted observations throughout the nesting cycle. Some breeding pairs remained together for up to four successive breeding seasons. Bermuda Vireos began building nests in late February and cared for fledglings until mid-August. Both sexes contributed to all nesting stages, which were similar in duration to those of other vireo species (N, mean ± standard deviation): nest building (13 nests, 6 ± 3 d), incubation (11 nests, 14 ± 2 d), nestling care (6 nests, 11 ± 2 d), and fledgling care (5 nests, 41 ± 12 d). Nests, eggs, and nestlings were all vireonine in structure and appearance; nests were found in 14 tree species (2 endemic, 2 native, and 10 introduced). Across six years, 25 of the 42 nests (60%) with eggs produced nestlings, and 10 of the 25 broods (40%) produced fledglings; introduced predators were likely the primary cause of nest failure (16 of 27 nests, 59%). The Bermuda White-eyed Vireo is the last endemic terrestrial bird that breeds on the island and is of local conservation importance. Our observations should prove beneficial to other researchers interested in the breeding biology and nesting threats of breeding vireo species beyond Bermuda and for creating effective recovery plans necessary for conservation and management.
RESUMEN
Las especies de aves endémicas de las islas son propensas a la extinción y la preservación de los taxones restantes requieren de estudios de largo plazo sobre su historia natural y las amenazas a la supervivencia. Por lo tanto, aqui proporcionamos, por primera vez un recuento detallado de la biología reproductiva y las amenazas en la anidación de una subespecie endémica no migratoria de la familia Vireonidae, Vireo griseus bermudianus. Entre 2016-2021, localizamos 84 nidos, colectamos datos de reproducción asociados a 47 parejas reproductivas y realizamos observaciones a lo largo de su ciclo de anidación. Algunas parejas reproductivas se mantuvieron juntas hasta por cuatro temporadas reproductivas sucesivas. Vireo griseus bermudianus empezó a construir los nidos a finales de Febrero y cuidaron a sus volantones hasta mediados de Agosto. Los dos sexos contribuyeron en todas las etapas de la anidación, la cual fue similar en duración que la de otras especies de Vireo (N, media ± desviación estándar): construcción de nidos (13 nidos, 6 ± 3 d), incubación (11 nidos, 14 ± 2 d), cuidado de pichones (6 nidos, 11 ± 2 d), y cuidado de volantones (5 nidos, 41 ± 12 d). Los nidos, huevos y pichones fueron similares en apariencia y estructura a la de otras especies de la familia; los nidos fueron encontrados en 14 especies de árboles (2 endémicas, 2 nativas y 10 introducidas). A lo largo de seis años, 25 de los 42 nidos (60%) con huevos produjo pichones y 10 de las 25 nidadas (40%) produjo volantones; los depredadores introducidos fueron probablemente la primera causa de fracaso de los nidos (15 de los 27 nidos, 59%). Vireo griseus bermudianus es la última especie endémica terrestre que se reproduce en la isla y es de importancia para la conservación local. Nuestras observaciones pueden ser beneficiosas para otros investigadores más allá de Bermuda y para la creación de planes de recuperación necesarios para la conservación y el manejo.
INTRODUCTION
Since the early 1600s, approximately 90% of all bird extinctions have occurred on islands (Johnson and Stattersfield 1990). Humans have directly and indirectly driven these extinctions through: (1) introduction of predators and browsing animals, (2) harvesting birds and their eggs, and (3) habitat destruction (Wood et al. 2017). For example, predation from the introduced brown tree snake (Boiga irregularis) caused the extinction of the Guam Flycatcher (Myiagra freycineti; Savidge 1987); herbivory from introduced domestic rabbits (Oryctolagus cuniculus domesticus) led to the extinction of the Laysan Millerbird (Acrocephalus familiaris familiaris; Johnson and Stattersfield 1990); hunting led to the extinction of the Cuban Macaw (Ara tricolor; Wiley and Kirwan 2013); and tree clearing contributed to the extinction of the St. Kitts Bullfinch (Melopyrrha grandis; Olson 1984, Johnson and Stattersfield 1990). Compared to continental avifauna, island endemics are more prone to extinction because of naivety toward novel threats (Banks and Dickman 2007) and restricted ranges (Biber 2002).
According to the fossil record, avian endemism in recent millennia has been lower in Bermuda than in other oceanic islands and has continued to decline over time (Sterrer et al. 2004). Some birds in Bermuda went extinct during the Pleistocene, when episodic sea-level fluctuations caused significant land reductions (Olson and Wingate 2001, Olson and Hearty 2003, Olson 2008). Other endemic birds on the island became extinct because of human activity. For example, hogs (Sus scrofa) introduced to Bermuda in the mid-1500s devastated breeding bird populations (Sterrer et al. 2004, Olson and Wingate 2006, Olson and Wingate 2012, Olson 2013). The early settlers, who arrived in 1609, consumed birds and their eggs (Lefroy 1877) and indirectly reduced bird numbers by introducing mammalian, avian, and insect predators (Lefroy 1877, Sterrer et al. 2004, Mejías et al. 2017). Extinct Bermuda endemics include a night heron (Nyctanassa carcinocatactes; Olson and Wingate 2006), a towhee (Pipilo naufragus; Olson and Wingate 2012), and the Bermuda Flicker (Colaptes oceanicus; Olson 2013). Centuries of tree felling and exotic plant introductions have also replaced most of Bermuda’s native forest (Wingate 1990, Sterrer et al. 2004), removing the precolonial flora in which endemic birds evolved.
Since the 2000s, only 14 resident landbird species breed on Bermuda, including the Bermuda White-eyed Vireo (Vireo griseus bermudianus; hereafter Bermuda Vireo). This vireonid is a small (9–15 g), non-migratory, endemic subspecies of White-eyed Vireo that commonly travels in pairs throughout the woodlands, thickets, and marshes of Bermuda (Bangs and Bradlee 1901, Amos 1991). Males are easily identified by their complex song known onomatopoeically as “chick-of-the-village.” The song, which gives the bird its local nickname, is used for mate attraction and year-round territory defense (Mejías et al. 2020, 2021, Mejías 2021). No official population estimate is available for the Bermuda Vireo, although historical and present-day observations suggest a large, island-wide population (Jones 1859, Amos 1991, Mejías and Nol 2020). Despite its widespread presence in wooded habitats across the island, conspicuous vocal displays, and tame nature, the Bermuda Vireo’s breeding biology remains unknown. Given the frequent fate of island endemics and paucity of data on nests and nesting behavior of Bermuda Vireos, our objectives were to provide fundamental information on the Bermuda Vireo’s nesting biology, including descriptions of: (1) breeding success and causes of reproductive failure; (2) pair bond duration; (3) breeding phenology and behavior; and (4) nests, eggs, and nestlings. This information provides a necessary foundation for conservation and management of the Bermuda Vireo and may provide insight into the breeding biology of other vireos.
METHODS
Study site
Bermuda is an island (32°18’ N, 64°47’ W) formed from fossilized, aeolian shell-sand (Fig. 1). It is a low-lying, hilly (0–76 m, mean: 38 m) archipelago with a subtropical climate; sunshine and light winds dominate April–September, whereas rain and gales are common in winter (Amos 1991). The average daily temperatures across the year range from 18–27.5ºC. Present-day wooded habitat is dominated by secondary forests of introduced vegetation, including Brazilian peppertree (Schinus terebinthifolia), Australian whistling pine (Casuarina equistetifolia), Chinese fan palm (Livistona chinensis), allspice (Pimenta dioica), and Suriname cherry (Eugenia uniflora). Although uncommon, some native trees remain, including Bermuda cedar, Bermuda palmetto (Sabal bermudana), Bermuda olivewood (Elaeodendron laneanum), and southern hackberry (Celtis laevigata).
Field methods
We observed breeding Bermuda Vireos (Fig. 2) across the main island and offshore islets from February to September 2016–2021, although pair bond data were collected until November 2021. We collected vireo nesting data as follows. Nesting observations in 2016 (N = 2 nests), 2020 (N = 2 nests), and 2021 (N = 5 nests) were based on chance encounters. In 2017, observations (N = 30 nests) were derived from intentional nest searches. In 2018–2019, observations (N = 45 nests) were collected opportunistically while studying Bermuda Vireos in the context of another study focused on singing behavior (Mejías et al. 2021).
We visited territories on days with little to no wind or rain, between 07:00 and 17:00. We found Bermuda Vireos by listening for their song or scolding calls (Mejías et al. 2021) and then followed them from a minimum distance of 5 m as they carried materials to their nest. During years of haphazard nest encounters, we also found nests directly along trails or in the foliage.
We obtained detailed breeding data from some nests by watching the birds with binoculars for 30–60 min/d, every 1–4 days, from a standard location concealed by vegetation at least 5 m away from the nest. Like many island birds, Bermuda Vireos are usually tolerant of humans, and our presence at this distance did not trigger scolding calls indicative of agitation (also known as “chatter vocalizations” [Bradley 1980] or “alarm chatter” [Hopp 2022]). We transcribed nest activities in field diaries, and, during some nest watches, monitored nests to note whenever a vireo added nest material, relieved its mate from incubation duties, or brought food to nestlings. The sex of each parent was determined by song, given that only males sing the species’ typical “discrete song” (Bradley 1980).
Breeding success and nest threats
We determined clutch size for 21 nests that were accessible by looking inside nests and counting eggs and/or nestlings when the parents were absent. For estimates of breeding success, we report the percentage of nests where at least one egg produced a hatchling, and the percentage of nests where at least one egg produced a fledgling.
We identified the cause of nest failure whenever possible. The black rat (Rattus rattus), Great Kiskadee (Pitangus sulphuratus), and Argentine ant (Linepithema humile), which are introduced species and known predators of Bermuda Vireo eggs and nestlings (Mejías 2021), were abundant across all study sites. Ant predation was identifiable by the tendency of swarms to cover nests and slowly devour eggs or nestlings. Predation from Great Kiskadees and rats, however, could not be confirmed because we did not have a trail camera; thus, the sudden disappearance of nest contents was attributed to an unknown predator. Lastly, some nests collapsed at various nesting stages. Collapse was characterized by unravelling of the nest rim from forked branches, outer wall material sloughing off, or the nest splitting in half. Nest collapse was a gradual process that usually occurred over multiple nest watches and sometimes resulted in the eggs or nestlings falling out.
After fledging, nest abandonment, or egg or nestling failure, we used a tape measure to measure the distance from the bottom of the nest to the ground. For nests that were too high to reach, we visually estimated nest height to the nearest 1 m. For descriptive purposes, we identified, whenever possible, the tree species containing the nest, collected the nests once breeding activity ceased, and identified lichen, plant, and anthropogenic materials composing the nest. We used a ruler to measure the inner nest depth (accuracy: ± 1 mm), and vernier calipers to measure (± 1 mm) nest length (i.e., from nest rim to nest base) and the minimum and maximum external nest diameters at the top of the nest. If abandoned eggs were present or if the parents were away from an active nest on an incubation break, we also measured the maximum length and width of eggs.
Pair bonds
We estimated the duration of pair bonds by counting the number of days that breeding pairs remained together inside a territory over the duration of the study period. We calculated pair duration based exclusively on vireos in which both members had been color-banded in the context of previous research; our estimates are conservative because some pairs formed before being banded, and others remained together after the study period ended. We considered a pair bond to have ended if one of the color-banded birds was no longer seen travelling or engaging in nesting behavior with its previously known mate or was no longer associated with the previous mate’s territory.
Breeding phenology
We estimated the phenology and duration of the following breeding stages: (1) nest building, (2) incubation, (3) nestling care, and (4) fledgling care. Nest building is the period when vireos are adding materials to the nest. We estimated the duration of the nest building period using nests (N = 17) that we were confident we had found on the first day of construction (e.g., there were only a few bits of material on a branch). Incubation is the period when parents tended to a clutch. Because we did not approach and inspect every nest, we inferred incubation whenever a parent vireo was seen sitting in the nest throughout most (≥ 70%) of a nest watch. Following Lapergola et al. (2012), we assumed that incubation began immediately after clutch completion. If we were uncertain about when a clutch was completed, as determined by counting eggs during consecutive, opportunistic viewings of unattended nests, we still inferred incubation behavior but did not include the nest in our estimate of the duration of the incubation stage. Nestling care is when parents feed chicks inside the nest, and fledgling care is when parents feed the young outside the nest. We presumed nestling care when we observed parents bringing food to the nest or when the parents perched on the nest rim and looked inside the nest cup. We inferred fledgling care when either parent was seen feeding young that were perched in the foliage. Begging calls helped us to follow the fledglings’ movements, though the number of calls gradually declined as the young gained independence. We considered fledgling care to have ended when young were capturing their own food in natal territories and were no longer seen being fed by either parent. Because we monitored nests and fledglings every 1–4 days, we estimated the transition between any two stages as the midpoint between the last day observed in the previous stage and the first day observed in the subsequent stage.
RESULTS
Breeding success and nest threats
Of 42 clutches, 25 (60%) produced at least one hatchling, and 10 of those 25 (40%) produced at least one fledgling. Breeding success rate (percentage of nests with eggs that fledged at least one chick) was 24% across the years. Bermuda Vireos produced one brood per breeding season, although re-nesting was common after nest failure. Following a nest failure, breeding pairs commenced construction of a new nest within 1–22 days (6 ± 5 days, mean ± standard deviation [SD]; N = 30 nests). Pairs always laid replacement clutches in a new nest.
We identified four causes of nest failure among 27 nests with eggs or nestlings: predation (N = 16, 59%), nest collapse (7, 26%), nest abandonment (3, 11%), and nestlings falling out (1, 4%). The Argentine ant was responsible for 5 (33%) predation events involving eggs and nestlings, as evidenced by ant swarms covering the nest, eggs, or dead nestlings. Kiskadees and rats were likely responsible for predation events for which predators were unconfirmed. We saw a Great Kiskadee divebomb a brooding vireo with newly hatched chicks, and this nest was found destroyed approximately 3 days later. Similarly, we often saw rats directly beneath or above vireo nests a few days before naked nestlings disappeared. Throughout all of these stages, adults defended nests, eggs, and nestlings from both animal and human intruders with scolding calls, active chasing, and divebombing.
Pair bonds
Across the study period, we documented breeding behavior in 81 individuals from 47 breeding pairs. Among the 43 vireos that were banded (Fig. 2), 26 were male and 17 were female; together, these individuals formed 18 pairs (one female was involved in four pairs; Female: RWG, Table 1) in which both partners were banded. Our sample also included 12 breeding pairs in which both members were unbanded. Unbanded pairs could not be used to infer pair duration, but they provided data on breeding behavior, nesting success, and nest predation.
Our observations of color-banded birds revealed that male-female pairs remain together and defend the same territories year-round, although pair bonds can end at any time (Mejías 2021). Paired vireos often travelled together while giving “fit-fit-fit” calls and engaged in nesting activity with one another inside their territory. Pair duration varied substantially among 18 color-banded pairs, with some pairs seen together for only 1 day and others for up to 1360 days (Table 1). Among these 18 nesting pairs, 7 made multiple breeding attempts, and 1 bred across multiple breeding seasons (Male: BBP/; Female: BWG/, 24 May 2017–11 February 2021; Table 1). Based on a subset of color-banded pairs in 2019, the number of nest attempts by a given pair within a single season ranged from 1–4 (3 ± 1, mean ± SD; N = 6). Following the end of pair bonds, newly unpaired males typically remained inside their year-round territories and increased their singing rate, whereas the newly unpaired females dispersed, sometimes visiting the territories of several neighboring males. We commonly observed pair bonds ending after one or more nest failures. In one case, a pair bond ended after the disappearance of a color-banded male in December 2019; the female remained in the same territory and was joined by a new, unbanded male. However, in May 2020, this same female was building a nest with a different color-banded neighboring male, apparently having ended her pair bond with the unbanded male.
Breeding phenology
We found 84 nests and obtained data on phenology for 60 (71%) of them. Information on the number of nests reaching each breeding stage, and the fate of the nests, is summarized in Table 2.
We observed nest building from 28 February–24 June, but most pairs built nests in April (Fig. 3A). We recorded nest building behavior for 22 pairs and 46 nests. The mean (± SD) duration of nest building, based on 13 nests for which it was observed in its entirety, was 6 ± 3 d (range: 2–12 d). Males often secured the first piece of nest material on a horizontal, forked branch. Pairs would then go on collecting trips together within the vicinity of the nest-site, both returning to the building site with material. Nests changed from being a clump of plant bits or polyfill stuffing on a forked branch to a shallow sling of plant and litter fibers, and finally, to a thick, basket-like structure (Fig. 4) that supported adults, eggs, and nestlings (Figs. 4 and 5). The outer nest wall contained lichen (Ramalina denticulata) and bark and leaves from Bermuda palmetto, allspice, and cow cane (Arundo donax); polyfill stuffing was the most common kind of human garbage in nests. Adults lined all nests with reddish-brown straw fibers from old leaf bases of the Bermuda palmetto and Chinese fan palm. We confirmed 14 nest tree species (N = 79 nests), including endemic or native (12 of 79 nests, 15%) and introduced (67 nests, 85%) species. Nest measurements (mean ± SD, range, N) were as follows: nest height (3 ± 1 m, 1–7 m, 63 nests); nest length (71 ± 13 mm, 49–116 mm, 34 nests); nest depth (46 ± 6 mm, 32–64 mm, 34 nests); widest outside nest diameter (76 ± 8 mm; 60–91 mm, 34 nests); narrowest outside nest diameter (68 ± 8 mm, 53–85 mm, 34 nests). Based on a subset of 25 bouts of nest-building from 15 different nests, we recorded 99 nest visits by building pairs. On average, building pairs returned from collecting trips after 11 ± 8 min (1–31 min). Once the nest structure was ~75% complete, males stopped contributing to nest construction and instead shadowed the female on her collecting trips, flicking his wings in front of the female while she finished the nest. A hiatus usually occurred between nest completion and the onset of egg laying (4±3 d; 1–12 d); the hiatus was not included in the nest construction or incubation duration calculations.
We observed vireos in nests from 31 March–5 July, with incubation peaking in May and June (Fig. 3B). The mean (± SD) duration of incubation, based on 11 nests for which the incubation stage was observed in its entirety, was 14 ± 2 d (range: 12–16 d). During this stage, we observed 25 pairs and 44 nests. While inside the nest, vireos sat motionless, with only the head, bill, and tail visible above the nest rim (Fig. 5). Rarely, males sang discrete songs with noticeably longer inter-song pauses while sitting inside the nest. Soon after, the female returned to the nest and relieved the male, which then resumed steady discrete song bouts away from the nest. Based on a subset of 51 bouts of incubation from 16 different nests, the mean (± SD) duration of an incubation bout was 30±14 min (5–60 min). We observed 24 changeovers, when the incubating bird exited and its mate entered the nest. Changeovers ranged from 5–53 min (22 ± 12 min). Eggs were pearly white with reddish-brown blotches concentrated on the blunt end (Fig. 4). Clutch size varied among nesting pairs (3 ± 1 egg, 1–4 eggs, 21 nests; Fig. 5). Egg measurements were as follows: egg length (19 ± 1 mm, 17–20 mm, 19 eggs), and egg width (14 ± 1 mm, 13–16 mm, 19 eggs).
We observed nestling care from 21 April–16 July, with nestling care peaking in May (Fig. 3C). The mean (± SD) duration of nestling care, based on six nests where it was observed in its entirety, was 11 ± 2 d (range: 9–14 d). Observations from a single brood of three nestlings in 2017 revealed that nestlings were altricial at hatching, with rapid growth, feathering, and eye-opening occurring over 9 d (Fig. 6). We monitored 16 pairs of vireos and 20 nests with nestlings. During this period, both sexes carried food in their bills, including insects, caterpillars, spiders, and Anolis lizards. After delivering food to the nest, parents removed white nestling fecal sacs from the nest and consumed them at the nest or dropped them several meters away. During the first 3–4 days of nestling care, at least one parent remained at the nest, either perched on the nest rim looking inside, or brooding the naked young; the mate was usually away collecting food. As nestlings aged, both parents went independently on foraging trips. Among a subset of 19 nestling feeding bouts, we recorded breeding pairs returning to nests to feed young every 14 ± 12 min (3–43 min), based on 113 individual nestling feedings trips from seven different nests.
We observed fledglings being reared from 30 April–11 August; fledgling care peaked in May and June (Fig. 3D). The mean (± SD) duration of fledgling care, based on five families for which it was observed in its entirety, was 41 ± 12 d (range: 26–57 d). We followed five pairs of vireos caring for five broods of fledglings. Throughout this stage, adult vireos gleaned food from the canopy foliage and branches while constantly being followed by their young, which trailed close behind while emitting begging calls and vibrating their wings. Despite being paired, the two parents ceased traveling together during fledgling feeding. Rather, observations from all color-banded pairs revealed that the parents split the brood during this period. Males, which used conspicuous, discrete song to guide fledglings, reared young inside their own territory, whereas females, who do not sing, attended the young either in or outside the natal territory; if the latter, it was usually done briefly inside neighboring territories. Parental aggression toward perceived threats such as human observers intensified after fledging. Females seemed to be more responsive to our presence and gave bouts of scolding calls when we were present anywhere in the natal territory. In contrast, males usually did not react defensively to our presence. In response to their parents’ scolding calls, fledglings ceased begging calls and remained motionless in the foliage; older offspring sometimes helped defend natal territories with scolding calls. Banding data from another study (Mejías and Watson, unpublished data) revealed that fledglings eventually disperse from natal territories at some point after parents stop feeding them, whereas parents remain inside their territory, with some parents even chasing their independent fledglings around natal territories (M. A. Mejías, personal observation).
DISCUSSION
This is the first detailed account of the nesting behavior of Vireo griseus bermudianus. The Bermuda Vireo nests in several tree species and has a breeding season that begins with nest construction in late February. Fledgling care extends into early August, due in part to multiple nest attempts following predation by introduced predators. Our observations reveal that Bermuda Vireo nesting habits are largely congruent with the Vireo genus.
Pair bond duration varied widely. Some pairs were together for less than one year and others for multiple years, although we confirmed multiyear breeding by only one banded pair. While data on vireonid pair duration is scant, Morton et al. (2010) observed several within-year divorces among Blue-headed Vireos (Vireo solitarius), with no indication of pairs lasting multiple seasons. Black-capped Vireos (V. atricapilla) have longer pair durations, with some surviving three consecutive seasons (Graber 1961). Our maximum pair duration of 4 yr was higher than those reported for either species. Pair duration might be more stable in Bermuda than in North America because Bermuda Vireos are non-migratory; thus, these pairs never leave territories vacant or risk mortality with long-distance movements. Interestingly, all recorded divorces in Blue-headed Vireos involved females disappearing from their mate’s territories (Morton et al. 2010), which is similar to our observations of Bermuda Vireos, for which the end of pair bonds involved males remaining in their territories and females departing and visiting nearby unpaired males in neighboring territories. During the 2017 breeding season, one female Bermuda Vireo was involved in sequentially monogamous pair bonds with four neighboring males from February to May; female Blue-headed Vireos also behave in this manner (Morton et al. 2010). Sexual selection (Andersson 1994) may favor a vireonid breeding system in which it is costly for males, as the competing sex, to relinquish their territories and associated resources, but beneficial for females, as the choosy sex, to desert.
Vireo breeding phenology varies across species, although most studies are biased toward temperate migrants breeding at more northerly latitudes. Temperate vireos have breeding seasons that generally span April–August (e.g., Bell’s Vireo, V. bellii [Nolan 1960]; Blue-headed Vireo [Marvil and Cruz 1989]; Red-eyed Vireo, V. olivaceus [Robinson 1981]; Philadelphia Vireo, V. philadelphicus [Robinson 1981]; Plumbeous Vireo, V. plumbeus [DeMarco et al. 2000]; and Cassin’s Vireo, V. cassinii [Hedley 2019]), whereas in our study, Bermuda Vireos began nest building in late February and cared for fledglings into mid-August; bermudianus fledglings are sometimes reared by their parents as late as early September (M. A. Mejías, personal observation). Despite differences in breeding phenology and parental care among Vireo species, the genus nevertheless shows overall conservatism in duration of each breeding stage at the individual level: nest building (5–8 d; Graber 1961, James 1997), incubation (14–17 d; Gómez-Montes and Moreno 2008, Morton et al. 2010), and nestling care (12–13 d; Graber 1961, James 1999). Data on the duration of fledgling care are unavailable for most vireonids. Fledgling care lasts ~28 d in San Andreas Vireo (V. caribaeus; Gómez-Montes and Moreno 2008), ~35–45 d in Black-capped Vireo (Graber 1961, Grzybowski 2020), and 26–57 d in Bermuda Vireo (this study). Interestingly, Winkler et al. (2020) noted the tendency of female Black-capped Vireos to rear their fledglings beyond natal territories, with the males largely feeding in their own territories, as we found for Bermuda Vireos. Finally, at least two patterns of parental contribution occur in Vireo species. In one, males do not assist with nest building or incubation, but help with nestling care (Rust 1920, Barlow and Rice 1977). In the other, both sexes contribute to all phases of the breeding cycle (Ritchison et al. 2019); the Bermuda Vireo belongs to the second group.
Bermuda Vireo nests were generally vireonine in structure: an open pensile cup made of fibrous strips of plant, lichen, and garbage, suspended from a horizontal forked branch by “basket handles” (Winkler et al. 2020). Despite all Vireo nests being cup-shaped, the outer wall and nest lining reflect the respective habitat being used (Lapergola et al. 2012, Mejías et al. 2020, Winkler et al. 2020). Bermuda Vireos used strips of bark and palm fronds, mosses, lichen (Ramalina denticulata), and garbage bits (especially polyfill stuffing) to form the outside wall, and lined nests with reddish-brown straw fibers from old leaf bases of either Bermuda palmetto or Chinese fan palm. An historical account by Reid (1877:14) states that their “pretty pensile nest” generally hangs 0.9–3.7 m above the ground in native mangroves and Bermuda cedar. In the 21st century, they build their nests in 14 tree species, most of which are introduced; this is typical of their generalist nature (Mejías and Nol 2020). A similar diversity of 11 tree species was used by a mainland population of White-eyed Vireos (Peake and Ritchison 1998). On average, Bermuda Vireos built their nests at a height of 2.5 m above the ground, whereas the mean height of nests in continental, White-eyed Vireos is 0.8–1.5 m (Peake and Ritchison 1998, Conkling 2010). Bermuda Vireos are allopatric with all other vireos and thus do not experience congeneric competition for nest sites (Mejías et al. 2021), which might explain why bermudianus selects slightly higher branches for nest placement than continental griseus.
Clutch size and chick growth also were similar to other vireonids. Crowell and Rothstein (1981) reported that Bermuda Vireos produce an average clutch size of 2 eggs compared to 3-egg clutches in the North American race; our larger sample size shows that clutch size in Bermuda Vireos is similar to that of continental V. griseus (2–4 eggs, average of 3 eggs; Ritchison et al. 2019). Like other vireo species, Bermuda Vireo nestlings are naked at hatching, with the first feather sheaths appearing by day 3, feathers bursting around day 6, and nestlings fully feathered by day 9, just prior to fledging (Winkler et al. 2020).
Most nesting attempts in Bermuda were unsuccessful. Nesting Bermuda Vireos made up to four nest attempts, whereas Puerto Rican Vireos (Vireo latimeri) made up to six attempts (Faaborg et al. 1997); such data appear unavailable for other vireonids. In Bermuda, only 50% (42 of 84) of nests that were initiated led to eggs being produced. Nest failure was common among the 42 nests containing eggs or nestlings. Although we often did not witness predation directly, introduced predators were observed near several nests immediately before or during nest failure, suggesting that predation from introduced predators (16 of 42 nests) is the most common cause of nest failure. Nest collapse (7 of 42 nests) was the second most common cause of nest failure. Low breeding success is common among mainland and island vireos (Graber 1961, Siepielski et al. 2001, Tossas 2008, Kovar et al. 2018, Ritchison et al. 2019). Brood parasitism from the native Brown-headed Cowbird (Molothrus ater) on the mainland (Pitelka and Koestner 1942, Graber 1961, Marvil and Cruz 1989) and from introduced Shiny Cowbirds (M. bonariensis) in the Caribbean (Wiley 1985, Woodworth 1995) is the primary cause of nesting failure in said populations. Neither of these brood parasites are present on Bermuda during the breeding season (Amos 1991). Nest collapse in Puerto Rican Vireos was attributed to the additional weight of cowbird nestlings (Woodworth 1997). We hypothesize that nest collapse in Bermuda was due to a lack of sufficient nest binding material, specifically, the durable silk of Golden Silk Orb-Weavers (Trichonephila clavipes) that was historically a common component of Bermuda Vireo nests (D. B. Wingate, personal communication). We found no evidence of golden silk in any of the nests we observed, possibly because this arachnid is no longer very abundant.
The Bermuda subspecies of White-eyed Vireo is of significant, local conservation importance because it is the last endemic terrestrial bird that breeds on the archipelago. Presently, introduced predators are likely the primary cause of nest failure. Although many bird species on the island, especially ground birds, are killed by cats (M. A. Mejías, personal observation), we are unaware of any record of cat predation on the Bermuda Vireo, probably because this subspecies seldom leaves the thickets. Despite nest predation and nest collapse, the Bermuda Vireo’s ability to live in introduced vegetation (Mejías and Nol 2020), to use diverse materials for constructing nests, and to make multiple breeding attempts may offset its poor nesting success. Future studies should use Bermuda Christmas Bird Count data (National Audubon Society 2020), comprising 74 years of single-day island-wide bird surveys, to estimate population trends of the Bermuda Vireo. A long-term mark-recapture study would also be useful in determining survival and recruitment of this subspecies. Mounted cameras should be used to confirm the identity of nest predators. Lastly, research can test the hypothesis that nest quality is dependent on the use of spider silk from T. clavipes, with the prediction that nests with the golden silk are less likely to collapse than those without the silk.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
M. A. M. conceived the manuscript idea, carried out fieldwork, collected and analyzed the data, and wrote the paper. D. R. W. provided funding that supported field research and gave substantial feedback that greatly improved the manuscript. Both authors reviewed and approved the finalized version of the manuscript.
ACKNOWLEDGMENTS
We are grateful to P. Watson for teaching M. Mejías how to capture, handle, and band songbirds, and to M. Outerbridge, who issued Bermuda fieldwork permits. Many thanks to P. Mejías and A. Mejías for their support and invaluable field assistance. We extend our thanks to members of the public who alerted us to the presence of vireo nests. Lastly, we thank T. Miller, J. Roncal, D. Innes, P. Martin, and I. Warkentin for providing feedback on an earlier version of this manuscript.
DATA AVAILABILITY
The dataset associated with this paper is openly available in Dryad at https://doi.org/10.5061/dryad.bvq83bkfs. Ethical approval for this research study was granted by a research permit issued by the Bermuda Department of Environment and Natural Resources (License 17-02-02-59), and was approved by the Animal Care Committee of Memorial University (Animal Use Protocols 15-02-DW and 18-02-DW).
LITERATURE CITED
Amos, E. J. R. 1991. A guide to the birds of Bermuda. Corncrake, Warwick, Bermuda.
Andersson, M. 1994. Sexual selection. Princeton University Press, Princeton, New Jersey, USA.
Bangs, O., and T. S. Bradlee. 1901. The resident land birds of Bermuda. Auk 18(3):249-257. https://www.jstor.org/stable/4069401
Banks, P. B., and C. R. Dickman. 2007. Alien predation and the effects of multiple levels of prey naiveté. Trends in Ecology and Evolution 22(5):229-230. https://doi.org/10.1016/j.tree.2007.02.006
Barlow, J. C., and J. C. Rice. 1977. Aspects of the comparative behavior of Red-eyed and Philadelphia Vireos. Canadian Journal of Zoology 55(3):528-542. https://doi.org/10.1139/z77-068
Biber, E. 2002. Patterns of endemic extinctions among island bird species. Ecography 25(6):661-676. https://doi.org/10.1034/j.1600-0587.2002.t01-1-250603.x
Bradley, R. A. 1980. Vocal and territorial behavior in the White-eyed Vireo. Wilson Bulletin 92(3):302-311. https://www.jstor.org/stable/4161354
Conkling, T. J. 2010. Analysis of the Black-capped Vireo and White-eyed Vireo nest predator assemblages. Thesis. Texas A&M University, College Station, Texas, USA. https://nri.tamu.edu/publications/theses-dissertations/2010/analysis-of-the-black-capped-vireo-and-white-eyed-vireo-nest-predator-assemblages/
Crowell, K. L., and S. I. Rothstein. 1981. Clutch sizes and breeding strategies among Bermudan and North American passerines. Ibis 123(1):42-50. https://doi.org/10.1111/j.1474-919X.1981.tb00171.x
DeMarco, T. E., C. B. Goguen, D. R. Curson, and N. E. Mathews. 2000. Breeding behavior of the Plumbeous Vireo in New Mexico. Western North American Naturalist 60(4):394-402. https://www.jstor.org/stable/41717056
Faaborg, J., K. M. Dugger, W. J. Arendt, B. L. Woodworth, and M. E. Baltz. 1997. Population declines of the Puerto Rican Vireo in Guánica forest. Wilson Bulletin 109(2):195-202. https://www.jstor.org/stable/4163802
Gómez-Montes, C., and M. I. Moreno. 2008. Breeding phenology and nesting habitat characterisation of the San Andres Vireo (Vireo caribaeus). Bird Conservation International 18(4):319-330. https://doi.org/10.1017/S0959270908007429
Graber, J. W. 1961. Distribution, habitat requirements, and life history of the Black-capped Vireo (Vireo atricapilla). Ecological Monographs 31(4):313-336. https://doi.org/10.2307/1950756
Grzybowski, J. A. 2020. Black-capped vireo (Vireo atricapilla). Version 1.0. In A. F. Poole and F. B. Gill, editors. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.bkcvir1.01
Hedley, R. W. 2019. Long-distance movements and evidence of post-breeding elevational movements by Cassin’s Vireos. Journal of Field Ornithology 90(4):335-347. https://doi.org/10.1111/jofo.12309
Hopp, S. L. 2022. White-eyed Vireo (Vireo griseus). Version 2.0. In P. G. Rodewald and B. K. Keeney, editors. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.whevir.02
James, R. D. 1997. Yellow-throated and Solitary Vireos in Ontario: 3. Nest building. Ontario Birds 15(1):14-20. http://www.ofo.ca/library/view/id/104
James, R. D. 1999. Yellow-throated and Blue-headed Vireos in Ontario: 5. Nestling period, and post-nesting activities. Ontario Birds 17(1):14-21. http://www.ofo.ca/library/view/id/110
Johnson, T. H., and A. J. Stattersfield. 1990. A global review of island endemic birds. Ibis 132(2):167-180. https://doi.org/10.1111/j.1474-919X.1990.tb01036.x
Jones, J. M. 1859. The naturalist in Bermuda: a sketch of the geology, zoology, and botany of that remarkable group of islands; together with meteorological observations. Reeves & Turner, London, UK. https://doi.org/10.5962/bhl.title.57198
Kovar, D. G., J. H. Sperry, and D. A. Cimprich. 2018. Comparative reproductive ecology of co-occurring rare and common vireos in shrublands and woodlands. Wilson Journal of Ornithology 130(2):410-426. https://doi.org/10.1676/16-224.1
Lapergola, J. B., J. G. Marina Hipolito, J. E. Martínez-Gómez, and R. L. Curry. 2012. First description of the nest and eggs of the island-endemic Cozumel Vireo, Vireo bairdi. Wilson Journal of Ornithology 124(4):743-749. https://doi.org/10.1676/1559-4491-124.4.743
Lefroy, J. H. 1877. Memorials of the discovery and early settlement of the Bermudas or Somers Islands, 1515–1685. Volume I. Reprinted by Eyre and Spottiswoode, London, UK.
Marvil, R. E., and A. Cruz. 1989. Impact of Brown-headed Cowbird parasitism on the reproductive success of the Solitary Vireo. Auk 106(3):476-480. https://www.jstor.org/stable/4087871
Mejías, M. A. 2021. Management plan for the Bermuda White-eyed Vireo (Vireo griseus bermudianus). Department of Environment and Natural Resources, Government of Bermuda, Hamilton, Bermuda. https://environment.bm/species-recovery-plans
Mejías, M. A., and E. Nol. 2020. Woodland size and vegetation effects on resident and non-resident woodland birds in Bermuda. Journal of Caribbean Ornithology 33:22-32. https://doi.org/10.55431/jco.2020.33.22-32
Mejías, M. A., J. Roncal, T. S. Imfeld, S. Boisen, and D. R. Wilson. 2020. Relationships of song structure to phylogenetic history, habitat, and morphology in the vireos, greenlets, and allies (Passeriformes: Vireonidae). Evolution 74(11):2494-2511. https://doi.org/10.1111/evo.14099
Mejías, M. A., J. Roncal, and D. R. Wilson. 2021. Territorial responses of Bermuda White-eyed Vireos (Vireo griseus subsp. bermudianus) reflect phylogenetic similarity of intruders and acoustic similarity of their songs. Journal of Field Ornithology 92(4):431-449. https://doi.org/10.1111/jofo.12384
Mejías, M. A., D. B. Wingate, J. L. Madeiros, Y. F. Wiersma, and G. J. Robertson. 2017. Nest-cavity selection and nesting success of Bermudian White-tailed Tropicbirds (Phaethon lepturus catesbyi). Wilson Journal of Ornithology 129(3):586-599. https://doi.org/10.1676/16-115.1
Morton, E. S., B. J. M. Stutchbury, and I. Chiver. 2010. Parental conflict and brood desertion by females in blue-headed vireos. Behavioral Ecology and Sociobiology 64:947-954. https://doi.org/10.1007/s00265-010-0910-7
National Audubon Society. 2020. Christmas bird count historical results. National Audubon Society, New York, New York, USA. http://www.christmasbirdcount.org
Nolan, V. Jr. 1960. Breeding behavior of the Bell Vireo in southern Indiana. Condor 62(4):225-244. https://doi.org/10.2307/1365515
Olson, S. L. 1984. The last St. Kitts Bullfinch Loxigilla portoricensis grandis (Emberizinae) and the extinction of its race. Bulletin of the British Ornithologists’ Club 104(4):121-124. https://repository.si.edu/bitstream/handle/10088/12777/VZ_154_Last_St_Kitts_Bullfinch.pdf?sequence=1&isAllowed=y
Olson, S. L. 2008. A new species of shearwater of the genus Calonectris (Aves: Procellariidae) from a middle Pleistocene deposit on Bermuda. Proceedings of the Biological Society of Washington 121(3):398-409. https://doi.org/10.2988/08-11.1
Olson, S. L. 2013. Fossil woodpeckers from Bermuda with the description of a new species of Colaptes (Aves: Picidae). Proceedings of the Biological Society of Washington 126(1):17-24. https://doi.org/10.2988/0006-324X-126.1.17
Olson, S. L., and P. J. Hearty. 2003. Probable extirpation of a breeding colony of Short-tailed Albatross (Phoebastria albatrus) on Bermuda by Pleistocene sea-level rise. Proceedings of the National Academy of Sciences 100(22):12825-12829. https://doi.org/10.1073/pnas.1934576100
Olson, S. L., and D. B. Wingate. 2001. A new species of large flightless rail of the Rallus longirostris/elegans complex (Aves: Rallidae) from the late Pleistocene of Bermuda. Proceedings of the Biological Society of Washington 114(2):509-516. https://repository.si.edu/handle/10088/1734
Olson, S. L., and D. B. Wingate. 2006. A new species of night-heron (Ardeidae: Nyctanassa) from Quaternary deposits on Bermuda. Proceedings of the Biological Society of Washington 119(2):326-337. https://doi.org/10.2988/0006-324X(2006)119[326:ANSONA]2.0.CO;2
Olson, S. L., and D. B. Wingate. 2012. A new species of towhee (Aves: Emberizidae: Pipilo) from Quaternary deposits on Bermuda. Proceedings of the Biological Society of Washington 125(1):85-96. https://doi.org/10.2988/11-21.1
Peake, D. R., and G. Ritchison. 1998. Nest site selection and nesting success in White-eyed Vireos. Kentucky Warbler 74:64-67.
Pitelka, F. A., and E. J. Koestner. 1942. Breeding behavior of Bell’s Vireo in Illinois. Wilson Bulletin 54(2):97-106. https://www.jstor.org/stable/4157121
Reid, S. G. 1877. The birds of the Bermudas. Bulletin of the American Museum of Natural History 1:393-424.
Ritchison, G., J. A. Hawkins, and B. C. Ritchison. 2019. Brooding and provisioning of nestlings by male and female White-eyed Vireos (Vireo griseus). Avian Biology Research 12(1):28-32. https://doi.org/10.1177/1758155919832138
Robinson, S. K. 1981. Ecological relations and social interactions of Philadelphia and Red-eyed Vireos. Condor 83(1):16-26. https://doi.org/10.2307/1367593
Rust, H. J. 1920. The home life of the Western Warbling Vireo. Condor 22(3):85-94. https://www.jstor.org/stable/1362756
Savidge, J. A. 1987. Extinction of an island forest avifauna by an introduced snake. Ecology 68(3):660-668. https://doi.org/10.2307/1938471
Siepielski, A. M., A. D. Rodewald, and R. H. Yahner. 2001. Nest site selection and nesting success of the Red-eyed Vireo in central Pennsylvania. Wilson Bulletin 113(3):302-307. https://doi.org/10.1676/0043-5643(2001)113[0302:NSSANS]2.0.CO;2
Sterrer, W., A. Glasspool, H. De Silva, and J. Furbert. 2004. Bermuda: an island biodiversity transported. Pages 118-170 in J. Davenport and J. L. Davenport, editors. The effects of human transport on ecosystems: cars and planes, boats and trains. Royal Irish Academy, Dublin, Ireland.
Tossas, A. G. 2008. Reproductive success of the Puerto Rican Vireo in a montane habitat. Wilson Journal of Ornithology 120(3):460-466. https://doi.org/10.1676/07-083.1
Wiley, J. W. 1985. Shiny Cowbird parasitism in two avian communities in Puerto Rico. Condor 87(2):165-176. https://doi.org/10.2307/1366878
Wiley, J. W., and G. M. Kirwan. 2013. The extinct macaws of the West Indies, with special reference to Cuban Macaw Ara tricolor. Bulletin of the British Ornithologists’ Club 133(2):125-156. http://boc-online.org/bulletins/downloads/BBOC1332-Wiley.pdf
Wingate, D. B. 1990. The restoration of Nonsuch Island as a living museum of Bermuda’s precolonial terrestrial biome. Pages 133-149 in G. M. Woodwell, editor. The Earth in transition: patterns and processes of biotic impoverishment. Cambridge University Press, Cambridge, UK.
Winkler, D. W., S. M. Billerman, and I. J. Lovette. 2020. Vireos, Shrike-Babblers, and Erpornis (Vireonidae), version 1.0. In S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, editors. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.vireon1.01
Wood, J. R., J. A. Alcover, T. M. Blackburn, P. Bover, R. P. Duncan, J. P. Hume, J. Louys, H. J. M. Meijer, J. C. Rando, and J. M. Wilmshurst. 2017. Island extinctions: processes, patterns, and potential for ecosystem restoration. Environmental Conservation 44(4):348-358. https://doi.org/10.1017/S037689291700039X
Woodworth, B. L. 1995. Ecology of the Puerto Rican Vireo and the Shiny Cowbird in Guánica Forest, Puerto Rico. Dissertation. University of Minnesota, St. Paul, Minnesota, USA.
Woodworth, B. L. 1997. Brood parasitism, nest predation, and season-long reproductive success of a tropical island endemic. Condor 99(3):605-621. https://doi.org/10.2307/1370473
Fig. 1
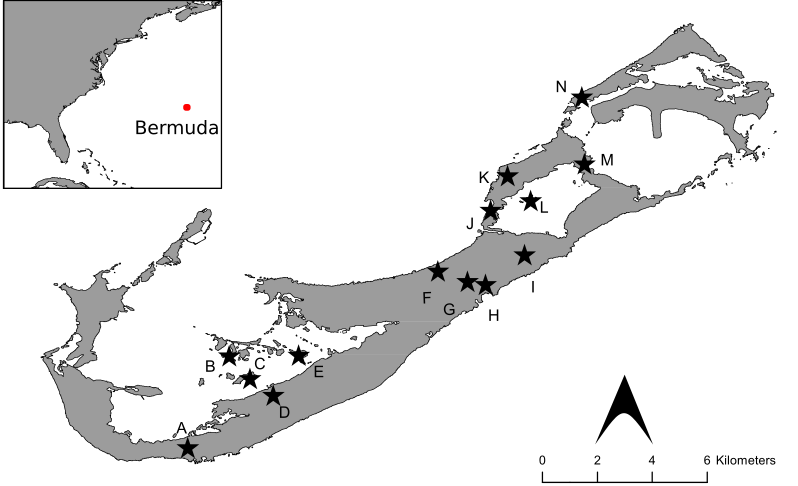
Fig. 1. Map of the Bermuda archipelago depicting all sites (stars) where nesting and breeding behavior of Bermuda Vireos was observed from 2016–2021. A, Seymour’s Pond (1 nest); B, Gamma Island (0 nests); C, Darrel–s Island (2 nests); D, Elm Lodge (1 nest); E, Hinson’s Island (3 nests); F, Oceanview Golf Course (1 nest); G, Brighton Plant Nursery (1 nest); H, Devonshire Bay (1 nest); I, Spittal Pond (29 nests); J, Shelly Bay Railway Trail (1 nest); K, Radnor Road/Shelly Hall Condos (2 nests); L, Trunk Island (1 nest); M, Blue Hole Park/Tom Moore’s Tavern (2 nests); and N, Ferry Reach/Lover’s Lake (38 nests). The inset map depicts the position of Bermuda (red circle) in the western North Atlantic Ocean, approximately 1048 km from the closest point along the east coast of North America (Cape Hatteras).
Fig. 2

Fig. 2. A color-banded male Bermuda White-eyed Vireo (Vireo griseus bermudianus) at Spittal Pond Nature Reserve, Bermuda. Photograph by Miguel Mejías.
Fig. 3
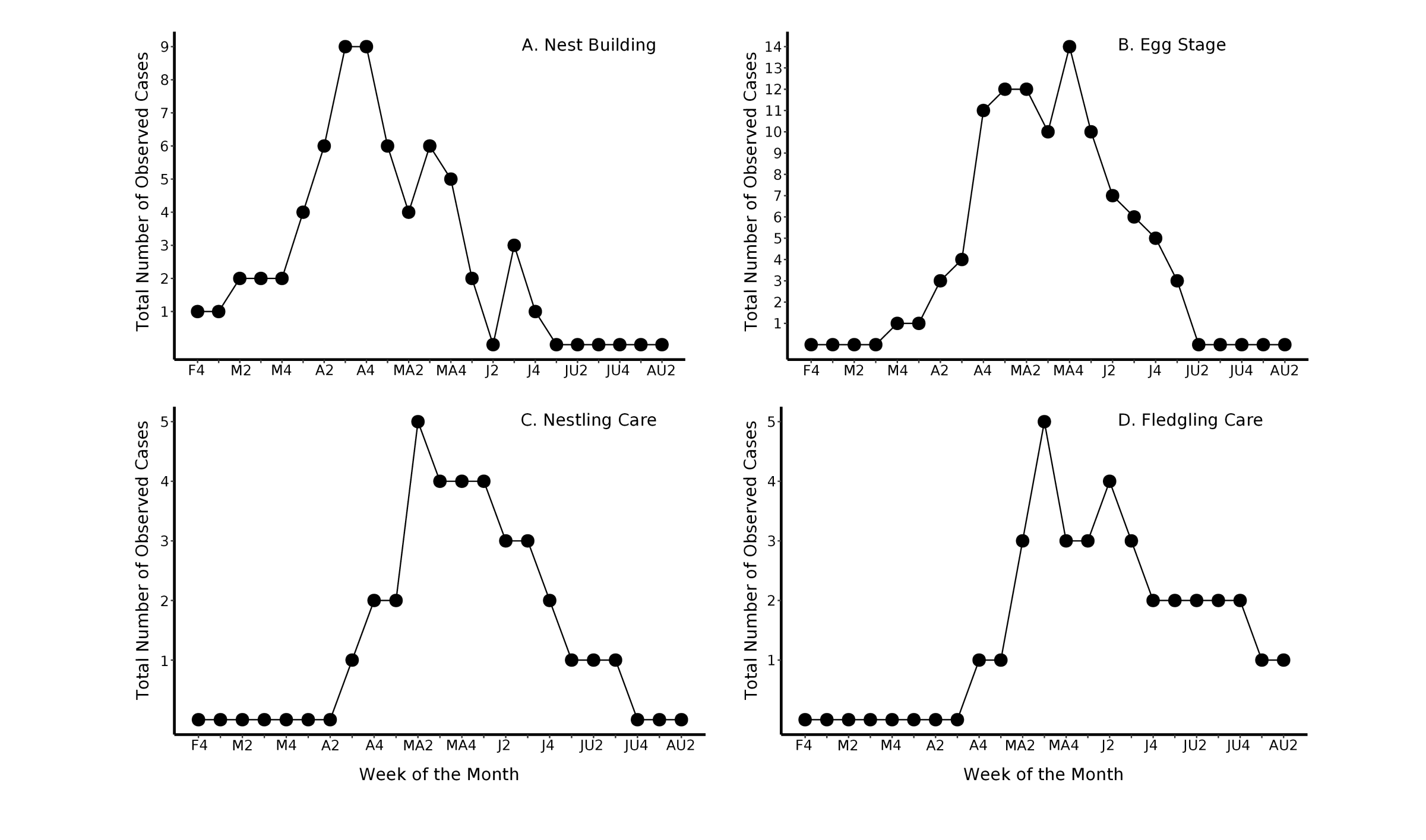
Fig. 3. Breeding phenology of Bermuda Vireos. The number of nests observed in each breeding stage is shown for each week between February and August during the 2016–2021 breeding seasons. Some nests were observed in the same stage over multiple weeks and thus contributed to multiple data points within the breeding stage panel. Because the number of days in each month varied from 28–31, we included any days beyond the 28th day of the month in week 4. Month abbreviations are as follows: F = February, M = March, A = April, MA = May, J = June, JU = July, and AU = August. Note that the final observation of fledgling care in the second week of August corresponds to a single pair whose fledgling care period ended that week.
Fig. 4

Fig. 4. From left to right, photographs show the progression of nest building in allspice (Pimenta dioica) for a breeding pair of Bermuda White-eyed Vireos. Photos were taken on days 0, 3, and 10 of nest construction between 21 and 31 March 2017. Photographs by Miguel Mejías.
Fig. 5
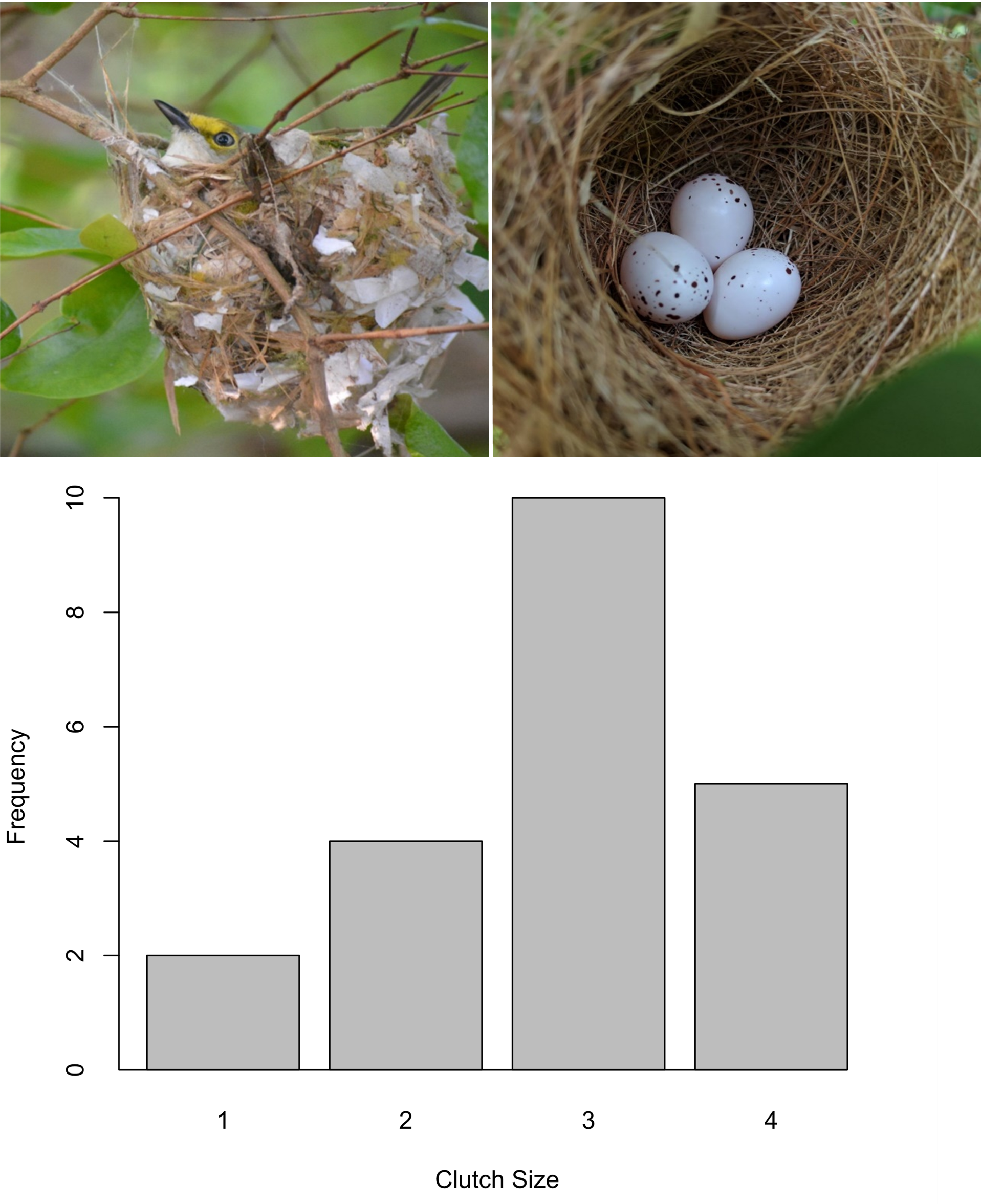
Fig. 5. A Bermuda White-eyed Vireo (left) and a 3-egg clutch inside a nest cup (right); clutches of up to 4 eggs occur in this subspecies. Both the pensile nest hanging from a forked branch and the small, white eggs with brown speckling and blotching are typical of vireonids. The histogram shows the frequency distribution of Bermuda White-eyed Vireo clutch sizes. Photographs by Jorge Sanchez (left) and Miguel Mejías (right).
Fig. 6

Fig. 6. Photographs showing the developmental stages, from top left to bottom right, of the same Bermuda White-eyed Vireo nestlings. Note the gradual change from naked and blind hatchings to feathered nestlings with open eyes. Photographs were taken on days 0 (hatch day), 3, 6, and 9 (fledging day), between 21 and 30 April 2017. Photographs by Miguel Mejías.
Table 1
Table 1. Estimates of pair bond duration of 18 nesting pairs of color-banded Bermuda Vireos from 2016–2021. Color bands (B = blue, G = green, LB = light blue, O = orange, P = pink, PU = purple, R = red, W = white, Y = yellow) on the left and right leg are separated by a slash. A status of “unknown” indicates a pair whose duration together was not followed closely. In all but one pair (Male O/BG and Female O/LBY), the end of a pair bond involved males remaining in their original territory and the females deserting, which often involved pairing with other males in neighboring territories; note female RWG/ was paired with four different males throughout the 2017 breeding season. Once separated, previously paired individuals never re-paired. Dates and pairing durations are conservative because it is possible that birds were paired before the “First seen paired” date and that they remained paired after the “Last seen paired” date.
| Pair (male; female) | First seen paired | Last seen paired (days paired) | Known number of nest attempts | ||||||
| BO/; B/ | 11 October 2016 | 19 July 2017 (282) | 2 | ||||||
| YG/; OPB/ | 29 January 2017 | 18 May 2017 (110) | 1 | ||||||
| R/; OY/ | 20 February 2017 | 26 May 2018 (461) | 1 | ||||||
| BBP/; RWG/ | 20 February 2017 | 20 February 2017 (1) | 0 | ||||||
| BBP/; BWG/ | 24 May 2017 | 11 February 2021 (1360) | 5 | ||||||
| BBP/; OY/GW | 26 May 2021 | 17 November 2021 (176) | 2 | ||||||
| O/; BWO/ | 14 April 2017 | 19 May 2017 (36) | 1 | ||||||
| O/; PUBW/ | 26 May 2018 | 1 August 2019 (433) | Unknown | ||||||
| O/P; O/BO | 3 May 2019 | 14 June 2019 (43) | 2 | ||||||
| O/YB; BWPB/ | 25 April 2019 | 26 December 2019 (246) | 4 | ||||||
| OPU/; RWG/ | 1 March 2017 | 17 March 2017 (17) | Unknown | ||||||
| G/LB; R/LB | 4 August 2017 | 7 August 2018 (369) | 1 | ||||||
| GY/; RWG/ | 14 April 2017 | 20 April 2017 (7) | 1 | ||||||
| BWB/; RWG/ | 28 April 2017 | 21 May 2017 (24) | 1 | ||||||
| G/Y; Y/Y | 13 July 2017 | 28 July 2018 (381) | Unknown | ||||||
| O/BG; O/LBY | 20 December 2018 | 16 August 2019 (240) | 4 | ||||||
| /RP; O/BY | 26 April 2019 | 4 May 2019 (9) | 1 | ||||||
| O/OP; O/LB | 19 December 2018 | 7 June 2019 (171) | 2 | ||||||
Table 2
Table 2. Fate of Bermuda White-eyed Vireo nests. Number of nests observed from 2016–2021 is shown for each nest status or event. For one nest that was depredated in 2019, we were unsure whether it contained eggs or nestlings at the time it was depredated.
| Nest status or event | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | Total | ||
| Nests initiated | 2 | 30 | 9 | 36 | 2 | 5 | 84 | ||
| Nests completed | 2 | 27 | 9 | 34 | 1 | 3 | 76 | ||
| Total nests abandoned | 0 | 5 | 0 | 12 | 0 | 2 | 19 | ||
| Total nests fell apart | 0 | 11 | 0 | 10 | 0 | 0 | 21 | ||
| Nest fell with eggs or nestlings | 0 | 5 | 0 | 2 | 0 | 0 | 7 | ||
| Nest fate unknown | 2 | 6 | 7 | 5 | 1 | 1 | 22 | ||
| Nests with eggs | Unknown | 17 | 3 | 18 | 1 | 3 | 42 | ||
| Known clutch size | Unknown | 12 | 1 | 7 | 0 | 1 | 21 | ||
| Nests with nestlings | Unknown | 8 | 3 | 10 | 1 | 3 | 25 | ||
| Clutch abandoned | Unknown | 1 | 0 | 2 | 0 | 0 | 3 | ||
| Clutch predation | Unknown | 2 | 0 | 2 | 0 | 0 | 4 | ||
| Nestling predation | Unknown | 3 | 0 | 7 | 0 | 1 | 11 | ||
| Nestling fell out of nest | 0 | 1 | 0 | 0 | 0 | 0 | 1 | ||
| Nests produced fledglings | Unknown | 3 | 2 | 3 | 1 | 1 | 10 | ||