







The following is the established format for referencing this article:
Khan, Z. Z., H. S. Sushma, P. Antony B., K. M. Koli, A. Neema, M. Meera, M. Arasumani, V. Robin, R. Jayapal, and S. Mukherjee. 2023. Habitat determinants of species occupancy and niche partitioning among sympatric owlets: the paradoxical role of agricultural lands for the endangered Forest Owlet, Athene blewitti. Journal of Field Ornithology 94(2):1.ABSTRACT
Species with highly restricted distributions are intriguing especially when they co-occur with similar and related species that are more widely distributed. They are also of conservation concern because of their restricted distributions and low population size. The Forest Owlet (Athene blewitti) is an endangered species endemic to the central Indian landscape that occurs in apparently unconnected pockets. Two other species of owlets, with wide distributions in India, Jungle Owlet (Glaucidium radiatum) and Spotted Owlet (Athene brama) co-occur with Forest Owlet. Understanding how species use the habitat matrix is central to prioritizing conservation action. We studied the three species across an extensive part of the predicted range of Forest Owlets through 350 surveys. Our results indicate that Jungle Owlet occupancy (ψ = 0.5896, SE = 0.08) was highest, followed by Forest Owlet (ψ = 0.2517, SE = 0.10), and Spotted Owlet (ψ = 0.1781, SE = 0.07). The Forest Owlet showed a preference for stands of forests with large trees but not necessarily with closed canopies, as well as open habitats (agricultural land cover). The Spotted Owlet showed a preference for agricultural land cover with lower litter and bare ground cover. In contrast, the Jungle Owlet showed a preference for dense forest and canopy cover. The preference for agricultural cover by the Forest Owlet provides important insights into our understanding of the ecology of this owlet. Most significantly, it brings up several conservation challenges for managing this endangered species across a human-use landscape.
RESUMEN
Las especies con una distribución altamente restringida son intrigantes especialmente cuando co-occurren con especies cercanamente emparentadas y similares que están más ampliamente distribuidas. Estas especies son también de interés para la conservación por su distribución restringida y bajo tamaño poblacional. Athene blewitti es una especie amenazada y endémica al paisaje del centro de India donde se encuentra en bolsillos de hábitat aparentemente desconectados. Otras dos especies de búhos, con una amplia distribución en India, Glaucidium radiatum y Athene brama co-occurren con Athene blewitti. Comprender como la especie utiliza la matriz del hábitat es central para priorizar acciones de conservación. Estudiamos las tres especies a lo largo de una porción extensiva del rango predicho de Athene blewitti usando 350 muestreos. Nuestros resultados indican que la ocupación de Glaucidium radiatum (ψ = 0.5896, SE = 0.08) fue la más alta, seguida por Athene blewitti (ψ = 0.2517, SE = 0.10) y Athene brama (ψ = 0.1781, SE = 0.07). Athene blewitti mostro una preferencia por fragmentos de bosque con árboles grandes, pero no necesariamente con dosel cerrado, así como hábitats abiertos (cobertura de agricultura). En contraste, Athene brama mostró una preferencia por bosque denso y con alta cobertura de dosel. La preferencia por coberturas de agricultura por parte de Athene blewitti provee información importante para nuestro entendimiento de la ecología de este búho. Mas significativamente, revela muchos retos en conservación para el manejo de esta especie amenazada a lo largo de un paisaje utilizado por humanos.
INTRODUCTION
Knowledge about and factors determining species occurrences are fundamental for effective conservation planning. Factors influencing species distributions vary at different spatial scales. Often information on species distributions is available at a larger spatial scale but because management decisions are focused on smaller geographical units, a finer resolution of species occurrences and factors influencing them are crucial, especially for endangered species. Human-dominated landscapes in tropical, developing countries continue to increase, creating a matrix of natural and human landscapes. Species may respond to elements of such a matrix depending on their specialization or habitat choice, some are restricted to interior forests, and some may occupy edges and human-use landscapes (Marzluff et al. 2004, Watson et al. 2004). In addition, interspecific interactions such as competition among ecologically similar species could also influence habitat choice and resource use (O’Reilly et al. 2022).
Many studies on owls have reported such interspecific interactions (Lanen et al. 2011, Wiens et al. 2014, Eriksen et al. 2018, Mangan et al. 2019), however, similar studies are inadequate in South Asian countries. In central India, the endangered and endemic Forest Owlet (Athene blewitti) co-occurs with two widely distributed owlets: the Spotted Owlet (Athene brama) and the Jungle Owlet (Glaucidium radiatum). However, the global distribution ranges of the three species are dissimilar (del Hoyo et al. 1999), suggesting varied factors that may limit their occurrences. The Forest Owlet is extremely restricted, with a small and patchy distribution across India’s narrow latitudinal range (19o-22° N; BirdLife International 2018). On the other hand, the closely related Spotted Owlet has a much wider distribution across Asia, occurring throughout India, and is categorized as least concern (BirdLife International 2018). Jungle Owlet is endemic to South Asia (India, Nepal, Bhutan, Bangladesh, and Sri Lanka) but is widespread within this range and listed as least concern (BirdLife International 2018). The Forest Owlet was rediscovered after 113 years (King and Rasmussen 1998) in the same region of Maharashtra. Subsequently, the species has been reported in new locations (Laad and Dagale 2014, Patel et al. 2017, Raha et al. 2017) and has been the focus of several distribution surveys and ecological studies because of its endangered and endemic status (Jathar and Rahmani 2004, Ishtiaq and Rahmani 2005, Mehta et al. 2007, 2017, Patel et al. 2015, 2017). These studies provide a broad understanding of the habitat requirements of the species. However, finer scale, quantitative measures through systematic and robust methods are lacking. Further, the species co-occurs with other similar-sized owlets, therefore it would be pertinent to study them together because there is a gap in our knowledge about what factors influence the distribution of these owlets at the local scale where they co-occur.
In a previous study, species distribution models (SDM) were constructed for the Forest Owlet using “presence-only data” and various combinations of environmental and climatic variables (Mukherjee et al. 2016, Koparde 2019). Although such presence/absence surveys can be implemented at the landscape level, these surveys fail to discriminate between the non-detection and true absence of a species. To understand how factors influence associations between species occurrence and their habitat, it is important to account for imperfect detection and sampling biases (Gu and Swihart 2004). Occupancy models estimate the probability of species presence while accounting for imperfect detection (MacKenzie et al. 2017). They also provide robust estimates for species monitoring to detect changes in site occupancy and thereby help to evaluate management interventions. They are suitable for monitoring especially when density or abundance estimation of rare and cryptic species will likely incur costs in terms of time and resources. Occupancy models have also been used to address ecological questions such as interspecific interactions (Mangan et al. 2019).
We aimed to evaluate the underlying factors determining the occurrence and detectability of the Forest Owlet, Jungle Owlet, and Spotted Owlet, focusing on Forest Owlet in an area where all three are known to co-occur. We used occupancy modeling to examine the role of (1) vegetation structure, (2) land cover variables such as forest and agriculture, and (3) topographic variables on the occurrence of the three owlets. Based on the literature available regarding the species, we expected the forest-associated species, i.e., Forest Owlet and Jungle Owlet, to be positively related to closed vegetation and forest structure, and the Spotted Owlet to show an opposite trend of negative relationships with closed habitats. Further, we explored interspecific associations between the three owlets based on their occupancy probability at each site.
METHODS
Study area
The current known distribution of the Forest Owlet falls within geographic coordinates, 19° N to 22° N and 72° E to 78° E, covering the states of Maharashtra, Gujarat, and Madhya Pradesh, and the Union Territory of Dadra and Nagar Haveli, and Daman and Diu. Recent species distribution models projected the distribution range over a large landscape (16,500 km²) within these states (Mukherjee et al. 2016, Koparde 2019). The landscape falls within the north-western Ghats and the Satpura and Nimar hill range with elevations ranging from 50 to 1350 m. Much of this landscape is human dominated. The forests within these regions are listed under the Champion and Seth (1968) categorization of Tropical Moist and Dry Deciduous Teak forests of the types 3B/C1b in Gujarat and Maharashtra and 5A/C1, C1a and C1b in Madhya Pradesh.
Study design
We created polygons around the niche clusters in the intersection region of the two SDMs (Mukherjee et al. 2016, Koparde 2019) to identify areas for sampling. Niche clusters included both high probability areas and a few low probability areas adjoining the high probability area predicted by the two SDMs. Polygons were drawn over three focused regions namely: Dangs, Tansa, and Toranmal in Maharashtra and Gujarat from where the owlets are known to occur (Appendix 1, Fig. A1.4). We then overlaid 1 x 1 km grids over these polygons, using ArcGIS v. 10.5 (Fig. 1). Because the home ranges of Forest Owlet and its sympatric owlets found in India are unknown, we used the maximum known home range (95.0 ha) of the Little Owl, Athene noctua, (Grzywaczewski 2009), a phylogenetically close relative of the Forest Owlet (Koparde et al. 2018), to determine the grid size.
From the grids generated, we removed grids that had ≥ 50% of the area covered with non-forest habitats such as large agricultural fields, urban settlements, wastelands, large water bodies, etc. because these are not considered to be suitable habitats. However, we included grids that had forest edges with agriculture and grasslands because these areas are known to be used by multiple species including the specialist Forest Owlet (Jathar and Rahmani 2004, Mehta et al. 2015, Kulkarni and Mehta 2020). We included “grasslands” as a class because the available land cover data from central India (NRSC LULC classification) includes deciduous forests with less than 50% canopy cover classified as grasslands (NRSC 2014). We then randomly selected 2.5% of the grids from the niche polygons from an area of approximately 3500 km² for the occupancy surveys. Within these grids, we assessed the habitat occupancy of the three owlets using temporally replicated surveys.
Survey methods
We conducted the surveys between November 2018 and May 2019, which coincided with the breeding season of the three owlets. The Forest Owlet is diurnal and crepuscular (Jathar and Rahmani 2004, Ishtiaq and Rahmani 2005), while the Jungle Owlet is crepuscular, and the Spotted Owlet is nocturnal (Ali 1996). Therefore, we conducted the surveys during the morning (0600 to 1000 h) and evening (1530 to 1900 h) hours. In each grid, we located its centroid and scanned the surroundings for direct sightings of the three owlets. In addition, we also played back calls of the three owlets because this method has been successfully used to detect Forest Owlets in central India (Ishtiaq and Rahmani 2000, Jathar and Rahmani 2004). If the centroid location was inaccessible, an alternate location within 200 m of the original point was selected. A hand-held Zoom H1 handy portable digital recorder (volume stop 100) was used to play the calls. The playback volume was standardized to amplitudes similar to natural calls following the protocols of Darras et al. (2018). The recorder was connected by an aux cable to a Sony SRS-XB10 portable Bluetooth speaker, which was usually placed on a tree. We first played the Forest Owlet calls. We played the contact “Kuhu” call once and the territorial “Kwak” call thrice and then the contact “Kuhu” call once again (call descriptions follow Ishtiaq and Rahmani 2005). This was followed by five minutes of silent listening. We repeated this twice if there was no response to the broadcasted calls. Next, we played the Jungle Owlet calls, followed by five minutes of listening. We followed the same protocol to detect the Spotted Owlet. If any of our target species was detected before or during the call playback of another owlet, its call was not played further. If our target species were detected during the call playback, then we stopped the trials for the species. We carried out these surveys for two consecutive days leading to four temporal replicates of the call playback survey.
Two trained observers surveyed each grid, and the same observers visited the grids for all the replicates, to avoid within-grid observer bias. A total of four observers in teams of two carried out surveys simultaneously in different grids. Each type of sign (sighting or call) was recorded as detection (1), while the absence of these signs was recorded as non-detection (0). If a response call was recorded, it was followed by a visual confirmation of the species. If we could not conduct a survey due to inclement weather or logistic constraints, we recorded it as a missing observation (“-”) while building detection histories for the analyses. We surveyed 88 grids, i.e., Dangs (58), Tansa (18), and Toranmal (11), putting a total field effort of 1703 person-days of surveys.
Site-specific covariates
We laid four circular plots of 10 m radius at 200 m intervals within each grid. We selected a set of variables to measure based on our a priori hypotheses on the occurrence of the species (Table 1). We measured habitat variables including tree height (m), girth at breast height (GBH) of trees (cm), tree species richness and abundance, and canopy cover (%) using a densiometer (Strickler 1959) at the 4 corners of an imaginary square within each 10-m radius plot. We also assessed ground cover (shrubs/herbs/grasses, bare ground, litter, and rock cover) in the 1-m² quadrat laid within the primary 10-m plots. In addition to the ground-based surveys, we collected remotely sensed covariates to examine the influence of habitat variables on species occupancy.
Land cover and topographic variables
We used the ASTER global digital elevation model (GDEM version 3) 30-m resolution (NASA/ASTER Team 2019) dataset to compute topographic variables. The dataset included separate DEM tiles, which were mosaicked to obtain mean values for elevation, slope, and aspect using the zonal statistics plugin in QGIS v. 3.10 (QGIS Development team 2019). We also derived the topographic ruggedness index (TRI) for each sampled grid. Land cover covariates such as forest cover (FC), grassland (GC), human settlements (HS), agriculture (AC), and waterbody were derived by supervised classification of Sentinel-2 imagery in Google Earth Engine (Gorelick et al. 2017). The classified land use and land cover data were used to calculate the edge density (m/ha) of the five classes in each survey grid in Fragstats 4.2 (McGarigal et al. 2012).
Previous studies reported that the Forest Owlet was also found in edge habitats near agriculture (Kulkarni and Mehta 2020), thus we calculated the edge density, which is expected to have a positive influence on the occupancy of the species. Shuttle radar topography mission (SRTM version 3.0), global (90-m resolution) data (NASA JPL 2013) was used to generate drainage lines and to compute drainage density (km-¹) using basin area. This was calculated using the formula drainage density = stream length/basin area. Because historical records report Forest Owlet occurrence in riverine forests (Ball 1877), we used enhanced vegetation index (EVI) as a surrogate for riverine areas within the grids because these patches could not be classified from Sentinel imagery due to the limitations of spatial and spectral resolution. Enhanced vegetation index is a measure of “greenness” or “green biomass” in a cell and is generally considered an alternative for forest types. Values of the EVI range from -1 to 1 in which lush and evergreen vegetation generally falls between values of 0.20 to 0.80. We used mean EVI for the analyses. We reasoned that the grids having riverine patches would remain “evergreen” throughout the year and therefore have higher mean EVI values than grids having dry forest types and barren or fallow lands. We processed and analyzed all the layers in QGIS 3.10 and ArcGIS 10.5.
Survey specific covariates
Detection of a species may be influenced by variables specific to each survey and therefore need to be modeled. We hypothesized that the detection of our study species would be influenced by factors such as ambient light (lx) and ambient noise (db). We measured these factors during each survey to determine their influence on the species’ detection probability. We measured ambient light (lx) through Lux Meter (Light Meter application v. 1.5; My Mobile Tools Dev 2018) and ambient noise (db) through the Sound Meter application v. 3.6 (Abc Apps 2018), using the same model of android smartphones. The readings were first calibrated on all the devices, and the average reading for 30 seconds was taken for both ambient light and noise before initiating call playback.
Data analysis
Habitat occupancy
We used occupancy modeling to determine factors influencing the distribution of the three owlets in the landscape. Occupancy models measure two key parameters: ψ, the probability that a site is occupied by the species, and p, the probability of detecting the species presence in a replicate, given that the grid is occupied by the species (MacKenzie et al. 2017). Because some of the variables may be correlated and collinear, we first performed Pearson’s correlation coefficient analysis on the entire set of covariates. Covariates that had r < 0.6 were not included in the same model. We also used the variance inflation factor (VIF) to assess collinearity between pairs of covariates. The final set of covariates used in the modeling and their predicted influence on the occupancy and detection probability of the three species is given in Table 1. We standardized all covariates (z-scores) before performing the occupancy modeling in the program PRESENCE v. 13.5 (Hines 2006). We constructed a global model containing all potential variables influencing the two model parameters (Burnham and Anderson 2002, MacKenzie et al. 2017). Subsequently, we constructed alternative plausible models to identify covariates influencing the model parameters. We performed a goodness of fit (GOF) test (MacKenzie and Bailey 2004) on the global model to assess model fit (10,000 bootstraps). The models were ranked based on Akaike information criterion (AICc) values and corrected for small sample size (Burnham and Anderson 2002). If the GOF test revealed overdispersion (ĉ > 1), then the models were ranked based on quasi Akaike information criterion (QAICc) values corrected for small sample size (Anderson et al. 1994). We performed model averaging if a single top model did not emerge and considered models with ΔAICc < 2 (Burnham and Anderson 2002). We used single-species, single-season occupancy models (MacKenzie et al. 2002) in PRESENCE v. 13.5 (Hines 2006). Further, model-averaged predictions were used to produce the response curves for each covariate (Burnham and Anderson 2002) from the top-ranking models using the package “unmarked” (Fiske and Chandler 2011) in R (R Core Team 2020). We overlaid the three owlets' cell level occupancy probabilities and mapped the occupancy estimates range across all survey grids (Fig. 2) using QGIS.
Interspecific association amongst the owlets
We estimated the correlation between occupancy estimates for the owlets using Spearman’s rank correlation test and obtained p values using the “rcorr” function in the package “Hmisc” (Harrell and Dupont 2020). We created scatter plots of the correlation matrix (Fig. 3) using the function “ggscatter” in the package “ggpubr” (Kassambara 2020). Both analyses were performed in R (R Core Team 2020).
RESULTS
Out of 350 surveys (88 grids visited ~ 4 times), we detected Forest Owlets in 22 grids (positive detections = 71), Jungle Owlets in 51 grids (positive detections = 140), and Spotted Owlets in 15 grids (positive detections = 34). The GOF test (with 10,000 bootstraps) revealed overdispersion in the case of Forest Owlet and Jungle Owlet (ĉ > 1), thus the value of ĉ was used to calculate QAICc for both species. We did not find any overdispersion in the case of the Spotted Owlet (ĉ < 1) and used AICc as the ranking criterion for this species. The QAICc and AICc values were calculated using the number of positive detections (71, 51, and 34 for Forest Owlet, Jungle Owlet, and Spotted Owlet, respectively) as the effective sample size (Burnham and Anderson 2002). See Table 2.
Forest Owlet
No single top-ranked model emerged. From a candidate set of 22 models, 6 models seemed to fit the data well, ΔQAICc < 2 (Appendix 1, Table A1.1). We, therefore, performed model averaging to derive final estimates of the model parameters (Burnham and Anderson 2002). Comparisons between all the models are reported in Table 2. Our analysis indicated GBH, agricultural land cover (%), and elevation had a positive influence while bare ground cover (%) and EVI had a negative influence on the site occupancy (ψ). Ambient noise had a negative influence on the detection probability (p) of the Forest Owlet (Table 3; Fig. 4). The model-averaged estimate of detection probability (p) of the Forest Owlet was 0.7871 (SE = 0.06), while the probability of occupancy (ψ) was 0.2517 (SE = 0.10), which was close to the naive occupancy estimate of 0.2500.
Jungle Owlet
As in the case of the Forest Owlet, no single best model emerged. From a candidate set of 8 models, 2 models fitted the data well, ΔQAICc < 2 (Appendix 1, Table A1.2). The analysis indicated canopy cover (%), forest cover (%) had a positive influence and bare ground cover (%) had a negative influence on the site occupancy (ψ) of the Jungle Owlet (Fig. 4). Ambient light had a positive influence on its detection probability (p). The model-averaged estimate of detection probability (p) for the Jungle Owlet was 0.6843 (SE = 0.04), while the probability of occupancy (ψ) was 0.5896 (SE = 0.08), which was close to the naive occupancy estimate of 0.5795.
Spotted Owlet
Only one top model fitted the data well (ΔAICc < 2) from a set of 58 models (Appendix 1, Table A1.3). Agricultural land cover (%) showed a positive influence, while bare ground cover (%) and litter ground cover (%) showed a negative influence on the site occupancy (ψ) of the Spotted Owlet (Fig. 4). Ambient light had a negative influence on its detection probability (p). The detection probability (p) for the Spotted Owlet was 0.5546 (SE = 0.08), while the probability of occupancy (ψ) was 0.1781 (SE = 0.07), which is close to the naive occupancy estimate of 0.1705.
Habitat partitioning among the three owlets
We found no significant correlation between the occupancy of Forest Owlet and Jungle Owlet (Rs = -0.18606, N = 86, p > 0.05). See Fig. 3. There was a weak negative correlation between the occupancy of Jungle Owlet and Spotted Owlet (Rs = -0.2435, N = 86, p < 0.05), and a significant positive correlation between the occupancy of Forest Owlet and Spotted Owlet (Rs = 0.6719, N = 86, p < 0.05).
DISCUSSION
Our study provides insights into the landscape and environmental attributes associated with the occurrences of the endangered Forest Owlet and its sympatric Jungle and Spotted owlets in central India. We found that the probability of occupancy was highest for Jungle, followed by Forest, and Spotted owlets.
Role of large trees and open cultivated land for Forest Owlet
Our results indicated that both forests with large trees (as indicated by high GBH values) and open cultivated lands (as indicated by high AC) positively influence the occupancy of the Forest Owlet (Fig. 4). Our results concur with previous studies that have shown that the species prefers sites that have a mix of these habitats. Although this may seem contradictory, specific ecological reasons may underlie the observed habitat choice as shown by previous studies. These studies reported that the species preferred large trees for nesting (Jathar and Rahmani 2004, Ishtiaq and Rahmani 2005, Mehta et al. 2007, 2017, Kulkarni and Mehta 2020). Open habitats are important for the species because they perhaps facilitate foraging efficiency (Jathar and Rahmani 2004, Mehta et al. 2018). Because rodents comprise a major proportion of its diet (Mehta et al. 2018), open cultivated lands, which possibly attract rodents, seem to influence the occurrence of Forest Owlet.
Other studies have also emphasized the importance of small agricultural holdings interspersed with forests for the species (Patel et al. 2015, 2017, Patel 2018). However, it is important to note that the current habitat, a mosaic of forested land and agriculture, is likely a remnant of its original habitat, as reported in historical records (BirdLife International 2018).
Role of closed forest and elevation for Forest Owlet
Although early literature suggested a preference for riverine and dense forests (Ball 1877, Ali and Ripley 1969), our results across a large landscape do not support this idea (negative influence of EVI on occupancy). This pattern is consistent with other studies stating that the species occurs in areas with lower canopy cover and tree density (Jathar and Rahmani 2004) and moderately dense forests (Mehta et al. 2008, 2015). This could also be because the original habitats with dense forests are no longer available or are confined to very few and small pockets due to deforestation and land use change (Kaul et al. 2009, Reddy et al. 2016), which our random sampling design does not represent adequately.
Elevation had a weak positive influence on the occurrence of the Forest Owlet, even though it was a covariate in one of the top-ranking models. Although the species has been recorded from 50-920 m, the higher elevation records are limited (Ishtiaq and Rahmani 2000, Jathar and Rahmani 2004, Patel et al. 2015, Kulkarni and Mehta 2020). Our random sampling design with relatively fewer spatial replicates may have also limited our elevational sampling, and it does not permit us to specifically test the elevation hypothesis for the Forest Owlet.
Fine scale habitat partitioning between the species
Forest Owlet and Spotted Owlet occurrence were positively associated with the presence of agriculture and negatively with bare ground cover (Fig. 4), reflecting patterns documented by others (Pande et al. 2006, 2007, Mahmood-ul-Hassan et al. 2007, Vanitha et al. 2018). Small mammals and invertebrates, which prefer habitats with more shrub, herb, and grass cover, are a major part of the owlet diet (Mehta et al. 2018), which explains their negative association with bare ground cover. This is reflected in the higher spatial overlap between the two species (Fig. 3). Both species appear to use similar foraging sites although avoiding competition through temporal separation in their activity. On the contrary, there may be habitat partitioning of the Forest Owlet with the Jungle Owlet. The latter occurs in areas associated with higher forest and canopy cover (Fig. 4); a pattern supported by studies on Jungle Owlet from other parts of India (Jayson and Sivaram 2009). We note that we are only reporting fine-scale habitat preferences, and any inference on their niche and competition must consider their diet, which was not examined in this study. The Forest Owlet appears to be using an intermediate habitat (with positive influences of agriculture and forests with trees having a high GBH), which overlaps more with the Spotted Owlet than the Jungle Owlet. However, for a more detailed study of spatial segregation or overlap between the three species, we suggest a multi-species occupancy modeling approach (Rota et al. 2016).
Landscape/local differences in occupancy
The average occupancy of Forest Owlet was higher around Dangs (0.27) and Tansa (0.24) compared to Toranmal RF (0.14). Forests in Toranmal RF are highly degraded (Jathar and Rahmani 2004, Jathar and Patil 2011) and had the lowest occupancy for all three owlets (Fig. 2). In contrast, the forest in and around Dangs and Tansa WLS still have pockets of teak plantations and disturbed forests. This habitat is also preferred by the Jungle Owlet, which had the highest occupancy overall among the three owlets (Jayson and Sivaram 2009). Because of limitations in sample sizes, we pooled datasets from Tansa WLS and Dangs in the formal analyses for a combined estimate of occupancy.
We note that our results for the Spotted Owlet must be taken with caution because our survey design did not include grids with extensive human settlements or agricultural fields that the species is strongly associated with (Pande et al. 2006, 2007, Mahmood-ul-Hassan et al. 2007, Vanitha et al. 2018).
Factors affecting detection probability and recommendations for future surveys
Detection probability and playback
Despite the relative rarity of the Forest Owlet, its detection probability was fairly high (~78%) and closely followed by the more common Jungle Owlet and the Spotted Owlet. Playback of two different calls (the contact call “Kuhu” and the territorial call “Kwak”), calls described in Ishtiaq and Rahmani (2005), may have increased the detectability of the Forest Owlet, and the timing of the survey during the breeding season when the birds are most vocal (Ishtiaq and Rahmani 2000, Mehta et al. 2015). In fact, playback may have aided the detection of all three sympatric owlets, as reported in other studies on owls in India (Ishtiaq and Rahmani 2000, Mehta et al. 2008, Jayson and Sivaram 2009). The owlets also responded to interspecific calls (Jungle Owlet responding to Forest Owlet and Spotted Owlet calls and vice versa), although these instances were few (n = 17). Jungle Owlet was highly vocal compared to the other two owlets and was often detected calling naturally (n = 25). It also responded to conspecific and interspecific calls. Similar behavior has been reported in a study from southern India, in which the encounter rate for Jungle Owlet was highest among the six other species of owls (Jayson and Sivaram 2009).
The use of calls needs to be tested further to determine its effects on the occupancy and detection of these species. The current study was conducted from November to May, coinciding with the breeding season of the three species. Increased territorial behavior and calling during the breeding season may have facilitated their detection, as suggested by other studies on owls (Jathar and Rahmani 2004, Ishtiaq and Rahmani 2005, Jayson and Sivaram 2009). However, multi-season and multi-year studies are required to support this observation, to determine the extent to which changes in behavior impact their detectability, and to learn which season is best suited for such surveys.
Noise negatively impacted the detection of the Forest Owlet. However, we were unable to assess whether noise affects detectability, or if the birds have a behavioral avoidance to noise. Factors like wind, precipitation, temperature, and time of night (for nocturnal species) may also affect the detection probability of owls, all of which could not be factored into this study because of logistic/resource constraints.
Recommendations
We sampled 2.5% of a relatively large part of the predicted landscape of the Forest Owlet and generated baseline occupancy estimates for the three species of owlets. We also provide some key insights on survey methods that can be used for future monitoring of their distribution range within this region. We note that although the occupancy of the Forest Owlet was relatively low, for a bird that was considered critically endangered until recently, this estimate is still high (~ 25%). Given the high detection probability of Forest Owlet in sharp contrast to its relatively low occupancy, we suggest future surveys be optimized with more spatial coverage and fewer temporal replicates in the central Indian landscape, in line with the recommendations from Mackenzie and Royle (2005). Because of the high detectability with calls, we suggest using passive sound recording using autonomous recording units (ARU) as an additional, non-intrusive method for long-term occupancy surveys of rare and endangered species (Pérez-Granados et al. 2018).
Conservation challenges
The importance of agricultural land for the Spotted Owlet is well established (Mahmood-ul-Hassan et al. 2007, Ali and Santhanakrishnan 2015, Vanitha et al. 2018). However, in the case of the endangered Forest Owlet, its apparent preference for agricultural land may raise major conservation challenges. It requires large trees for nesting and roosting (Jathar and Rahmani 2004, Ishtiaq and Rahmani 2005, Mehta et al. 2007, 2017, Kulkarni and Mehta 2020), while exploiting open areas and agricultural land for foraging (Patel et al. 2015, 2017, Patel 2018, Kulkarni and Mehta 2020). This mosaic of forested and agricultural land is likely a remnant of its original habitat. However, these areas are vulnerable to constant threats such as agricultural expansion, encroachment, habitat degradation, and fragmentation due to anthropogenic activities (Mehta et al. 2008, Jathar and Patil 2011, Patel et al. 2015, Mehta et al. 2017, Kulkarni and Mehta 2020). In Toranmal, threats due to loss of vegetation, illicit wood cutting, and collection of firewood, especially of teak, are well documented (Jathar and Patil 2011). The Forest Owlet occurrence is known to be strongly associated with these teak-dominated forests (Jathar and Rahmani 2004, Ishtiaq and Rahmani 2005, Mehta et al. 2007, 2008, 2017, Kulkarni and Mehta 2020), and it is known to avoid areas with anthropogenic activities such as fuelwood collection and logging (Kulkarni and Mehta 2020). The vegetation cover has shrunk in this region due to the expansion of agriculture (Jathar and Patil 2011, Roy et al. 2015), and this continued expansion will likely exacerbate existing pressure on the remaining forest patches in sustaining this species.
It is important to note that among the three regions sampled in this study, only areas within Purna WLS (in Dangs) and Tansa WLS are under formal protection. However, several records of the species are from outside the protected area (Fig. 2), making monitoring these threats and their regulation a major challenge for their conservation. Community engagement and changing traditional perceptions and superstitions around owlets, especially among landowners bordering these forests, is yet another challenge that needs to be addressed alongside protection for long-term success. With a large dependency on forest produce, which was observed in all the field sites, creating alternate livelihood options while maintaining political will for the same is a conservation hurdle that needs to be overcome.
CONCLUSION
We provide baseline occupancy estimates for the three species of owlets, which can be useful for monitoring and planning future surveys. Our results show that the endangered Forest Owlet occupies a habitat niche that includes forests with large trees and agricultural lands and is more common in some parts of its range than previously thought. Among the other sympatric owlets, Jungle Owlet shows a preference for the dense forest with low bare ground cover, while Spotted Owlet shows a preference for habitats with higher agricultural land cover. The high detection probability (> 50%) of all three species with call playback methods indicates a greater potential for acoustic detections in future studies.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Z. K.: investigation, data curation, formal analysis, validation, visualization, writing (original draft); H. S. S.: conceptualization, methodology, validation, writing (original draft), writing (review and editing), visualization, supervision; P. A. and A. N.: investigation, data curation, formal analysis. K. K.: investigation, data curation; M. M. and M. A.: formal analysis; V. V. R., R. J., and S. M.: conceptualization, methodology, validation, resources, writing (review and editing), visualization, supervision, funding acquisition, project administration.
ACKNOWLEDGMENTS
The study was funded by the Ministry of Environment, Forest and Climate Change, GoI, through a grant to Shomita Mukherjee, Rajah Jayapal, and V. V. Robin (File Number: J.22012/61/2009-CS(W), dated: 29th September 2017). We thank the State Forest Departments of Gujarat (No:WLP/RTC/28/C181-196/2018-19, dated 09/07/2018) and Maharashtra (No. Desk-22(8)/WL/CR-45/16-17/3448/2017-18, dated 06/02/2018) for permits to conduct our study. We used the land use/land cover information in our research work from Natural Resources Census Project of National Remote Sensing Centre (NRSC), ISRO, Hyderabad, India. We thank the Director and staff of the Sàlim Ali Centre for Ornithology and Natural History for their support during the project. We thank the Ecology lab members from IISER Tirupati, and specifically Viral Joshi, Jobin Varughese, and Krishna Prasad for their help with standardizing acoustic protocols and volume settings for playback. We thank our field assistants (Tanmay Gokhale, Ravikiran Kashid, Vaheada Sanah A, Hiren Patel, Viraj Athalye, and Prasad Gouraje) for their help during fieldwork.
DATA AVAILABILITY
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
LITERATURE CITED
Abc Apps Team. 2018. Version 3.6. Cincinnati, Ohio, USA. https://play.google.com/store/apps/details?id=com.gamebasic.decibel&hl=en_IN&gl=US
Ali, S. 1996. The book of Indian birds. 12th edition. Oxford University Press, Oxford, UK.
Ali, S., and S. D. Ripley. 1969. Handbook of the birds of India and Pakistan: together with those of Nepal, Sikkim, Bhutan, and Ceylon, volume 3: stone curlews to owls. First edition. Oxford University Press, Mumbai, Maharashtra, India.
Ali, A. H. M. S., and R. Santhanakrishnan. 2015. Nest trees, habitat and breeding biology of the Spotted Owlet Athene brama brama (Temminck, 1821) in human habitation and agricultural landscape of India. Zoology and Ecology 25(3):211-219. https://doi.org/10.1080/21658005.2015.1061248
Anderson, D. R., K. P. Burnham, and G. C. White. 1994. AIC model selection in overdispersed capture-recapture data. Ecology 75(6):1780-1793. https://doi.org/10.2307/1939637
Ball, V. 1877. Notes on birds observed in the region between the Mahanadi and Godavari rivers. Stray Feathers 5:410-420.
BirdLife International. 2018. Heteroglaux blewitti. The IUCN Red List of threatened species 2018:e.T22689335A132251554. BirdLife International, Cambridge, UK. http://dx.doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22689335A132251554.en
Burnham, K. P., and D. R. Anderson. 2002. Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag, New York, New York, USA.
Champion, H. G., and S. K. Seth. 1968. A revised forest types of India. Manager of Publications, Government of India, Delhi, India.
Darras, K., B. Furnas, I. Fitriawan, Y. Mulyani, and T. Tscharntke. 2018. Estimating bird detection distances in sound recordings for standardizing detection ranges and distance sampling. Methods in Ecology and Evolution 9(9):1928-1938. https://doi.org/10.1111/2041-210X.13031
del Hoyo, J., A. Elliott, and J. Sargatal. 1999. Handbook of the birds of the world. Barn-owls to Hummingbirds. Lynx Edicions, Barcelona, Spain.
Eriksen, A., and P. Wabakken. 2018. Activity patterns at the Arctic Circle: nocturnal Eagle Owls and interspecific interactions during continuous midsummer daylight. Journal of Avian Biology 49(7):e01781. https://doi.org/10.1111/jav.01781
Fiske, I., and R. Chandler. 2011. Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. Journal of Statistical Software 43(10):1-23. https://doi.org/10.18637/jss.v043.i10
Gorelick, N., M. Hancher, M. Dixon, S. Ilyushchenko, D. Thau, and R. Moore. 2017. Google Earth Engine: planetary-scale geospatial analysis for everyone. Remote Sensing of Environment 202:18-27. https://doi.org/10.1016/j.rse.2017.06.031
Grzywaczewski, G. 2009. Home range size and habitat use of the Little Owl Athene noctua in East Poland. Ardea 97(4):541-545. https://doi.org/10.5253/078.097.0420
Gu, W., and R. K. Swihart. 2004. Absent or undetected? Effects of non-detection of species occurrence on wildlife-habitat models. Biological Conservation 116(2):195-203. https://doi.org/10.1016/S0006-3207(03)00190-3
Harrell, F. E. Jr., and C. Dupont. 2020. Hmisc: Harrell miscellaneous. Version 4.4-0. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/Hmisc/index.html
Hines, J. E. 2006. PRESENCE2 - software to estimate patch occupancy and related parameters. Version 2.4. United States Geological Survey-Patuxent Wildlife Research Center (USGS-PWRC), Reston, Virginia, USA. https://doi.org/10.1177/089801019201000403
Ishtiaq, F., and A. R. Rahmani. 2000. Further information on the status and distribution of the Forest Owlet Athene blewitti, India. Forktail 16:225-130. https://biostor.org/reference/250868
Ishtiaq, F., and A. R. Rahmani. 2005. The Forest Owlet Heteroglaux blewitti: vocalization, breeding biology and conservation. Ibis 147(1):197-205. https://doi.org/10.1111/j.1474-919x.2004.00389.x
Jathar, G. A., and D. N. Patil. 2011. Reassessment of the status of Forest Owlet in its known distribution and evaluation of conservation issues. Final report. Watershed Organization Trust, Pune, Maharashtra, India.
Jathar, G. A., and A. R. Rahmani. 2004. Ecological studies of the Forest Spotted Owlet Athene (Heteroglaux) blewitti. Final report. Bombay Natural History Society, Hornbill House, Mumbai, India.
Jayson, E. A., and M. Sivaram. 2009. Ecology and behaviour of Forest Owls in the Western Ghats and developing a habitat model for their conservation. Research Report No.343. Kerala Forest Research Institute, Peechi, Thrissur, India. https://www.academia.edu/31641316/Ecology_and_behaviour_of_forest_owls_in_the_Western_Ghats_and_developing_a_habitat_model_for_their_conservation
Kassambara, A. 2020. ggpubr: 'ggplot2' Based Publication Ready Plots. Version 0.4.0. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/ggpubr/ggpubr.pdf"
Kaul, M., V. K. Dadhwal, and G. M. J. Mohren. 2009. Land use change and net C flux in Indian forests. Forest Ecology and Management 258(2):100-108. https://doi.org/10.1016/j.foreco.2009.03.049
King, B. F., and P. C. Rasmussen. 1998. The rediscovery of the Forest Owlet Athene (Heteroglaux) blewitti. Forktail 14:53-55. https://static1.squarespace.com/static/5c1a9e03f407b482a158da87/t/5c1f58468a922d911fea34ec/1545558086638/King-Owlet.pdf
Koparde, P. 2019. Molecular phylogeny and comparative biogeography of owlets in India. Thesis. Manipal Academy of Higher Education, Manipal, Karnataka, India.
Koparde, P., P. Mehta, S. Reddy, U. Ramakrishnan, S. Mukherjee, and V. V. Robin. 2018. The critically endangered Forest Owlet Heteroglaux blewitti is nested within the currently recognized Athene clade: a century-old debate addressed. PLoS ONE 13(2):e0192359. https://doi.org/10.1371/journal.pone.0192359
Kulkarni, J., and P. Mehta. 2020. Habitat selectivity by the Forest Owlet Athene blewitti in Nandurbar District, Maharashtra, India. Indian BIRDS 16(2):33-39. https://indianbirds.in/pdfs/IB_16_2_Kulkarni_Mehta_ForestOwletHabitat.pdf
Laad, S., and R. Dagale. 2014. First report of Forest Owlet Heteroglaux blewitti from Tansa Wildlife Sanctuary (Western Ghats), Maharashtra, India. Journal of Bombay Natural History Society 111(2):134. https://doi.org/10.17087/jbnhs/2014/v111i2/72238
Lanen, N. J. V., A. B. Franklin, K. P. Huyvaert, R. F. Reiser, and P. C. Carlson. 2011. Who hits and hoots at whom? Potential for interference competition between Barred and Northern Spotted owls. Biological Conservation 144(9):2194-2201. https://doi.org/10.1016/j.biocon.2011.05.011
MacKenzie, D. I., and L. L. Bailey. 2004. Assessing the fit of occupancy of site-occupancy models. Journal of Agricultural Biology and Environmental Statistics 9(3):300-318. https://doi.org/10.1198/108571104X3361
MacKenzie, D. I., J. D. Nichols, G. B. Lachman, S. Droege, J. A. Royle, and C. A. Langtimm. 2002. Estimating site occupancy rates when detection probabilities are less than one. Ecology 83(8):2248-2255. https://doi.org/10.1890/0012-9658(2002)083[2248:ESORWD]2.0.CO;2
MacKenzie, D. I., J. D. Nichols, J. A. Royle, K. H. Pollock, J. E. Hines, and L. L. Bailey. 2017. Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier, San Diego, California, USA.
MacKenzie, D. I., and J. A. Royle. 2005. Designing occupancy studies: general advice and allocating survey effort. Journal of Applied Ecology 42(6):1105-1114. https://doi.org/10.1111/j.1365-2664.2005.01098.x
Mahmood-ul-Hassan, M., M. A. Beg, M. Mushtaq-ul-Hassan, and S. A. Rana. 2007. Nesting and breeding habits of the Spotted Owlet (Athene brama) in Punjab, Pakistan. Journal of Raptor Research 41(1):50-52. https://doi.org/10.3356/0892-1016(2007)41[50:NABHOT]2.0.CO;2
Mangan, A. O., T. Chestnut, J. C. Vogeler, I. K. Breckheimer, W. M. King, K. E. Bagnall, and K. M. Dugger. 2019. Barred Owls reduce occupancy and breeding propensity of Northern Spotted Owl in a Washington old-growth forest. Condor 121(3):1-20. https://doi.org/10.1093/condor/duz031
Marzluff, J. M., J. J. Millspaugh, P. Hurvitz, and M. S. Handcock. 2004. Relating resources to a probabilistic measure of space use: forest fragments and Steller’s Jays. Ecology 85(5):1411-1427. https://doi.org/10.1890/03-0114
McGarigal, K., S. A. Cushman, and E. Ene. 2012. FRAGSTATS v4: spatial pattern analysis program for categorical and continuous maps. University of Massachusetts, Amherst, Massachusetts, USA. http://www.umass.edu/landeco/research/fragstats/fragstats.html
Mehta, P., J. Kulkarni, S. Mukherjee, S. Chavan, and A. V. Anand. 2017. A distribution survey of the Forest Owlet Heteroglaux blewitii in north-western Maharashtra. Indian BIRDS 13(4):103-108. https://indianbirds.in/pdfs/IB_13_4_MehtaETAL_ForestOwlet.pdf
Mehta, P., J. Kulkarni, and D. Patil. 2008. A survey of the critically endangered Forest Owlet Heteroglaux blewitti in Central India. BirdingASIA 10:77-87.
Mehta, P., J. Kulkarni, D. Patil, P. Kolte, and P. Khatavkar. 2007. A survey of critically endangered Forest Owlet (Heteroglaux blewitti) in Central India. Final report. Envirosearch, Pune, Maharashtra, India.
Mehta, P., J. Kulkarni, S. Talmale, and R. Chandarana. 2018. Diets of sympatric Forest Owlets, Spotted Owlets, and Jungle Owlets in East Kalibhit forests, Madhya Pradesh, India. Journal of Raptor Research 52(3):338-348. https://doi.org/10.3356/JRR-17-00002.1
Mehta, P., N. S. Prasanna, A. K. Nagar, and J. Kulkarni. 2015. Occurrence of Forest Owlet Heteroglaux blewitti in Betul District, and the importance of its conservation in the Satpura landscape. Indian BIRDS 10(6):157-159. https://indianbirds.in/pdfs/IB_10_6_MehtaETAL_ForestOwlet.pdf
Mukherjee, S., V. V. Robin, P. Mehta, and P. Koparde. 2016. Determining taxonomic and conservation status of the critically endangered Forest Owlet (Heteroglaux blewitti). Sálim Ali Centre for Ornithology and Natural History, Tamil Nadu, India.
My Mobile Tools Dev. 2018. Version 1.5. Alphabet Inc., Menlo Park, California, USA. https://play.google.com/store/apps/details?id=com.tsang.alan.lightmeter&hl=en_IN&gl=US
National Aeronautics and Space Administration Jet Propulsion Laboratory (NASA JPL). 2013. NASA shuttle radar topography mission global 3 arc second. NASA EOSDIS Land Processes DAAC. United States Geological Survey, Reston, Virginia, USA. https://doi.org/10.5067/MEaSUREs/SRTM/SRTMGL3.003
National Aeronautics and Space Administration/Messaging Extraterrestrial Intelligence/Advanced Information Systems Technology (NASA/METI/AIST) Japan spacesystems and U.S./Japan Advanced Spaceborne Thermal Emission and Reflection Radiometer (ASTER) science team. 2019. ASTER global digital elevation model v003. NASA Earth Observing System Data and Information System (EOSDIS) Land Processes DAAC. United States Geological Survey, Reston, Virginia, USA. https://doi.org/10.5067/ASTER/ASTGTM.003
National Remote Sensing Centre (NRSC). 2014. Land Use/Land Cover database on 1:50,000 scale, Natural Resources Census Project, LUCMD, LRUMG, RSAA, National Remote Sensing Centre, ISRO, Hyderabad, India.
O’Reilly, E., R. D. Gregory, A. Aunins, L. Brotons, T. Chodkiewicz, V. Escandell, R. P. B. Foppen, A. Gamero, S. Herrando, F. Jiguet, J. A. Kålås, J. Kamp, A. Klvaňová, A. Lehikoinen, Å. Lindström, D. Massimino, I. J. Øien, J. Reif, E. Šilarová, N. Teufelbauer, S. Trautmann, C. V. Turnhout, T. Vikstrøm, P. Voříšek, and S. J. Butler. 2022. An assessment of relative habitat use as a metric for species’ habitat association and degree of specialization, Ecological Indicators 135:108521. https://doi.org/10.1016/j.ecolind.2021.108521
Pande, S., A. Pawashe, M. N. Mahajan, and A. Mahabal. 2006. Changing nest site preference for holes in earth cuttings in Spotted Owlet Athene brama. Indian BIRDS 2(1):7-8. https://indianbirds.in/pdfs/IB2.1_PandeETAL_SpottedOwlet.pdf
Pande, S., A. Pawashe, M. N. Murlidhar, C. Joglekar, and A. Mahabal. 2007. Effect of food and habitat on breeding success in Spotted Owlets (Athene brama) nesting in villages and rural landscapes in India. Journal of Raptor Research 41(1):26-34. https://doi.org/10.3356/0892-1016(2007)41[26:EOFAHO]2.0.CO;2
Patel, K. 2018. In search of the Forest Owlet. Saevus, March-May 2018 edition.
Patel, J. R., S. B. Patel, S. C. Rathor, J. A. Patel, P. B. Patel, and A. G. Vasava. 2015. New distribution record of the Forest Owlet Heteroglaux Blewitti Hume, 1873, (Aves: Strigiformes: Strigidae) in Purna Wildlife Sanctuary, Gujarat, India. Journal of Threatened Taxa 7(12):7940-7944. https://doi.org/10.11609/JoTT.o4237.7940-4
Patel, J., A. Vasava, and N. Patel. 2017. Occurrence of the Forest Owlet Heteroglaux blewitti in Navsari and Valsad Districts of Gujarat, India. Indian BIRDS 13(3):78-79. https://indianbirds.in/pdfs/IB_13_3_PatelETAL_ForestOwlet.pdf
Pérez-Granados, C., D. Bustillo-de la Rosa, J. Gómez-Catasús, A. Barrero, I. Abril- Colón, and J. Traba. 2018. Autonomous recording units as effective tool for monitoring of the rare and patchily distributed Dupont’s Lark Chersophilus duponti. Ardea 106(2):139-146. https://doi.org/10.5253/arde.v106i2.a6
Quantum Geographic Information System (QGIS) Development Team. 2019. QGIS. Open Source Geospatial Foundation, Beaverton, Oregon, USA. http://qgis.org
R Core Team. 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Raha, B., R. Gadgil, and S. Bhoye. 2017. Sighting of the Forest Owlet Heteroglaux blewitti in Harsul, Nashik District, Maharashtra. Indian BIRDS 13(3):80-81. https://indianbirds.in/pdfs/IB_13_3_RahaETAL_ForestOwlet.pdf
Reddy, C. S., C. S. Jha, V. K. Dadhwal, P. Hari Krishna, S. V. Pasha, K. V. Satish, K. Dutta, K. R. L. Saranya, F. Rakesh, G. Rajashekar, and P. G. Diwakar. 2016. Quantification and monitoring of deforestation in India over eight decades (1930-2013). Biodiversity and Conservation 25:93-116. https://doi.org/10.1007/s10531-015-1033-2
Rota, C. T., M. A. R. Ferreira, R. W. Kays, T. D. Forrester, E. L. Kalies, W. J. McShea, A. W. Parsons, and J. J. Millspaugh. 2016. A multispecies occupancy model for two or more interacting species. Methods in Ecology and Evolution 7(10):1164-1173. https://doi.org/10.1111/2041-210X.12587
Roy, P. S., A. Roy, P. K. Joshi, M. P. Kale, V. K. Srivastava, S. K. Srivastava, R. S. Dwevidi, C. Joshi, M. D. Behera, P. Meiyappan, Y. Sharma, A. K. Jain, J. S. Singh, Y. Palchowdhuri, R. M. Ramachandran, B. Pinjarla, V. Chakravarthi, N. Babu, M. S. Gowsalya, P. Thiruvengadam, M. Kotteeswaran, V. Priya, K. M. V. N. Yelishetty, S. Maithani, G. Talukdar, I. Mondal, K. S. Rajan, P. S. Narendra, S. Biswal, A. Chakraborty, H. Padalia, M. Chavan, S. N. Pardeshi, S. A. Chaudhari, A. Anand, A. Vyas, M. K. Reddy, M. Ramalingam, R. Manonmani, P. Behera, P. Das, P. Tripathi, S. Matin, M. L. Khan, O. P. Tripathi, J. Deka, P. Kumar, and D. Kushwaha. 2015. Development of decadal (1985-1995-2005) land use and land cover database for India. Remote Sensing 7:2401-2430. https://doi.org/10.3390/rs70302401
Strickler, G. S. 1959. Use of the densiometer to estimate density of forest canopy on permanent sample plots. U.S. Forest Service, Pacific Northwest Forest and Range Experiment Station, Washington, D.C., USA.
Vanitha, V., K. Thiyagesan, N. Baskaran. 2018. Does southern Spotted Owlet Athene brama brama (Temminck, 1821) serve as a biocontrol agent of agricultural pests? A case study from Cauvery Deltaic region of southern India. Pages 139-149 in C. Sivaperuman and K. Venkataraman, editors. Indian hotspots. Springer, Singapore, Asia. https://doi.org/10.1007/978-981-10-6983-3_7
Watson, J. E. M., R. J. Whittaker, and T. P. Dawson. 2004. Habitat structure and proximity to forest edge affect the abundance and distribution of forest-dependent birds in tropical coastal forests of southeastern Madagascar. Biological Conservation 120(3):311-327. https://doi.org/10.1016/j.biocon.2004.03.004
Wiens, J. D., R. G. Anthony, and E. D. Forsman. 2014. Competitive interactions and resource partitioning between northern Spotted Owls and Barred Owls in western Oregon. Wildlife Monographs 185(1):1-50. https://doi.org/10.1002/wmon.1009
Fig. 1
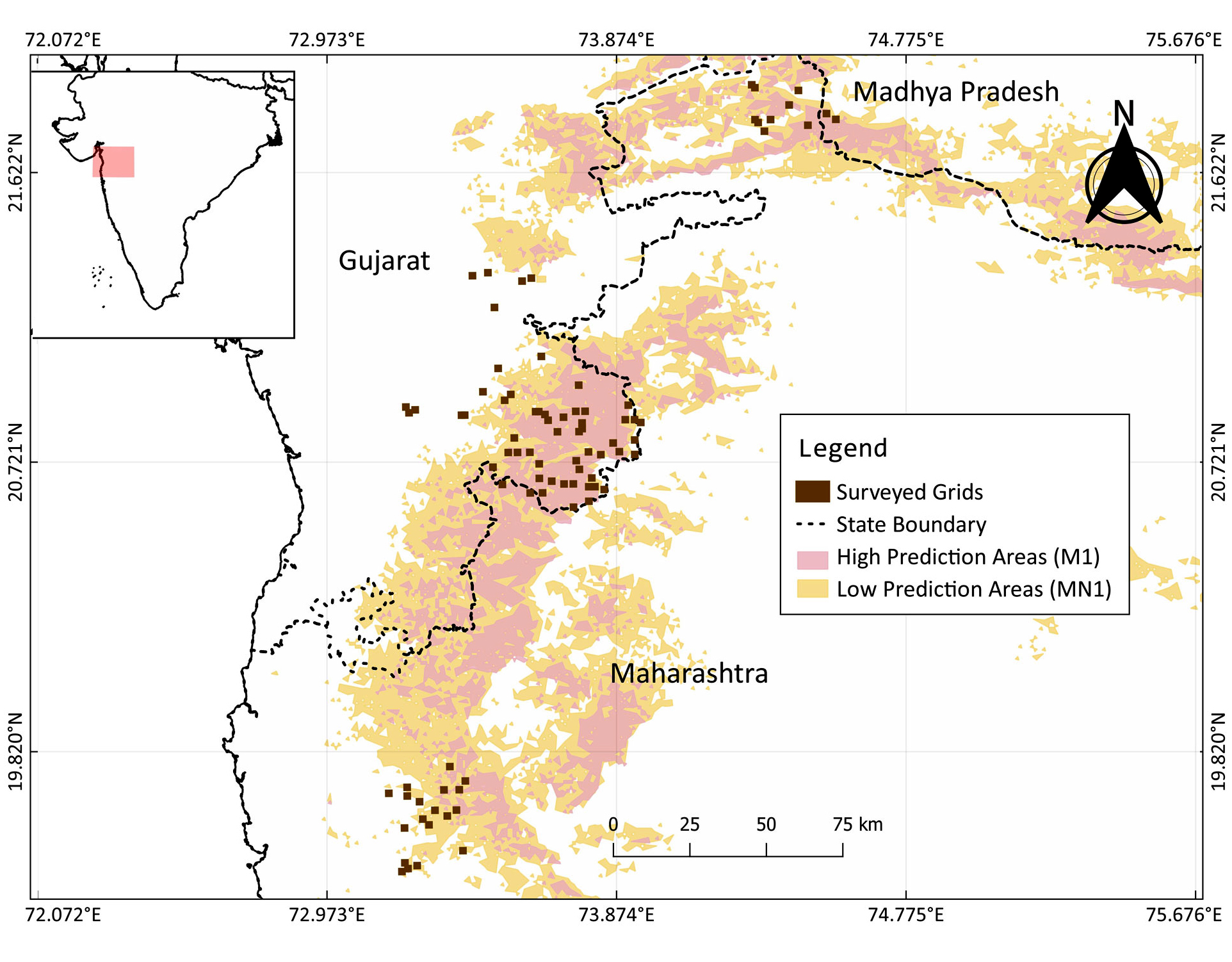
Fig. 1. Map representing 88 survey locations (2018-2019) spread across two states namely: Gujarat and Maharashtra based on the species distribution models (SDMs) for Forest Owlet, Athene blewitti, (Mukherjee et al. 2016, Koparde 2019). Location of the study area (pink polygon) is indicated in the inset map of India.
Fig. 2
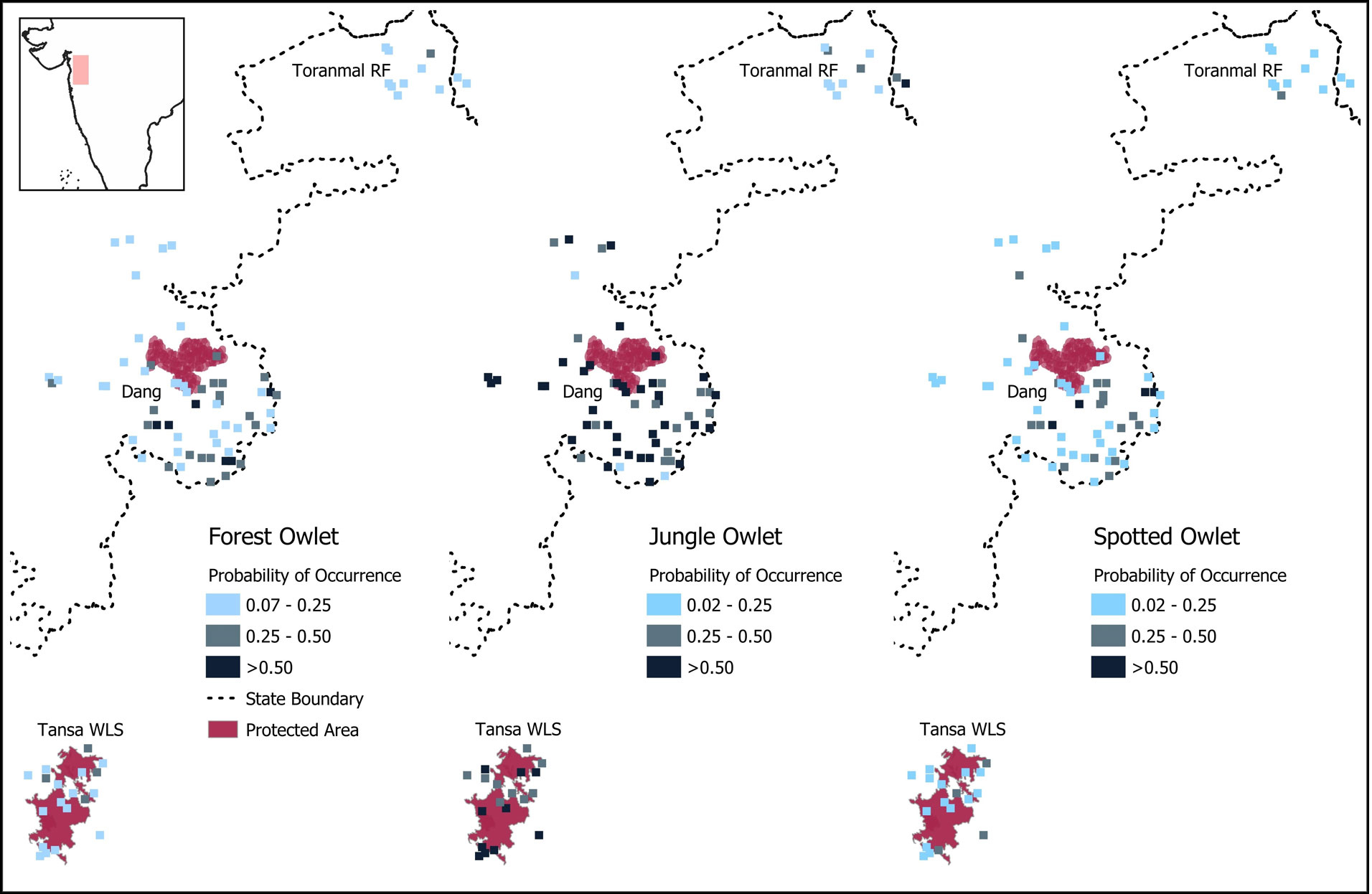
Fig. 2. A comparison of individual site occupancy (ψ) estimates of the three owlet species: Forest Owlet (Athene blewitti), Jungle Owlet (Glaucidium radiatum), and Spotted Owlet (Athene brama) in the study sites in Gujarat and Maharashtra.
Fig. 3
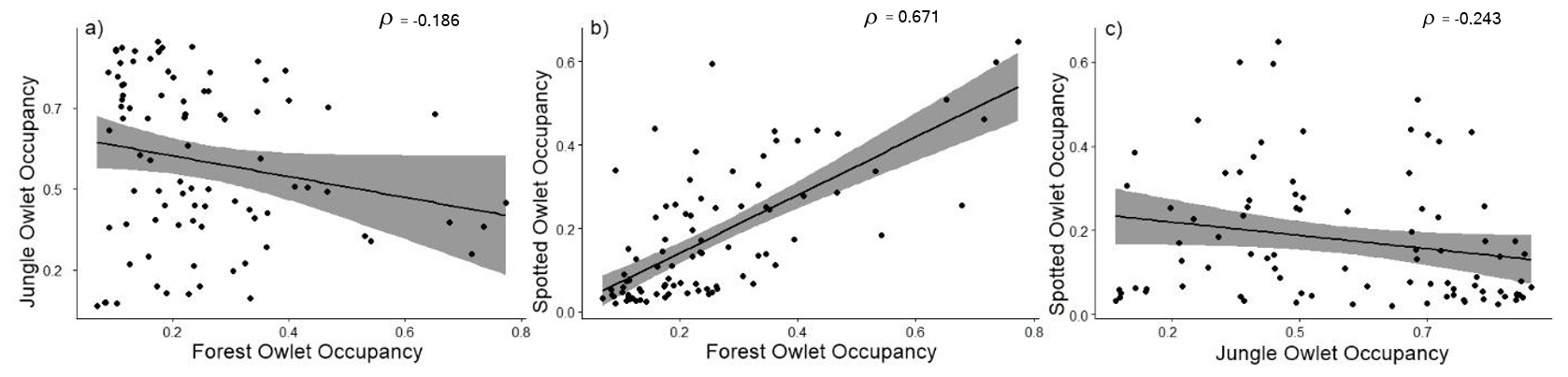
Fig. 3. Relationship between a) Forest Owlet, Athene blewitti, and Jungle Owlet, Glaucidium radiatum, b) Forest Owlet and Spotted Owlet, Athene brama, c) Jungle Owlet and Spotted Owlet as a function of the occupancy probability. Each point corresponds to cell-wise occupancy estimates of the three species.
Fig. 4
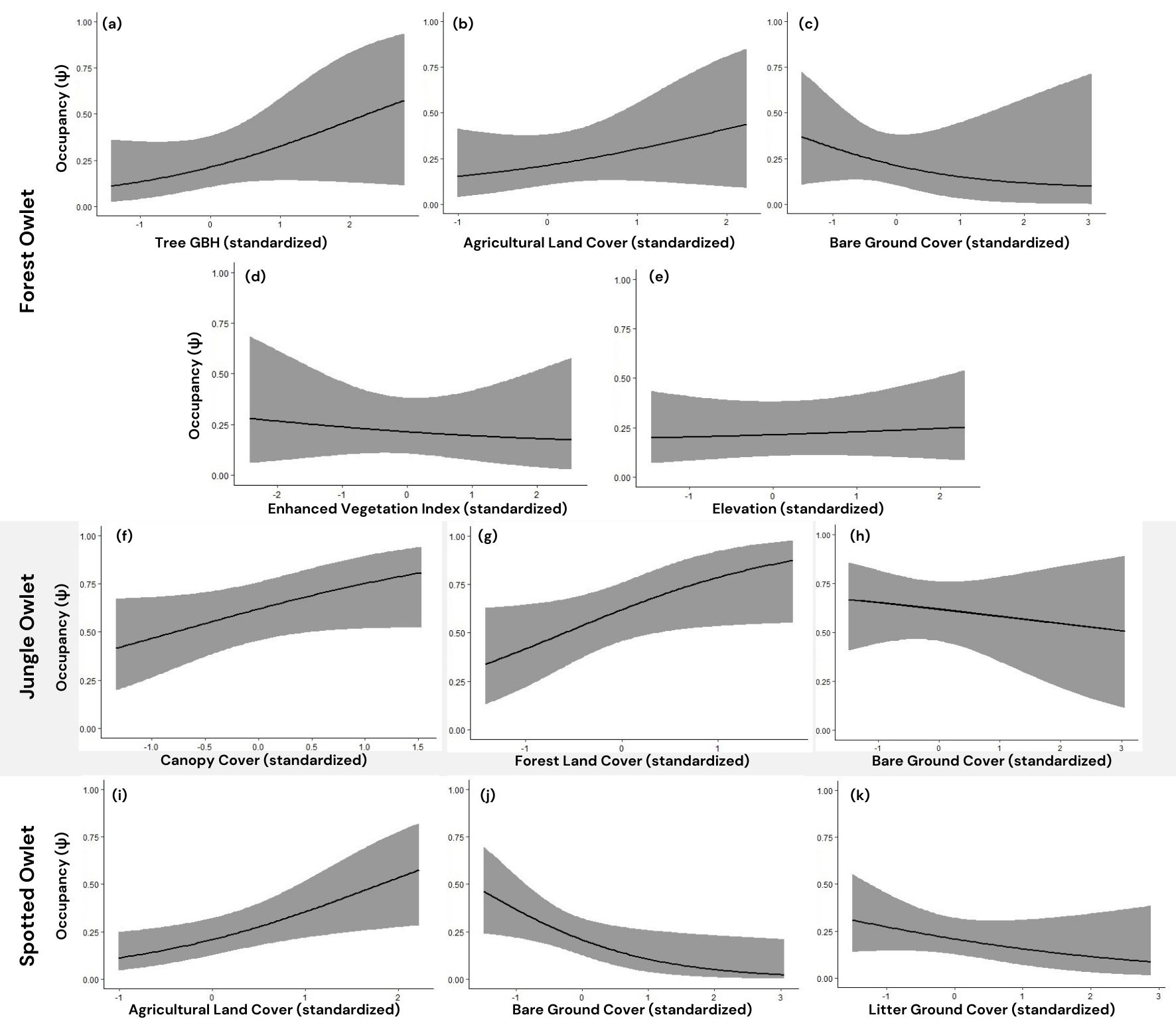
Fig. 4. Model-averaged predicted occupancy in response to the covariates that are present in the top models for Forest Owlet, Athene brama: (a) tree girth at breast height (cm), (b) agricultural land cover (%), (c) bare ground cover (%), (d) enhanced vegetation index, (e) elevation (msl), Jungle Owlet, Glaucidium radiatum: (f) canopy cover (%), (g) forest land cover (%), (h) bare ground cover (%), and Spotted Owlet, Athene brama: (i) agricultural land cover (%), (j) bare ground cover (%), (k) litter ground cover (%). All other variables are set to mean values. The predicted probability of occupancy is plotted over the observed range of values of each predictor. Bands represent 95% confidence interval.
Table 1
Table 1. Predicted influence of site level covariates on the occupancy probability and survey level covariates on the detection probability of the three owlets: Forest Owlet (Athene blewitti), Jungle Owlet (Glaucidium radiatum), and Spotted Owlet (Athene brama). The “+” signifies a positive effect on the response variable, and “–” signifies a negative effect on the response variable.
| Data source | Covariate | Predicted effect on occupancy of Forest Owlet† | Predicted effect on occupancy of Jungle Owlet | Predicted effect on occupancy of Spotted Owlet | |
| Site covariates | Primary | Average tree girth at breast height (GBH) | + | + | + |
| Percent canopy cover (CC) | - | + | - | ||
| Percent bare ground cover (BC) | - | - | - | ||
| Percent litter cover (LC) | + | - | - | ||
| Percent rock cover (RC) | + | + | |||
| Derived | Percent forest cover (FC) | + | |||
| Percent grassland cover (GC) | + | + | |||
| Percent agricultural land cover (AC) | + | + | |||
| Percent human settlement (HS) | + | ||||
| Average topographic ruggedness index (TRI) | + | ||||
| Enhanced vegetation index (EVI) | + | ||||
| Average elevation (ELE) | + | - | |||
| Forest edge density (FED) | + | + | |||
| Agricultural edge density (AED) | + | ||||
| Drainage density (DD) | + | ||||
| Expected effect on detection of Forest Owlet | Expected effect on detection of Jungle Owlet | Expected effect on detection of Spotted Owlet | |||
| Survey-specific covariates | Primary | Noise (N) | - | - | - |
| Ambient light (AL) | + | + | - | ||
Table 2
Table 2. Summary of the top ranked models according to quasi Akaike information criterion (QAICc) values for Forest Owlet (Athene blewitti), Jungle Owlet (Glaucidium radiatum), and Akaike information criterion (AICc) values for Spotted Owlet (Athene brama) are represented below. (GBH: girth at breast height, CC: canopy cover, AC: agricultural land cover, FC: forest land cover, BC: bare ground cover, LC-: litter cover, ELE: elevation, EVI: enhanced vegetation index, N: noise, AL: ambient light). ψ(.)p(.) is the null model.
| Species | Model | QAICc | ΔQAICc | AIC wgt | Model Likelihood | no. Par. | -2*LogLike |
| Forest Owlet | ψ(GBH+AC)p(N) | 89.18 | 0.00 | 0.2001 | 1.0000 | 5 | 167.47 |
| ψ(GBH+BC+AC)p(N) | 89.29 | 0.11 | 0.1894 | 0.9465 | 6 | 162.59 | |
| ψ(GBH+EVI)p(N) | 90.19 | 1.01 | 0.1208 | 0.6035 | 5 | 169.63 | |
| ψ(GBH+BC+EVI)p(N) | 90.30 | 1.12 | 0.1143 | 0.5712 | 6 | 164.76 | |
| ψ(GBH+BC+ELE+AC)p(N) | 90.66 | 1.48 | 0.0955 | 0.4771 | 7 | 160.24 | |
| ψ(.)p(.) | 90.75 | 1.57 | 0.0913 | 0.4561 | 2 | 185.27 | |
| ψ(GBH+BC+EVI+AC)p(N) | 91.32 | 2.14 | 0.0686 | 0.3430 | 7 | 161.65 | |
| Jungle Owlet | ψ(CC+FC),p(AL) | 230.12 | 0.00 | 0.5608 | 1.0000 | 5 | 347.08 |
| ψ(CC+BC+FC),p(AL) | 231.50 | 1.38 | 0.2813 | 0.5016 | 6 | 345.82 | |
| ψ(CC+BC+FC+ELE),p(AL) | 233.53 | 3.41 | 0.1019 | 0.1818 | 7 | 345.52 |
|
| Model | AICc | ΔAICc | AIC wgt | Model Likelihood | no. Par. | -2*LogLike | |
| Spotted Owlet | ψ(BC+LC+AC),p(AL) | 154.44 | 0 | 0.3941 | 1 | 6 | 139.33 |
| ψ(BC+AC),p(AL) | 157.39 | 2.95 | 0.0902 | 0.2288 | 5 | 145.25 | |
Table 3
Table 3. Model-specific β coefficient estimates (from top ranking models) for covariates determining Forest Owlet, Athene blewitti (FO), Jungle Owlet, Glaucidium radiatum (JO), and Spotted Owlet, Athene brama (SO), occupancy ψ in Gujarat and Maharashtra. Number of sites = 88. (GBH: girth at breast height, CC: canopy cover, AC: agricultural land cover, FC: forest land cover, BC: bare ground cover, LC-: litter cover, ELE: elevation, EVI: enhanced vegetation index, N: noise, AL: ambient light). ψ(.)p(.) is the null model.
| Species | Models | β (SE) | βGBH (SE) | βBC (SE) | βEVI (SE) | βCC (SE) | βELE (SE) | βAC (SE) | βFC (SE) | βLC (SE) | βN (SE) | βAL (SE) |
| FO | ψ(GBH+AC)p(N) | -2.5098 (0.5498) | 0.2409 (0.1159) |
- | - | - | - | 0.1504 (0.0869) |
- | - | -0.5784 (0.2538) |
- |
| FO | ψ(GBH+BC+AC)p(N) | -1.7689 (0.6379) | 0.2430 (0.1227) |
-0.3329 (0.1692) |
- | - | - | 0.2221 (0.0999) |
- | - | -0.5844 (0.2552) |
- |
| FO | ψ(GBH+EVI)p(N) | -0.0474 (2.4664) | 0.3095 (0.1113) |
- | -0.2818 (0.3141) |
- | - | - | - | - | -0.5783 (0.2542) |
- |
| FO | ψ(GBH+BC+EVI)p(N) | 3.7723 (3.1473) | 0.3379 (0.1202) |
-0.3139 (0.1508) |
-0.6636 (0.3850) |
- | - | - | - | - | -0.5901 (0.2574) |
- |
| FO | ψ(GBH+BC+ELE+AC)p(N) | -2.3087 (0.7580) | 0.2162 (0.1271) |
-0.3877 (0.1764) |
- | - | 0.1728 (0.1133) |
0.2314 (0.1036) |
- | - | -0.5960 (0.2594) |
- |
| JO | ψ(CC+FC),p(AL) | -1.6058 (0.5098) |
- | - | - | 0.1755 (0.0802) |
- | - | 0.2916 (0.0975) |
- | - | 0.1901 (0.1639) |
| JO | ψ(CC+BC+FC),p(AL) | -0.8839 (0.8009) |
- | -0.1595 (0.1444) |
- | 0.1847 (0.0828) |
- | - | 0.2269 (0.1086) |
- | - | 0.1905 (0.1640) |
| SO | ψ(BC+LC+AC),p(AL) | 0.1556 (1.0151) | - | -0.4525 (0.2103) | - | - | - | 0.2808 (0.1221) | - | -0.4792 (0.2309) | - | -0.3940 (0.2471) |