







The following is the established format for referencing this article:
Clarke, J. R., T. J. Hovick, and K. Sedivec. 2026. Breeding-site fidelity of an obligate grassland bird in the Northern Great Plains. Journal of Field Ornithology 97(2):9.ABSTRACT
Many migratory birds show breeding-site fidelity, but research has focused primarily on shrubland and forest species, with grassland birds largely overlooked. Fidelity is typically higher in woody systems because of the greater predictability and stability of these environments. Furthermore, breeding-site fidelity has been shown to influence territory size, with returning birds occupying higher-quality habitat in familiar landscapes and maintaining smaller territories. We investigated Western Meadowlark (Sturnella neglecta) breeding-site fidelity in the Northern Great Plains from 2022 to 2024, with objectives to (1) quantify breeding-site fidelity in grasslands managed with fire and grazing, and (2) characterize the relationship between individual breeding-site fidelity and territory size. We observed high breeding-site fidelity, with annual return rates of 61% in 2023 and 60% in 2024, and 33% of birds banded in 2022 returning in both subsequent years. Territory sizes declined over time regardless of return status (2022: mean = 7.80 ha; 2024: mean = 5.50 ha; χ²₂ = 7.86, p = 0.02). Mean territory size did not differ by return status (χ²₂ = 0.53, p = 0.77) and variability in territory size was not different between returning and non-returning birds (F1,68 = 3.39, p = 0.07). We documented strong breeding-site fidelity in Western Meadowlarks. Although the drivers of this fidelity remain uncertain, several non-exclusive mechanisms may contribute, including stable habitat maintained through fire and grazing, individual experience, and landscape configuration that limits dispersal. Declines in meadowlark territory size over time may reflect interannual variation in precipitation. Precipitation likely influences resource availability, with wetter years allowing birds to establish smaller, more resource-rich territories. Our findings contribute additional evidence that grassland birds can exhibit strong breeding-site fidelity despite dynamic management and environmental conditions and highlight the need for future research explicitly linking management regimes, environmental variability, and individual behavior.
RESUMEN
Muchas aves migratorias exhiben fidelidad a los sitios de cría, pero las investigaciones se han enfocado principalmente en especies de arbustales y bosques, siendo las aves de pastizales generalmente pasadas por alto. La fidelidad es típicamente mayor en sistemas boscosos dada la mayor predictibilidad y estabilidad de estos ambientes. Además, se ha demostrado que la fidelidad a los sitios de cría influye sobre los tamaños de los territorios, ya que las aves que retornan ocupan hábitats de mayor calidad en paisajes conocidos y mantienen territorios más pequeños. Investigamos la fidelidad a los sitios de cría en el Pradero del Oeste (Sturnella neglecta), con los objetivos de (1) cuantificar la fidelidad a los sitios de cría en pastizales manejados con fuego y pastoreo, y (2) caracterizar la relación entre la fidelidad individual a los sitios de cría y el tamaño territorial. Observamos una elevada fidelidad a los sitios de cría, con tasas anuales de retorno del 61% en 2023 y 60% en 2024, y con el 33% de las aves marcadas en 2022 retornando en los años siguientes. Los tamaños territoriales disminuyeron a lo largo del tiempo independientemente de la condición de retorno (2022: media=7,80 ha; 2024: media: 5,50 ha; χ ²₂ = 7.80, p = 0.02). El tamaño medio del territorio no difirió en función de la condición de retorno (χ²₂ = 0.53, p = 0.77), y la variabilidad en el tamaño territorial tampoco varió entre aves que retornaban y aquellas que no (F1,68 = 3.39, p = 0.07). Documentamos una fuerte fidelidad a los sitios de cría en el Pradero del Oeste. Si bien los factores que determinan esta fidelidad permanecen inciertos, varios mecanismos no excluyentes pueden contribuir, incluyendo la estabilidad del hábitat mantenida mediante fuego y pastoreo, la experiencia individual y una configuración del paisaje que limita la dispersión. La disminución en el tamaño de los territorios a lo largo del tiempo en esta especie podría reflejar variaciones interanuales en las precipitaciones. Es probable que las precipitaciones influyan en la disponibilidad de los recursos, permitiendo que, en años más húmedos, las aves establezcan territorios más pequeños, pero con mayor disponibilidad de recursos. Nuestros hallazgos aportan evidencia adicional acerca de la marcada fidelidad a los sitios de cría que las aves de pastizal pueden exhibir a pesar de los manejos dinámicos y las condiciones ambientales, y resaltan la necesidad de futuras investigaciones que vinculen explícitamente los regímenes de manejo, la variabilidad ambiental y el comportamiento individual.
INTRODUCTION
Many migratory birds demonstrate fidelity to their breeding sites, often returning to the same location year after year (Gill et al. 2006, Schlossberg 2009, Groce et al. 2010). Breeding-site fidelity has often been attributed to reproductive success, where successful individuals from the previous year return to the same areas (Hoover 2003, Buehler et al. 2008). Forest and shrubland birds often demonstrate high breeding-site fidelity, but trends are less consistent with grassland birds (Green 1992, Murphy 1996, Porneluzi 2003, Wenny 2003, Gill et al. 2006, Jones et al. 2007, Stauffer 2008, Fajardo et al. 2009, Schlossberg 2009, Small et al. 2012, Bozzo 2023). In grasslands, vegetation composition and structure vary both temporally and spatially because of frequent but patchy disturbance including grazing, fire, and drought (Higgins 1984, Igl and Johnson 1999, Fuhlendorf and Engle 2001). A substantial change in vegetation structure at large spatial scales can change habitat quality, reducing the advantages of returning to previously successful breeding sites (Gill et al. 2006, Ortega et al. 2006). Despite its importance, breeding-site fidelity in grassland birds remains poorly understood. This research aims to fill this gap by providing new insights into breeding-site fidelity patterns for grassland birds.
Grazing practices designed to benefit grassland birds often emphasize achieving variability in vegetation structure (McNew et al. 2015, Duchardt et al. 2016). By implementing historic disturbance regimes, specifically the interacting effects of fire and grazing, land managers can restore the spatial and temporal variability that once characterized these systems (Fuhlendorf et al. 2006). The resulting mosaic of shifting vegetation structure and composition has been shown to stabilize avian community dynamics, despite high levels of within-patch turnover (Hovick et al. 2015). Similarly, management that maintains shifting habitat conditions across years can sustain site use by grassland birds, even though individuals may alter territories slightly in response to changes in vegetation structure (Dechant et al. 2002, Gill et al. 2006). Maintaining dynamic habitat variability is therefore essential to sustain the ecological complexity that underpins grassland bird diversity.
Although many grassland birds exhibit low breeding-site fidelity in response to dynamic and unpredictable habitat conditions (Murray 1969, Jones et al. 2007), some show a remarkable capacity to return to previously used sites (Bollinger and Gavin 1989, Gill et al. 2006, Ingold et al. 2010). North American grasslands east of the Mississippi River are small and highly fragmented, functioning as islands within a forested landscape (Askins et al. 2007). Grassland bird return rates during the breeding season are correspondingly higher, ranging from 19 to 83% (Bédard and LaPointe 1984, Bollinger and Gavin 1989, Balent and Norment 2003, Gill et al. 2006, Stauffer 2008, Fajardo et al. 2009, Ingold et al. 2010, Small et al. 2012). In contrast, the more contiguous grasslands of the Great Plains in western North America offer abundant grassland environments and show lower return rates, ranging from 2 to 49% (Best 1977, Colwell and Oring 1989, Zimmerman and Finck 1989, Green 1992, Fletcher et al. 2006, Jones et al. 2007, Bozzo 2023). Lower breeding-site fidelity in western grasslands may reflect the greater challenge birds have in relocating specific territories within expansive and highly dynamic grassland landscapes.
Breeding-site fidelity may also influence territory size, a metric that has been used as a proxy for habitat quality (Wiens et al. 1985, Whitaker and Warkentin 2010, Cooper et al. 2014, Diemer and Nocera 2014). Because territories encompass multiple functions, including mate attraction, foraging, breeding, and rearing young (Hinde 1956), territory size can reflect the overall suitability of a site for supporting reproductive success. Smaller territories are typically found in higher-quality areas where resources are more abundant and accessible (Diemer and Nocera 2014). Consistent with this pattern, older birds with more experience often return to familiar, high-quality areas and establish smaller, clustered territories (Beauchamp et al. 1997, Nocera et al. 2009). In contrast, younger or less experienced individuals typically settle in surrounding lower-quality areas and establish larger territories where resources are more limited (Nocera et al. 2009).
As a declining, yet still locally abundant, obligate grassland bird, the Western Meadowlark (Sturnella neglecta; hereafter meadowlark) provides a valuable model for studying breeding-site fidelity and habitat use in dynamic grassland systems. Meadowlarks are ground-nesting birds of conservation concern in North Dakota, having declined by over 60% since the 1970s (Dyke et al. 2015, Davis and Lanyon 2020). Despite these declines, they remain relatively abundant across all three grazing treatments in our study area (Duquette 2020), providing an opportunity to examine patterns of breeding-site fidelity and territory dynamics. To date, only one peer-reviewed study has documented breeding-site fidelity in this species. In Wisconsin’s smaller and more fragmented grasslands, 57% of males and 53% of females returned at least once over three years, with 92% of returning males reoccupying the same territory (Lanyon 1957). However, some individuals may return over even longer timescales, as suggested by a recaptured meadowlark six years after banding in Colorado (Klimkiewicz and Futcher 1989). Reported territory sizes across the species’ range vary widely, from 1 to 13 ha (Kendeigh 1941, Lanyon 1956, Laubach 1984), but studies in mixed-grass prairie have documented a narrower range (2–7 ha; Wiens 1970, 1971, Schaeff and Picman 1988).
Our objectives were to (1) quantify breeding-site fidelity of meadowlarks in grasslands managed with fire and grazing, and (2) characterize the relationship between individual breeding-site fidelity and territory size as a potential indicator of experience-related space use. The limited scope of research on this species, and on grassland birds more broadly, leaves important gaps in our understanding. By linking return rates with territory size, our study addresses this gap and advances understanding of how individual experience may influence habitat selection in dynamic grassland systems.
METHODS
Study area
Our research was conducted at the Central Grasslands Research Extension Center (CGREC) in Streeter, North Dakota, located along the border between Kidder and Stutsman counties (46°42′56″ N, 99°27′08″ W). This region has a temperate, continental climate with an average growing season (April to September) precipitation of 433 mm and average temperatures of 13.6 °C (NDAWN 2025). However, precipitation showed substantial variability during the study period, with mean growing season totals of 343 mm in 2022, 385 mm in 2023, and 596 mm in 2024 (NDAWN 2025, Fig. 1).
The site is classified as northern mixed-grass prairie and is characterized by a mix of cool- and warm-season grasses and forbs, with vegetation communities increasingly dominated by non-native, invasive cool-season grasses such as Kentucky bluegrass (Poa pratensis Leyss.) and smooth brome (Bromus inermis L.). Woody vegetation is patchy and dominated by western snowberry (Symphoricarpos occidentalis Hook.). We utilized twelve 65-hectare pastures, each assigned to one of three grazing treatments: season-long grazing, patch-burn grazing, or modified twice-over rest-rotation grazing, with four replicates per treatment. Stocking rates were consistent across treatments and managed to maintain approximately 40 to 50% average degree of disappearance across the unit. A full description of the experimental design is provided in Duquette et al. (2022a).
Territory mapping
Each year, we captured and banded meadowlarks using target mist-netting with audio lures. Meadowlarks lack clear post-fledging age indicators, so all adult birds were determined to be in their definitive cycle and basic plumage (Pyle 1997). Sex was determined with a combination of morphological measurements and behavioral observations (Pyle 1997). Birds were banded with the standard U.S. Fish and Wildlife Service band and a combination of three color bands. In the second and third years, we conducted surveys in late May to relocate previously banded individuals. For all banded birds, we mapped territories using a combination of flush- and spot-mapping techniques (Reed 1985). Mapping began at sunrise and concluded by approximately 14:00 hours, during which time we recorded all flush and landing sites for each bird until reaching 5–15 locations on the first day (Cooper et al. 2014, Verheijen et al. 2019). We repeated this over approximately one week to collect a total of 20–40 locations (Cooper et al. 2014, Verheijen et al. 2019). If individuals repeatedly used linear anthropogenic structures (e.g., fences, powerlines) during mapping efforts, we stopped delineation for that day and returned later to improve accuracy. This approach accounted for the possibility that such structures may be used opportunistically or transiently for territorial behaviors, rather than indicating stable territory boundaries.
Analysis
We estimated territory size (ha) using minimum convex polygons generated with the adehabitatHR package (version 0.4.22, Calenge 2006) in R version 4.4.2 (R Core Team 2025). Annual return rates were calculated as the number of birds returning in the current year divided by the number of territories delineated the previous year, as not all banded birds were relocated. Although most birds remained within project boundaries, one individual banded in 2022 returned in both 2023 and 2024 but established territories outside of the study area in both years. It was included in return rate calculations, but only its 2022 territory was used in territory size estimates. Another bird, banded after territory delineation in 2023, returned to the same area in 2024. This bird was also included in return rate calculations for both years, but only its 2024 territory was used in territory size estimates. We modeled the effects of return status (0 = non-returner, 1 = returned 1 year, 2 = returned 2 years) and year on territory size using a generalized linear mixed-effects model with glmmTMB (version 1.1.12, Brooks et al. 2017), including USGS band number as a random effect to account for repeated measures. Significance of fixed effects was assessed with a Type II Wald χ² test (car package version 3.1.3, Fox and Weisberg 2019), and pairwise comparisons were conducted with emmeans (version 1.11.2.8, Lenth 2025). Model assumptions were assessed with DHARMa (version 0.4.7, Hartig 2022), and equality of variances among return-status groups was evaluated using Levene’s test (car package version 3.1.3, Fox and Weisberg 2019).
RESULTS
We banded 50 adult male meadowlarks at the CGREC over the three-year study, with 46 establishing breeding territories (Table 1). Return rates were consistent across years: 61% of males that held territories in 2022 returned in 2023, and 60% of all territorial males from 2023 returned in 2024 (i.e., includes birds from 2022 and 2023; Table 1). In 2024, returning birds included six individuals that were originally banded in 2022 (33% of new territories in 2022) and nine that were originally banded in 2023 (64% of new territories in 2023, Table 1).
Seventy territories were delineated across the study period. Territory size ranged from 1.97 ha to 16.46 ha, with an overall mean of 6.47 ha (SD = 2.76). Territory size declined over the study period (2022–2024; χ² = 7.86, df = 2, p = 0.02, Fig. 2a). Estimated marginal means showed smaller territories in 2024 compared to 2022 (β = −2.32 ± 0.85 SE, p = 0.02), with no differences between 2024 and 2023 (β = −1.31 ± 0.75 SE, p = 0.20) or 2023 and 2022 (β = −1.00 ± 0.88 SE, p = 0.49). Mean territory size among birds that did not return was 6.76 ha (SD = 3.09), compared to 5.87 ha (SD = 1.98) for those that returned in one subsequent year and 6.14 ha (SD = 1.99) for those that returned in two consecutive years (Table 2). However, territory size did not differ based on return status (χ² = 0.53, df = 2, p = 0.77; Fig. 2b). Although non-returning birds exhibited greater variability in territory size (SD = 3.09) than birds that returned either one (SD = 1.98) or two (SD = 1.99) years, these differences were not significant (F1,68 = 3.39, p = 0.07, Fig. 2b).
DISCUSSION
Grassland birds evolved in dynamic landscapes impacted by fire, grazing, and variation in weather (Kirsch and Kruse 1973, Higgins 1984, Igl and Johnson 1999, Ahlering 2005). Our results add to a growing body of evidence that grassland birds exhibit high breeding-site fidelity, measured through annual return rates, in grasslands managed with historic disturbance regimes (Best 1977, Colwell and Oring 1989, Zimmerman and Finck 1989, Green 1992, Fletcher et al. 2006, Jones et al. 2007, Bozzo 2023). We documented an overall annual return rate of approximately 60% for meadowlarks in the northern Great Plains over a three-year study period. This is comparable to estimates from the eastern, more forested portions of the species’ range (57%, Lanyon 1957). Moreover, 33% of individuals returned for two consecutive years, exhibiting exceptionally strong breeding-site fidelity.
Although the drivers of the strong breeding-site fidelity we observed remain uncertain, we propose three hypotheses that may explain this pattern. First, imposed grassland structural variability from fire and grazing at the CGREC may create temporal and spatial stability (Hovick et al. 2015). Specifically, our study sites use fire and varying cattle grazing intensities to promote structural variation in the plant community (Duquette et al. 2022b). This, coupled with the inherent heterogeneity of the Prairie Pothole region, may maintain high-quality habitat over space and time that allows grassland birds to reoccupy the same general area each year (Duquette et al. 2022a, b). By sustaining spatial and temporal diversity in vegetation composition and structure through intentional management (Figge 2004, Clarke et al. 2026), managers create broadly stable landscapes that may allow grassland birds to exhibit long-term breeding-site fidelity (Doak et al. 1998, Tilman et al. 1998, Ortega et al. 2006, Gill et al. 2006). Second, individual experience may influence breeding-site fidelity if birds use information from prior breeding attempts to inform their settlement decisions (Gavin and Bollinger 1988, Hoover 2003, Porneluzi 2003). For example, individuals that successfully reproduce at a site may be more likely to return in subsequent years, whereas unsuccessful breeders may disperse to seek better conditions (Hoover 2003). Third, the landscape configuration of our study sites may create a perceived “oasis effect” where the intact grasslands around the CGREC stand out from the more intensive agricultural landscapes located in the immediate vicinity and the eastern portions of North Dakota. Limited availability of suitable habitat, combined with the area sensitivity of grassland birds, may constrain dispersal and increase the likelihood that individuals return to larger, intact patches like the CGREC (Balent and Norment 2003, Ribic et al. 2009).
Territory size declined from the beginning to the end of the study period, regardless of return status. The interannual decline in territory size may be driven by changing environmental conditions associated with precipitation. Our study area experienced below-average precipitation in 2022 (343 mm) and 2023 (385 mm) with markedly wetter conditions in 2024 (596 mm; NDAWN 2025). In water-limited systems like the Great Plains, increased precipitation boosts primary productivity (Petrie et al. 2018), improving both nest concealment and foraging opportunities for ground-nesting birds (Klug et al. 2010, Marta et al. 2021). Optimal territory sizes change in response to these shifting conditions where years with greater resource availability (i.e., wetter years with more production) likely result in smaller territories (Zammarelli et al. 2024).
CONCLUSION
The high annual return rates we observed in meadowlarks have rarely been recorded in western grassland systems and suggest strong breeding-site fidelity (Bédard and LaPointe 1984, Bollinger and Gavin 1989, Balent and Norment 2003, Gill et al. 2006, Stauffer 2008, Fajardo et al. 2009, Ingold et al. 2010, Small et al. 2012). We propose several competing hypotheses that could explain the patterns we observed but cannot definitively state why meadowlarks had such high return rates during this study. Nevertheless, our findings suggest that meadowlarks can exhibit strong breeding-site fidelity within actively managed grassland systems. Previous research in the Great Plains has demonstrated that management approaches restoring spatial and temporal variability in vegetation structure can influence grassland bird habitat use across multiple life-history stages and affects stability and turnover in grassland bird communities (Fuhlendorf et al. 2009, Hovick et al. 2015, 2017a, b, Davis et al. 2016). Furthermore, as climate becomes more variable due to more frequent and prolonged droughts (Parmesan and Yohe 2003, Stott 2016, Ojima et al. 2021) and anthropogenic drivers continue to alter land use and landscape configuration (Wright and Wimberly 2013), the availability and predictability of critical resources in grassland systems are increasingly threatened (Brook et al. 2008, Mantyka‐Pringle et al. 2013). In the case of grassland birds, this will create unpredictable environments that could affect stability and population growth. Collectively, our findings contribute additional information on breeding-site fidelity in grassland birds and underscore the need for future work that explicitly links management regimes, environmental variability, and individual behavior to patterns of breeding-site fidelity and long-term habitat use.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Justin R. Clarke: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review and editing, Torre J. Hovick: Conceptualization, Writing – review and editing, Supervision, Kevin Sedivec: Funding acquisition, Resources, Writing – review and editing.
ACKNOWLEDGMENTS
We thank Evalynn Trumbo-Clarke for assisting in training technicians in proper banding procedures. Additionally, we thank all the technicians who helped band and spot-map territories. Without them, this work could not have been done. Resources for this project were purchased using funding from Central Grasslands Research Extension Center and the Robert H. Levis II Cross Ranch Fellowship.
DATA AVAILABILITY
The data and R scripts used in this analysis are publicly available in Figshare repositories. GPS data used to calculate territory sizes can be accessed at https://doi.org/10.6084/m9.figshare.29160797, and the R scripts used to make figures and analyses are available at https://doi.org/10.6084/m9.figshare.29159819.
This work was conducted under USGS banding permit #24312 and North Dakota Game and Fish permits OLN05753725, OLN06223604, and OLN06333702 for 2022, 2023, and 2024. All procedures were approved by the Institutional Animal Care and Use Committee under permit #A21062.
LITERATURE CITED
Ahlering, M. A. 2005. Settlement cues and resource use by Grasshopper Sparrows and Baird’s Sparrows in the upper Great Plains. Dissertation. University of Missouri - Columbia, Columbia, Missouri, USA. https://www.proquest.com/docview/305449072/abstract/29D1BB99C8064CD1PQ/1
Askins, R. A., F. Chávez-Ramírez, B. C. Dale, C. A. Haas, J. R. Herkert, F. L. Knopf, and P. D. Vickery. 2007. Conservation of grassland birds in North America: understanding ecological processes in different regions: “Report of the AOU Committee on Conservation.” Ornithological Monographs (64):1-46. https://doi.org/10.2307/40166905
Balent, K. L., and C. J. Norment. 2003. Demographic characteristics of a Grasshopper Sparrow population in a highly fragmented landscape of western New York state. Journal of Field Ornithology 74(4):341-348. https://doi.org/10.1648/0273-8570-74.4.341
Beauchamp, G., M. Bélisle, and L. A. Giraldeau. 1997. Influence of conspecific attraction on the spatial distribution of learning foragers in a patchy habitat. Journal of Animal Ecology 66(5):671-682. https://doi.org/10.2307/5920
Bédard, J., and G. LaPointe. 1984. Banding returns, arrival times, and site fidelity in the Savannah Sparrow. Wilson Bulletin 96(2):196-205.
Best, L. B. 1977. Territory quality and mating success in the Field Sparrow (Spizella pusilla). Condor 79(2):192-204. https://doi.org/10.2307/1367162
Bollinger, E. K., and T. A. Gavin. 1989. The effects of site quality on breeding-site fidelity in Bobolinks. Auk 106(4):584-594.
Bozzo, J. M. 2023. Eastern Meadowlark migration, nest success, and response to land use change in Illinois. Thesis. University of Illinois at Urbana-Champaign, Illinois, USA. https://hdl.handle.net/2142/120183
Brook, B., N. Sodhi, and C. Bradshaw. 2008. Synergies among extinction drivers under global change. Trends in Ecology & Evolution 23(8):453-460. https://doi.org/10.1016/j.tree.2008.03.011
Brooks, M. E., K. Kristensen, K. J. van Benthem, A. Magnusson, C. W. Berg, A. Nielsen, H. J. Skaug, M. Mächler, and B. M. Bolker. 2017. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R Journal 9(2):378-400. https://doi.org/10.32614/RJ-2017-066
Buehler, D. A., J. J. Giocomo, J. Jones, P. B. Hamel, C. M. Rogers, T. A. Beachy, D. W. Varble, C. P. Nicholson, K. L. Roth, J. Barg, R. J. Robertson, J. R. Robb, and K. Islam. 2008. Cerulean Warbler reproduction, survival, and models of population decline. Journal of Wildlife Management 72(3):646-653. https://doi.org/10.2193/2006-339
Calenge, C. 2006. The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling 197:516-519. https://doi.org/10.1016/j.ecolmodel.2006.03.017
Clarke, J. R., T. J. Hovick, B. Geaumont, J. P. Harmon, and K. Sedivec. 2026. Plant litter as a cornerstone of grassland ecosystem services. Agriculture, Ecosystems & Environment 399:110171. https://doi.org/10.1016/j.agee.2025.110171
Colwell, M. A., and L. W. Oring. 1989. Return rates of prairie shorebirds: sex and species differences. Wader Study Group Bulletin 55(1).
Cooper, B. S., R. D. Elmore, F. S. Guthery, and P. Van Els. 2014. Territory characteristics of Cassin’s Sparrows in northwestern Oklahoma. American Midland Naturalist 171(1):90-96. https://doi.org/10.1674/0003-0031-171.1.90
Davis, C. A., R. T. Churchwell, S. D. Fuhlendorf, D. M. Engle, and T. J. Hovick. 2016. Effect of pyric herbivory on source-sink dynamics in grassland birds. Journal of Applied Ecology 53(4):1004-1012. https://doi.org/10.1111/1365-2664.12641
Davis, S. K., and W. E. Lanyon. 2020. Western Meadowlark (Sturnella neglecta), version 1.0. In A. F. Poole, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.wesmea.01
Dechant, J. A., M. L. Sondreal, D. H. Johnson, L. D. Igl, C. M. Goldade, A. L. Zimmerman, and B. R. Euliss. 2002. Effects of management practices on grassland birds: Western Meadowlark. USGS Northern Prairie Wildlife Research Center, Jamestown, North Dakota, USA.
Diemer, K. M., and J. J. Nocera. 2014. Associations of Bobolink territory size with habitat quality. Annales Zoologici Fennici 51(6):515-525. https://doi.org/10.5735/086.051.0607
Doak, D. F., D. Bigger, E. K. Harding, M. A. Marvier, R. E. O’Malley, and D. Thomson. 1998. The statistical inevitability of stability‐diversity relationships in community ecology. American Naturalist 151(3):264-276. https://doi.org/10.1086/286117
Duchardt, C. J., J. R. Miller, D. M. Debinski, and D. M. Engle. 2016. Adapting the fire-grazing interaction to small pastures in a fragmented landscape for grassland bird conservation. Rangeland Ecology & Management 69(4):300-309. https://doi.org/10.1016/j.rama.2016.03.005
Duquette, C. A. 2020. Restoring heterogeneity-based management to rangelands for the benefit of grassland birds and floral resources. Dissertation. North Dakota State University, Fargo, North Dakota, USA.
Duquette, C. A., T. J. Hovick, B. A. Geaumont, J. P. Harmon, R. F. Limb, and K. K. Sedivec. 2022a. Embracing inherent and imposed sources of heterogeneity in rangeland bird management. Ecosphere 13(12):e4304. https://doi.org/10.1002/ecs2.4304
Duquette, C., D. A. McGranahan, M. Wanchuk, T. Hovick, R. Limb, and K. Sedivec. 2022b. Heterogeneity-based management restores diversity and alters vegetation structure without decreasing invasive grasses in working mixed-grass prairie. Land 11(8):1135. https://doi.org/10.3390/land11081135
Dyke, S. R., K. K. Johnson, and P. T. Isakson. 2015. North Dakota state wildlife action plan. North Dakota Game and Fish Department, Bismarck, North Dakota, USA.
Fajardo, N., A. M. Strong, N. G. Perlut, and N. J. Buckley. 2009. Natal and breeding dispersal of Bobolinks (Dolichonyx oryzivorus) and Savannah Sparrows (Passerculus sandwichensis) in an agricultural landscape. Auk 126(2):310-318. https://doi.org/10.1525/auk.2009.07097
Figge, F. 2004. Bio-folio: applying portfolio theory to biodiversity. Biodiversity and Conservation 13(4):827-849. https://doi.org/10.1023/B:BIOC.0000011729.93889.34
Fletcher Jr., R. J., R. R. Koford, and D. A. Seaman. 2006. Critical demographic parameters for declining songbirds breeding in restored grasslands. Journal of Wildlife Management 70(1):145-157. https://doi.org/10.2193/0022-541X(2006)70[145:CDPFDS]2.0.CO;2
Fox, J., and S. Weisberg. 2019. An R companion to applied regression. Third edition. SAGE, Thousand Oaks, California, USA.
Fuhlendorf, S. D., and D. M. Engle. 2001. Restoring heterogeneity on rangelands: ecosystem management based on evolutionary grazing patterns. BioScience 51(8):625-632. https://doi.org/10.1641/0006-3568(2001)051[0625:RHOREM]2.0.CO;2
Fuhlendorf, S. D., D. M. Engle, J. Kerby, and R. Hamilton. 2009. Pyric herbivory: rewilding landscapes through the recoupling of fire and grazing. Conservation Biology 23(3):588-598. https://doi.org/10.1111/j.1523-1739.2008.01139.x
Fuhlendorf, S. D., W. C. Harrell, D. M. Engle, R. G. Hamilton, C. A. Davis, and D. M. Leslie, Jr. 2006. Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecological Applications 16(5):1706-1716. https://doi.org/10.1890/1051-0761(2006)016[1706:SHBTBF]2.0.CO;2
Gavin, T. A., and E. K. Bollinger. 1988. Reproductive correlates of breeding‐site fidelity in Bobolinks (Dolichonyx oryzivorus). Ecology 69(1):96-103. https://doi.org/10.2307/1943164
Gill, D. E., P. Blank, J. Parks, J. B. Guerard, B. Lohr, E. Schwartzman, J. G. Gruber, G. Dodge, C. A. Rewa, and H. F. Sears. 2006. Plants and breeding bird response on a managed conservation reserve program grassland in Maryland. Wildlife Society Bulletin 34(4):944-956. https://doi.org/10.2193/0091-7648(2006)34[944:PABBRO]2.0.CO;2
Green, M. T. 1992. Adaptations of Baird’s Sparrows (Ammodramus bairdii) to grasslands: acoustic communication and nomadism. Dissertation. University of North Carolina, Chapel Hill, North Carolina, USA.
Groce, J. E., H. A. Mathewson, M. L. Morrison, and N. Wilkins. 2010. Scientific evaluation for the 5-year status review of the Golden-cheeked Warbler. Institute of Renewable Natural Resources, Department of Wildlife & Fisheries Sciences, Texas A&M University, College Station, Texas, USA. https://nri.tamu.edu/publications/research-reports/2010/scientific-evaluation-for-the-5-year-status-review-of-the-golden-cheeked-warbler/
Hartig, F. 2022. DHARMa: Residual diagnostics for hierarchical (multi-Level/mixed) regression models. https://doi.org/10.32614/CRAN.package.DHARMa
Higgins, K. F. 1984. Lightning fires in North Dakota grasslands and in pine-savanna lands of South Dakota and Montana. Journal of Range Management 37(2):100-103. https://doi.org/10.2307/3898892
Hinde, R. A. 1956. The biological significance of the territories of birds. Ibis 98(3):340-369. https://doi.org/10.1111/j.1474-919X.1956.tb01419.x
Hoover, J. P. 2003. Decision results for site fidelity in a migratory bird, the Prothonotary Warbler. Ecology 84(2):416-430. https://doi.org/10.1890/0012-9658(2003)084[0416:DRFSFI]2.0.CO;2
Hovick, T. J., J. M. Carroll, R. D. Elmore, C. A. Davis, and S. D. Fuhlendorf. 2017a. Restoring fire to grasslands is critical for migrating shorebird populations. Ecological Applications 27(6):1805-1814. https://doi.org/10.1002/eap.1567
Hovick, T. J., R. D. Elmore, S. D. Fuhlendorf, D. M. Engle, and R. G. Hamilton. 2015. Spatial heterogeneity increases diversity and stability in grassland bird communities. Ecological Applications 25(3):662-672. https://doi.org/10.1890/14-1067.1
Hovick, T. J., D. A. McGranahan, R. D. Elmore, J. R. Weir, and S. D. Fuhlendorf. 2017b. Pyric-carnivory: raptor use of prescribed fires. Ecology and Evolution 7(21):9144-9150. https://doi.org/10.1002/ece3.3401
Igl, L. D., and D. H. Johnson. 1999. Le Conte’s Sparrows breeding in conservation reserve program fields: precipitation and patterns of population change. Studies in Avian Biology 19:178-186.
Ingold, D. J., J. L. Dooley, and N. Cavender. 2010. Nest-site fidelity in grassland birds on mowed versus unmowed areas on a reclaimed surface mine. Northeastern Naturalist 17(1):125-134. https://doi.org/10.1656/045.017.0110
Jones, S. L., J. S. Dieni, M. T. Green, and P. J. Gouse. 2007. Annual return rates of breeding grassland songbirds. Wilson Journal of Ornithology 119(1):89-94. https://doi.org/10.1676/05-158.1
Kendeigh, S. C. 1941. Birds of a prairie community. Condor 43(4):165-174. https://doi.org/10.2307/1364328
Kirsch, L. M., and A. D. Kruse. 1973. Prairie fires and wildlife. Proceedings Tall Timbers Fire Ecology Conference 12:289-303.
Klimkiewicz, K. M., and A. G. Futcher. 1989. Longevity records of North American birds, Supplement 1. Journal of Field Ornithology 60(4):469-494.
Klug, P. E., S. L. Jackrel, and K. A. With. 2010. Linking snake habitat use to nest predation risk in grassland birds: the dangers of shrub cover. Oecologia 162(3):803-813. https://doi.org/10.1007/s00442-009-1549-9
Lanyon, W. E. 1956. Territory in the meadowlarks, genus Sturnella. Ibis 98(3):485-489. https://doi.org/10.1111/j.1474-919X.1956.tb01433.x
Lanyon, W. E. 1957. The comparative biology of the meadowlarks (Sturnella) in Wisconsin. Publications of the Nuttall Ornithological Club. No. 1. Cambridge, Massachusetts, USA.
Laubach, R. 1984. Breeding birds of sheeder prairie preserve, west-central Iowa. Proceedings of the Iowa Academy of Science 91(4):153-163.
Lenth, R. V. 2025. emmeans: Estimated marginal means, aka least-squares means. https://doi.org/10.32614/CRAN.package.emmeans
Mantyka‐Pringle, C. S., T. G. Martin, and J. R. Rhodes. 2013. Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta‐analysis. Global Change Biology 19(5):1642-1644. https://doi.org/10.1111/gcb.12148
Marta, S., M. Brunetti, R. Manenti, A. Provenzale, and G. F. Ficetola. 2021. Climate and land-use changes drive biodiversity turnover in arthropod assemblages over 150 years. Nature Ecology & Evolution 5(9):1291-1300. https://doi.org/10.1038/s41559-021-01513-0
McNew, L. B., V. L. Winder, J. C. Pitman, and B. K. Sandercock. 2015. Alternative rangeland management strategies and the nesting ecology of Greater Prairie-Chickens. Rangeland Ecology & Management 68(3):298-304. https://doi.org/10.1016/j.rama.2015.03.009
Murphy, M. T. 1996. Survivorship, breeding dispersal and mate fidelity in Eastern Kingbirds. Condor 98(1):82-92. https://doi.org/10.2307/1369511
Murray, B. G. 1969. A comparative study of the Le Conte’s and Sharp-Tailed Sparrows. Auk 86(2):199-231. https://doi.org/10.2307/4083496
Nocera, J. J., G. J. Forbes, and L.-A. Giraldeau. 2009. Aggregations from using inadvertent social information: a form of ideal habitat selection. Ecography 32(1):143-152. https://doi.org/10.1111/j.1600-0587.2008.05614.x
North Dakota Agricultural Weather Network (NDAWN). 2025. North Dakota agricultural weather network. https://ndawn.ndsu.nodak.edu/
Ojima, D. S., R. T. Conant, W. J. Parton, J. M. Lackett, and T. L. Even. 2021. Recent climate changes across the Great Plains and implications for natural resource management practices. Rangeland Ecology & Management 78:180-190. https://doi.org/10.1016/j.rama.2021.03.008
Ortega, Y. K., K. S. McKelvey, and D. L. Six. 2006. Invasion of an exotic forb impacts reproductive success and site fidelity of a migratory songbird. Oecologia 149(2):340-351. https://doi.org/10.1007/s00442-006-0438-8
Parmesan, C., and G. Yohe. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421(6918):37-42. https://doi.org/10.1038/nature01286
Petrie, M. D., D. P. C. Peters, J. Yao, J. M. Blair, N. D. Burruss, S. L. Collins, J. D. Derner, L. A. Gherardi, J. R. Hendrickson, O. E. Sala, P. J. Starks, and J. L. Steiner. 2018. Regional grassland productivity responses to precipitation during multiyear above- and below-average rainfall periods. Global Change Biology 24(5):1935-1951. https://doi.org/10.1111/gcb.14024
Porneluzi, P. A. 2003. Prior breeding success affects return rates of territorial male ovenbirds. Condor 105(1):73-79. https://doi.org/10.1093/condor/105.1.73
Pyle, P. 1997. Identification guide to North American birds, part I: Columbidae to Ploceidae. Slate Creek Press, Bolinas, California, USA.
R Core Team. 2025. R: a language and environment for statistical computing (version 4.4.2). R Foundation for Statistical Computing. https://doi.org/10.32614/R.manuals
Reed, M. J. 1985. A comparison of the "flush" and spot-map methods for estimating the size of Vesper Sparrow territories. Journal of Field Ornithology 56(2):5.
Ribic, C. A., R. R. Koford, J. R. Herkert, D. H. Johnson, N. D. Niemuth, D. E. Naugle, K. K. Bakker, D. W. Sample, and R. B. Renfrew. 2009. Area sensitivity in North American grassland birds: patterns and processes. Auk 126(2):233-244. https://doi.org/10.1525/auk.2009.1409
Schaeff, C., and J. Picman. 1988. Destruction of eggs by Western Meadowlarks. Condor 90(4):935-937. https://doi.org/10.2307/1368852
Schlossberg, S. 2009. Site fidelity of shrubland and forest birds. Condor 111(2):238-246. https://doi.org/10.1525/cond.2009.080087
Small, D. M., M. E. Gimpel, and D. E. Gill. 2012. Site fidelity and natal philopatry in Dickcissels. Northeastern Naturalist 19(1):123-129. https://doi.org/10.1656/045.019.0110
Stauffer, G. E. 2008. Nesting ecology and site fidelity of grassland sparrows on reclaimed surface mines in Pennsylvania. Thesis. Pennsylvania State University, Pennsylvania, USA.
Stott, P. 2016. How climate change affects extreme weather events. Science 352(6293):1517-1518. https://doi.org/10.1126/science.aaf7271
Tilman, D., C. L. Lehman, and C. E. Bristow. 1998. Diversity-stability relationships: statistical inevitability or ecological consequence? American Naturalist 151(3):277-282. https://doi.org/10.1086/286118
Verheijen, B. H. F., H. L. Clipp, A. J. Bartolo, W. E. Jensen, and B. K. Sandercock. 2019. Effects of patch-burn grazing on breeding density and territory size of Dickcissels. Avian Conservation and Ecology 14(1):7. https://doi.org/10.5751/ACE-01343-140107
Wenny, D. G. 2003. Site fidelity and return rates of Grasshopper Sparrows at three sand prairies in Northwest Illinois (Technical Report 2002 (23)). Illinois Department of Natural Resources, Springfield, Illinois, USA.
Whitaker, D. M., and I. C. Warkentin. 2010. Spatial ecology of migratory passerines on temperate and boreal forest breeding grounds. Auk 127(3):471-484. https://doi.org/10.1525/auk.2010.127.3.471
Wiens, J. A. 1970. Avian populations and patterns of habitat occupancy at the Pawnee site, 1968-1969. Technical Report No. 63. U.S. International Biological Program, Oregon State University, Corvalis, Oregon, USA. https://api.mountainscholar.org/server/api/core/bitstreams/652f5200-c0fd-4c83-94ae-35a296e5c39a/content
Wiens, J. A. 1971. Avian ecology and distribution in the comprehensive network, 1970. Grassland Biome Technical Report No. 77. Colorado State University, Fort Collins, Colorado, USA. http://hdl.handle.net/10217/16087
Wiens, J. A., J. T. Rotenberry, and B. Van Horne. 1985. Territory size variations in shrubsteppe birds. Auk 102(3):500-505. https://doi.org/10.1093/auk/102.3.500
Wright, C. K., and M. C. Wimberly. 2013. Recent land use change in the western corn belt threatens grasslands and wetlands. Proceedings of the National Academy of Sciences 110(10):4134-4139. https://doi.org/10.1073/pnas.1215404110
Zammarelli, M. B., M. P. Ayres, H. M. ter Hofstede, D. A. Lutz, and R. T. Holmes. 2024. Territory sizes and patterns of habitat use by forest birds over five decades: ideal free or ideal despotic? Ecology Letters 27(12):e14525. https://doi.org/10.1111/ele.14525
Zimmerman, J. L., and E. J. Finck. 1989. Philopatry and correlates of territorial fidelity in male Dickcissels. North American Bird Bander 14(3):5.
Fig. 1
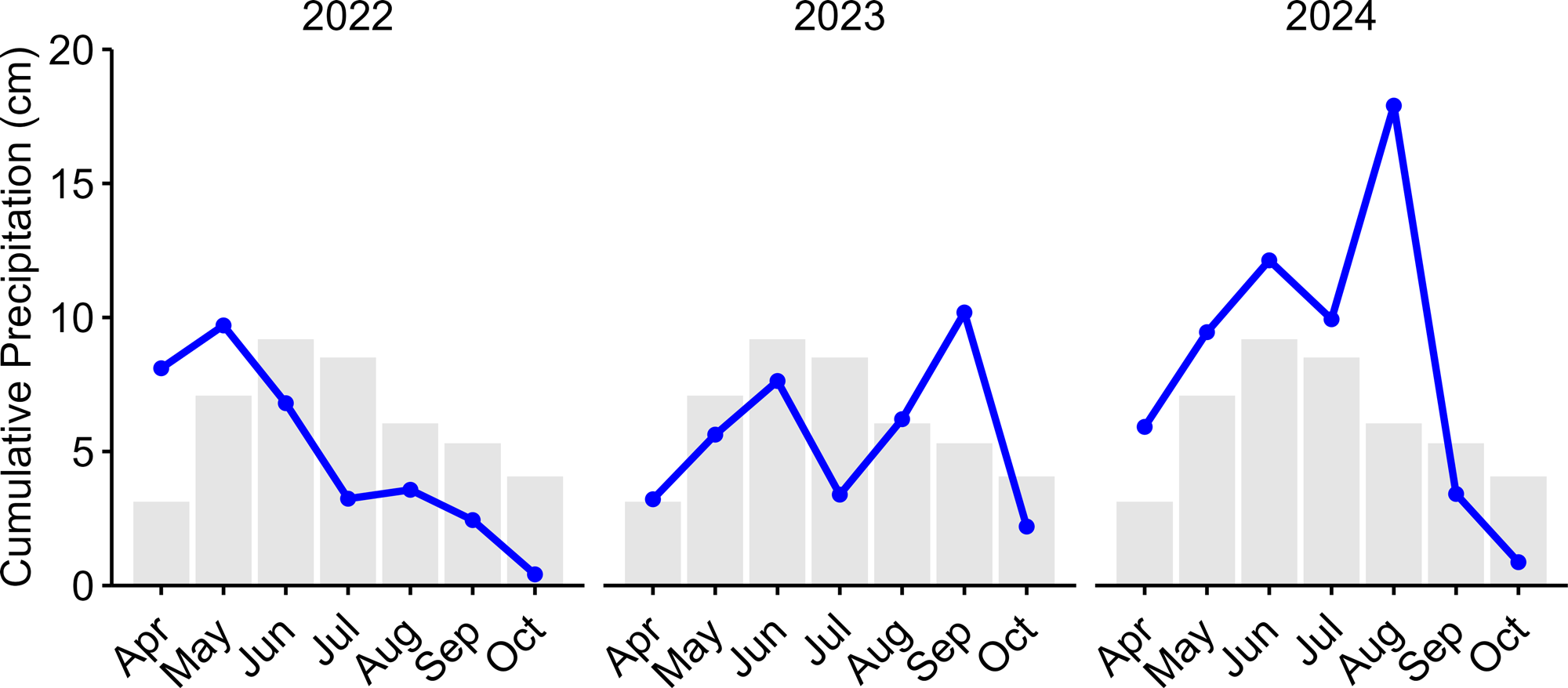
Fig. 1. Monthly precipitation from the North Dakota Agricultural Weather Network (NDAWN) station located in Streeter, North Dakota. Colored points indicate cumulative monthly precipitation for 2022 to 2024, with lines connecting monthly trends for each year. Gray bars represent the 30-year average. This data was accessed on 2 March 2025 (NDAWN 2025).
Fig. 2
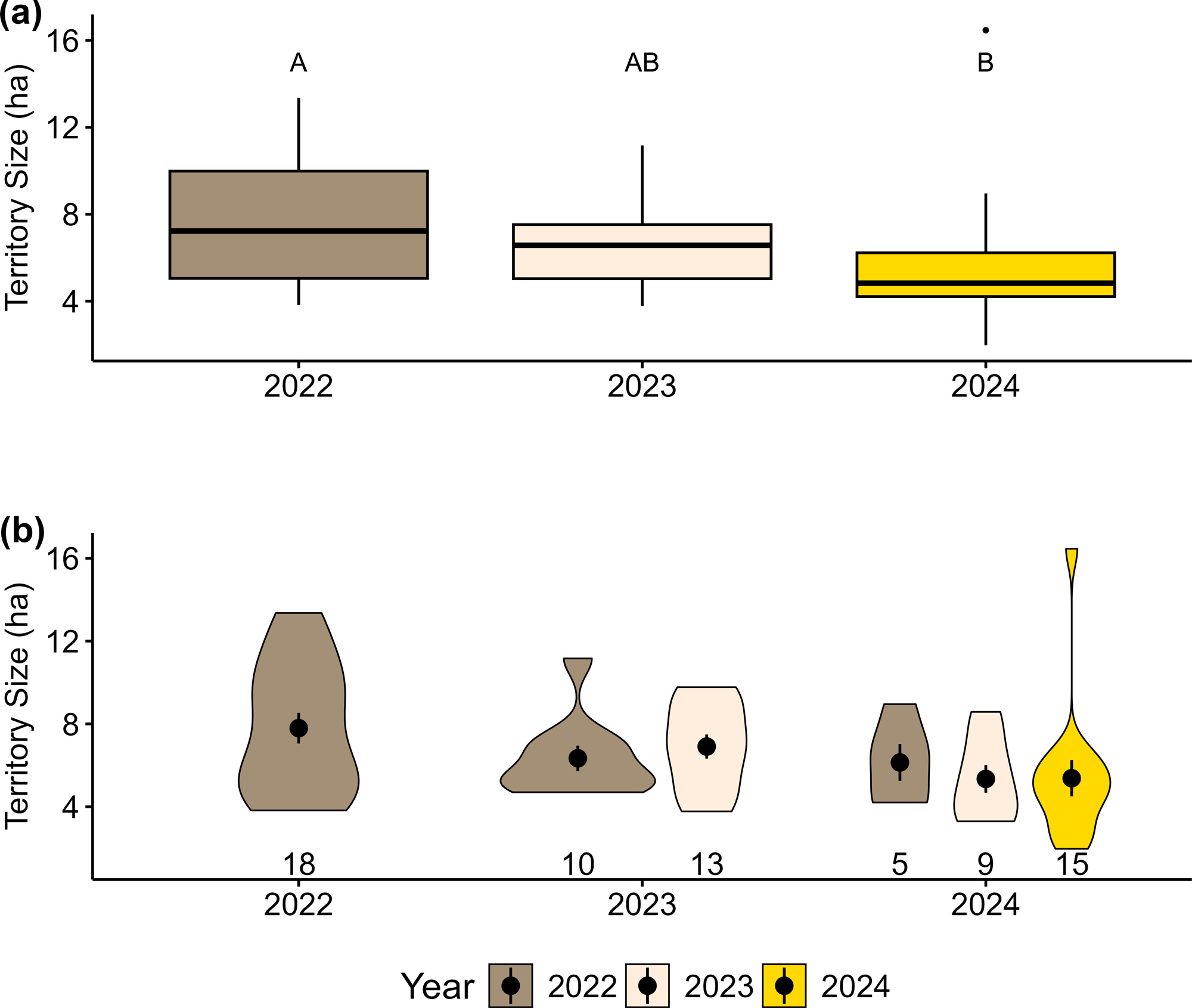
Fig. 2. Western Meadowlark (Sturnella neglecta) territory sizes (n = 70) from 2022 to 2024 at the Central Grasslands Research Extension Center near Streeter, North Dakota. Panel (a) shows a significant decline in territory size over time with territories in 2024 being smaller than those in 2022 as indicated by letters above each boxplot. Panel (b) shows the distribution of territory sizes by return status across years. Violin plots display the density and distribution of territory sizes, with plots color coded based on when each cohort was originally banded. There was no significant difference in territory size among return status categories. Points and error bars represent means ± standard errors. Sample sizes for each year/return combination are shown below each violin plot. Letters denote significant differences based on Tukey-adjusted pairwise comparisons.
Table 1
Table 1. Summary of new Western Meadowlark (Sturnella neglecta) territories, number of returning birds for each year, and total territories (new territories + returning) delineated at the Central Grasslands Research Extension Center near Streeter, North Dakota from 2022 to 2024. A total of 46 banded males established breeding territories, and 70 territories were delineated across the study period. Numbers in parenthesis indicate additional individuals that were used in return rate calculations but not territory size estimates.
| Year | New territories | Returning from 2022 |
Returning from 2023 |
Total territories | |||||
| 2022 | 18 | - | - | 18 | |||||
| 2023 | 13 (1) | 10 (1) | - | 23 (2) | |||||
| 2024 | 15 | 5 (1) | 9 | 29 (1) | |||||
| Overall | 46 | - | - | 70 (3) | |||||
Table 2
Table 2. Summary of Western Meadowlark (Sturnella neglecta) territory sizes at the Central Grasslands Research Extension Center near Streeter, North Dakota from 2022 to 2024. Territory sizes are reported according to the number of years a bird returned as follows: individuals that did not return (none), individuals that returned in 1 year (one), and individuals that returned in two years (two). The “all returns” row reports territory sizes for all birds that returned in either one or two years. Territory size is reported as the minimum, maximum, mean, and standard deviation for each year in hectares (ha).
| Years returning | Total territories |
Minimum territory size | Maximum territory size | Mean territory size | Standard deviation | ||||
| None | 46 | 1.97 | 16.46 | 6.76 | 3.09 | ||||
| One | 19 | 3.30 | 11.16 | 5.87 | 1.98 | ||||
| Two | 5 | 4.21 | 8.96 | 6.14 | 1.99 | ||||
| All returns | 24 | 3.30 | 11.16 | 5.93 | 1.94 | ||||