







The following is the established format for referencing this article:
Vaishnav, T., R. C. Shaw, and K. C. Burns. 2026. What they do in the shadows: partial nocturnality in a large forest parrot. Journal of Field Ornithology 97(2):8.ABSTRACT
Approximately 97% of all bird species are considered diurnal, although diel activity patterns of birds may be more flexible than previously believed. Much remains unknown about nocturnal activity in typically diurnal birds, which may result in potential loss of information about an important aspect of their behavior and ecology. Parrots (Order Psittaciformes), except for the truly nocturnal species, are generally not known to be active at night. However, Kākā (Nestor meridionalis) is an interesting candidate to understand how nocturnal investigations can challenge the preconceptions of avian activity patterns. Kākā are large forest parrots endemic to Aotearoa New Zealand, with historical and anecdotal evidence of remaining active after dark. We conducted the first field study to assess the activity of wild Kākā during both night and day, using ad libitum and instantaneous sampling techniques. In 122 sampling hours, we recorded 326 Kākā encounters, 17.5% of which occurred at night. In addition to resting, Kākā were observed moving through and flying above the canopy, grooming, and foraging nocturnally. The estimated percentages of observed behaviors differed between sampling techniques, as higher estimates of relatively inconspicuous behaviors, such as resting, were detected using instantaneous sampling. Kākā vocalized during day and night, with high-pitched, long-distance calls being predominant at both times of day. Nocturnal call rates were unrelated to moon phases. Our findings suggest that Kākā exhibit partial nocturnality, which is an unexplored aspect of parrot ecology with implications for conservation and habitat management. Paying greater attention to nocturnal behaviors in typically diurnal species may offer valuable insights for conservation in response to artificial light at night and noise pollution in a rapidly urbanizing environment.
RESUMEN
Aproximadamente el 97 % de todas las especies de aves se consideran diurnas, aunque los patrones de actividad diurna de las aves pueden ser más flexibles de lo que se creía anteriormente. Aún se desconoce mucho sobre la actividad nocturna en aves típicamente diurnas, lo que puede suponer una pérdida potencial de información sobre un aspecto importante de su comportamiento y ecología. Los loros (orden Psittaciformes), salvo las especies verdaderamente nocturnas, no suelen ser conocidos por estar activos por la noche. Sin embargo, el kākā (Nestor meridionalis) es un candidato interesante para comprender cómo las investigaciones nocturnas pueden cuestionar las ideas preconcebidas sobre los patrones de actividad de las aves. Los kākā son grandes loros forestales endémicos de Aotearoa (Nueva Zelanda), con pruebas históricas y anecdóticas de que permanecen activos tras el anochecer. Llevamos a cabo el primer estudio de campo para evaluar la actividad de los kākā silvestres tanto durante la noche como durante el día, utilizando técnicas de muestreo ad libitum e instantáneo. En 122 horas de muestreo, registramos 326 encuentros con kākā, el 17,5 % de los cuales se produjeron por la noche. Además de descansar, se observó a los kākā desplazándose a través del dosel y volando por encima de él, acicalándose y buscando alimento durante la noche. Los porcentajes estimados de los comportamientos observados variaron según las técnicas de muestreo, ya que se detectaron estimaciones más altas de comportamientos relativamente discretos, como el descanso, al utilizar el muestreo instantáneo. Los kākā vocalizaron tanto de día como de noche, predominando en ambos momentos del día los llamados agudos y de largo alcance. Las tasas de las llamadas nocturnas no guardaron relación con las fases lunares. Nuestros hallazgos sugieren que los kākā muestran una nocturnidad parcial, lo que constituye un aspecto inexplorado de la ecología de los loros con implicaciones para la conservación y la gestión del hábitat. Prestar mayor atención a los comportamientos nocturnos en especies típicamente diurnas puede ofrecer información valiosa para la conservación en respuesta a la luz artificial nocturna y la contaminación acústica en ambientes con rápida urbanización.
INTRODUCTION
Time is an important aspect in the fundamental niche of living organisms that has ecological, evolutionary, and physiological implications (Kronfeld-Schor and Dayan 2003, Pollard and Blumstein 2008). Animals have evolved daily activity patterns in the 24-hour periodic cycle to regulate their metabolic and energetic needs (Halle and Stenseth 2000), and in response to variation in external abiotic (e.g., temperature, light) and biotic (e.g., predation, competition) factors (Daan 1981, Cox and Gaston 2024). Diel activity patterns of animals are broadly classified as diurnal (active during day), nocturnal (active at night), crepuscular (active at twilight), and cathemeral (active both day and night; Martin 1990, Yambem and Jain 2023). Although these activity patterns are generally thought to be invariable within species, recent evidence suggests that circadian rhythms of some animals may be more flexible than previously believed (Mukhin et al. 2009, Riede et al. 2017, van der Veen et al. 2017, Alonso et al. 2021, Cox and Gaston 2024).
Nocturnality, or being active at night, is rare among birds (Martin 1990, Ward et al. 2014). In general, most birds are diurnal, with truly nocturnal birds comprising less than 3% of all avian species (Martin 1990). Birds belonging to the orders Strigiformes (owls), Caprimulgiformes (nightjars), and Apterygiformes (kiwis) are typically nocturnal (Wu 2020). Many shorebirds (Order Charadriiformes) forage at night, while two parrot species (Night Parrot Pezoporus occidentalis, and Kākāpō Strigops habroptilus, Order Psittaciformes) exhibit true nocturnality (Wood 1986, Chambers and Worthy 2013, Wu 2020). Nevertheless, some diurnal birds undertake partial or occasional nocturnal activity, chiefly involving movement, vocalization, and foraging (Martin 1990, Martin 2017). Night-time activity in diurnal birds remains under-explored, which may result in potential loss of information about their behavior and ecology.
There are several functional hypotheses for night-time activity in diurnal birds (La 2012). For instance, European Robins (Erithacus rubecula) may vocalize at night to avoid acoustic competition with day-time noises (Fuller et al. 2007). Similarly, the nocturnal calls of Common Loons (Gavia immer) travel across longer distances compared to diurnal calls, likely due to the enhanced sound transmission at night (Mennill 2014). Nocturnal activity also includes behaviors that potentially promote successful breeding, such as mate attraction in the Field Sparrows (Spizella pusilla; Celis-Murillo et al. 2016a, 2016b), pair bonding in Grasshopper Sparrows (Ammodramus savannarum; Vickery 1996), and mate guarding in the Common Nightingales (Luscinia megarhynchos; Thomas 2002). The extension of activities by typically diurnal birds into night-time may, therefore, impact their energy allocation and reproductive success (La 2012).
“Nighttime ecology” remains poorly represented in ecological research (Gaston 2019); however, interest in the nocturnal behavior of diurnal birds has increased in recent years, particularly in relation to vocalization (La 2012, Celis-Murillo et al. 2016a, 2016b, Foote et al. 2017, Dickerson et al. 2020) and local movements (Mukhin et al. 2005, 2009, Roth et al. 2009, Ward et al. 2014). Vocalization and movement are conspicuous behaviors that can be studied using remote sensing techniques, such as acoustic recorders, thermal cameras, and activity loggers (e.g., Mitchell and Clarke 2019, Alonso et al. 2021, Gillings and Scott 2021). By contrast, direct observations of wild bird behavior at night by a human observer are uncommon, likely because of the inherent difficulties of nocturnal field research, such as poor visual capability of humans, and logistical challenges of detecting birds at night (Santos et al. 2008). Consequently, much remains unknown about the night life of many diurnal birds. Direct field observation by a human observer remains a valuable method for detecting rare and inconspicuous behaviors, and behaviors that are difficult to observe using remote sensing techniques, such as foraging, grooming, and resting.
Parrots (Order Psittaciformes) exhibit behavioral flexibility to cope with both naturally occurring and anthropogenic environmental variation (Salinas-Melgoza et al. 2013). Parrot populations in modified landscapes, such as urban habitats, show rapid plastic adaptations in habitat use, including flexible movement patterns to track green spaces, and the use of non-native plant resources for nesting, roosting, and foraging (Charles and Linklater 2013, Salinas-Melgoza et al. 2013, Groom et al. 2014, Sourav et al. 2018, Geary et al. 2021). Many species employ dietary plasticity in response to variation in food availability (Renton et al. 2015). This behavioral flexibility may also extend to the diel activity patterns of parrots. Several parrot species might be more plastic in their behavior than previously assumed. For instance, the Ground Parrot (Pezoporus wallicus) was once considered to be nocturnal, but subsequent investigations of their movement and vocalization revealed that they were active throughout the day (McFarland 1991), and hence, cannot be classified as truly nocturnal. Similarly, Bourke’s Parrot (Neopsephotus bourkii) is known to be active in the crepuscular period (Fisher et al. 1972). Roosting is the primary nocturnal behavior observed in diurnal parrots (e.g., Chapman et al. 1989, Harms and Eberhard 2003, Boyes and Perrin 2009), and parrots are generally believed not to fly at night (Gilardi and Munn 1998). These studies suggest that much is unknown about the activity patterns of many parrot species. Partial nocturnality, or extending diurnal activities into the night, remains understudied in the typically-diurnal parrots.
Aotearoa New Zealand is home to the most basal clade of parrots (Family Strigopidae) consisting of three extant endemic species, Kākā (Nestor meridionalis), Kea (N. notabilis) and the nocturnal Kākāpō (Chambers and Worthy 2013). Of these, Kākā are the most widely distributed (see Higgins 1999). Kākā are large, non-migratory forest parrots that are active diurnally, with activity typically commencing half an hour before sunrise and persisting until half an hour after sunset (Beggs and Wilson 1991). The diurnal behaviors of Kākā include foraging, resting, preening, and social interactions (Moorhouse 1997). The vocal ethology of Kākā consists of at least five distinct call types associated with specific behaviors such as flight, copulation, and other types of social contact (Van Horik et al. 2007). There are anecdotal reports of Kākā remaining active after dark on Kāpiti Island and vocalizing prominently on moonlit nights (Beggs and Wilson 1991, Moorhouse 1997, Higgins 1999). Historically, Kākā have been described as being semi-nocturnal in Māori lore (Riley 2001), suggesting that this behavior is acknowledged in mātauranga (Indigenous Māori knowledge system), but the details are likely lost or fragmented post-colonization (Reilly 2011, Broughton and McBreen 2015). The extent of the nocturnal activity of Kākā, and the behaviors in which they engage after dark, have not yet been quantified.
Kākā populations are threatened throughout their distribution due to nest predation by introduced mammals, habitat loss, and degradation (Powlesland et al. 2009). Nonetheless, North Island Kākā (N. m. septentrionalis) are increasing in abundance because of recent conservation efforts such as pest management and relocation to fenced sanctuaries (Robertson et al. 2021). Kākā were reintroduced to Zealandia Te Māra A Tāne, a predator-free wildlife reserve near Wellington city (henceforth referred to as “Zealandia”) in 2002 (Miskelly et al. 2005). Since then, their population has grown and dispersed into the surrounding semi-urban and urban areas (see Charles and Linklater 2013). In Wellington city, Kākā frequently vocalize at night while flying over buildings and parks and visit residential gardens after dark (personal observation). These anecdotal observations make Kākā a suitable candidate for the study of flexible diel activity in a parrot species that is generally considered to be diurnal.
We conducted the first field study of wild Kākā with the explicit objective of understanding their nocturnal behavior. Given the historical and anecdotal evidence of night-time movements and vocalizations, we expected to observe Kākā engaging in a variety of other behaviors at night, such as foraging, flying, grooming, and resting. We aimed to quantify the diel activity of Kākā, to examine whether the observed behaviors differed between night and day, and whether vocalizations varied within and between night and day. During daytime, Kākā are often observed foraging on wood-boring invertebrates (Moorehouse 1997). There is some evidence of Kākā preying on nocturnal insects such as tree wētā (Hemideina spp.) in the daytime by extracting them from holes in trees (Gibbs 1998). Therefore, we expected the nocturnal activities of Kākā to possibly include foraging on nocturnal invertebrates such as tree wētā.
MATERIALS AND METHODS
Study site
All data were collected within Zealandia wildlife sanctuary (41°18.3′S, 174°44.8′E) located in Wellington on the southern tip of the North Island of New Zealand (Fig. 1). Situated 160–380 m above sea level, the reserve experiences a mild, temperate climate, and receives around 120 cm of rainfall annually (NIWA 2024). The vegetation in Zealandia consists of mixed native broadleaf forest, along with introduced radiata pines (Pinus radiata, Pinaceae; see Blick et al. 2008). In 1995, an 8.6 km-long perimeter predator-exclusion fence was built encircling a 225-ha section of the regenerating forest. Following fence installation, 15 species of introduced mammals were eradicated (mice Mus musculus later recolonized, see Blick et al. 2008), and thereafter, many rare and threatened bird species were translocated within the reserve (Miskelly et al. 2005). Six captive-bred Kākā were relocated to Zealandia in 2002, and since then their population has increased because of ongoing pest management, additional reintroductions, and other conservation efforts (Miskelly et al. 2005, Charles and Linklater 2013, Loepelt et al. 2016).
Field data collection
First, we identified and classified the behaviors of wild Kākā observed in Zealandia. We used ad libitum sampling to record opportunistic Kākā encounters during both day and night, and estimated the percentage of time birds were engaged in different behaviors. Ad libitum sampling is useful for its heuristic nature, as it enables researchers to identify patterns and develop systematic sampling protocols (Altmann 1974, Mann 1999). We then compared the percentages of behaviors observed during ad libitum sampling between night and day. Additionally, we generated an activity budget for a subset of individuals at night and in the daytime using instantaneous sampling, which provides an estimate of the percentage of time spent while engaged in various activities by recording the behavioral state of the individual at pre-selected moments in time (e.g., every minute on the minute for 30 minutes; Altmann 1974). Finally, we quantified the vocalization rates of the five most common call types of Kākā and compared them between night and day. We also evaluated the effect of moon phase on nocturnal vocalizations. If Kākā called more often on moonlit nights as previously noted (Higgins 1999), we expected call rates to be higher on nights with a full moon.
Data were collected over nine months, between February 2024 and October 2024 at night and in the daytime. We conducted the sampling within Zealandia and did not include urban study sites, such as residential areas, to avoid the potential effects of factors such as artificial light at night that may influence the behavior of urban birds. Before commencing the study, we identified various behavioral “states” defined by Altmann (1974) as ongoing behaviors of appreciable length, which were subsequently classified into six broad behavioral categories (Table 1). Behaviors related to foraging, such as feeding on wood-boring invertebrates, fruits, flowers, sap, ground-foraged seeds, and pinecones, eating pellets from the supplementary feeders, and drinking water, were grouped in a single category, foraging. Flying above canopy and moving through canopy were treated as separate categories. Self-preening, head-scratching, and wing-stretching were considered as grooming behaviors, while behaviors such as perching and sleeping were categorized as resting behaviors. Antagonistic and affiliative social interactions were grouped together in the social interactions category. We identified different call types by ear, and classified the five most common calls as whistle, screech, long, croak, and warble calls (Fig. 2).
Van Horik et al. (2007) previously identified five distinct call types of Kākā on Kāpiti Islands and termed them as shraak, shraak-woo, gurgle, bark, and snicker. However, vocalizations can sometimes differ between populations and groups of parrots, leading to local and regional dialects (Wright 1996, Bradbury et al. 2001, Baker 2011, Wright and Dahlin 2018, Aplin 2019). Geographical variation in Kākā calls has previously been observed but not studied (Higgins 1999). Therefore, to avoid any inconsistencies in call types caused by potential differences in calls between the Kāpiti Island and Zealandia populations of Kākā, we named the call types independently of previously published studies. The whistle, screech, and warble, correspond closely to the shraak-woo, bark, and snicker calls from the Kāpiti Island study, respectively (Van Horik et al. 2007). The croak and long calls from the current study are seemingly analogous to the gurgle and shraak calls on Kāpiti Island, respectively, but are structurally distinct from the sonograms presented by Van Horik et al. (2007; Fig. 2). However, there are similarities in the descriptions of these call types. The gurgle call was given by solitary birds while perched, and the shraak call was given almost exclusively by birds in flight on Kāpiti Island (Van Horik et al (2007). Correspondingly, in the current study, the croak was a softer call given mainly by perched birds, and the long call was a loud, long-duration call given by birds in flight.
Given the changing day length throughout the study period, field data was collected at least 60 minutes after sunrise for the day and 60 minutes after sunset for night by a sole observer. Daytime observations were initiated an average of 209 ± 57 minutes after sunrise (ranging from 127 and 331 minutes after sunrise), while night-time observations commenced an average of 159 ± 58 minutes after sunset (ranging from 60 to 256 minutes after sunset). For observations at night, we used the Nitecore MH25 v2 LED Flashlight with a red filter to minimize disturbance to the birds (Finley 1959). Once a bird was located using the flashlight, it was turned off and we switched to using digital night vision binoculars to continue the observations with minimal disturbance to the bird. Because Zealandia is a popular daytime tourist destination that also runs night-time tours, Kākā are likely accustomed to human presence. As a result, the effect of observer presence on bird behavior was considered negligible. Many Kākā in Zealandia are not color banded for individual identification. Therefore, sexual, and age-related differences in the diel activity of Kākā were not evaluated in this study.
Only the discernible activities of birds within a 10 m visual range of the observer were noted, except for birds flying over the canopy, which often flew over 50 m. At night, it was not possible to visually observe birds flying over the canopy, hence we used auditory cues, i.e., the vocalizations made during flight, to infer the activity of flying above the canopy. In instances where the bird moved and became visually obscured from the observer, no behavioral observations were recorded. If multiple birds were observed at the same time, the activity of a focal individual, i.e., the bird that was located closest to the observer at the onset of the observation, was documented. For each sampling session, we noted environmental variables such as cloud cover and wind conditions. For sampling sessions conducted at night, we also noted the moon phase. Sampling was terminated in unfavorable weather conditions, such as extreme wind and rain.
Behavioral estimates
We used ad libitum sampling to record encounters of Kākā and document their behavior during both day and night. In total, we conducted field observations for approximately 122 sampling hours, with 61 sampling hours during each time of day (day and night), spread over 28 daytime sampling sessions and 32 night-time sampling sessions. During each sampling session, the observer walked along the observation trail (Fig. 1), documenting opportunistic encounters of Kākā, and the behavior in which they were engaged. Once a bird was visually located, the observer stood still for 60 seconds to allow the bird to acclimatize to their presence. The observer then recorded a snapshot of the behavioral state in which the bird was engaged at that moment (e.g., foraging, perching, moving through the canopy). If the bird stayed within visual range until 60 seconds after noting the initial behavioral state, we proceeded to record the activity budget for that individual using instantaneous sampling as described below. If the bird moved out of visual range within 60 seconds after noting the behavioral state, the observation was ended. After each observation, the observer walked at least 50 m down the trail before locating birds again to avoid sampling the same bird more than once. The time spent recording instantaneous sampling data was deducted from the overall sampling time for the ad libitum sampling.
Ad libitum sampling enabled the detection and behavioral observations of Kākā during opportunistic encounters, but it may not reflect the precise activity budget of Kākā (Altmann 1974, Mann 1999). Therefore, we used instantaneous sampling to generate activity budgets of wild Kākā for day and night separately. In total, we conducted instantaneous sampling for a total of 20 individual Kākā in Zealandia. Of these, 12 birds were sampled at night for a total of 224 minutes (observation length per bird ranging from nine to 30 minutes), while eight birds were sampled during daytime for a total of 240 minutes (30 minutes per bird).
At the end of the ad libitum sampling observation, if the bird stayed within visual range until 60 seconds after noting the initial behavioral state, we proceeded to record the activity budget for that individual using instantaneous sampling. A snapshot of the behavioral state of the focal bird (e.g., foraging, perching, moving through the canopy) was noted at 60-second intervals for up to 30 minutes. The observation was completed either after 30 minutes or when the focal bird moved out of view. For night-time observations, once the focal bird was located with the help of the flashlight, the observer turned it off and switched to using night vision binoculars to make video recordings of the birds with minimum disturbance to the bird. The videos were reviewed later, and the observed behavior was noted at 60-second intervals for up to 30 minutes. At night, the observer often lost track of the bird when it went out of sight, making it difficult to sample them for the entire 30 minutes. This resulted in variable observation lengths of focal birds at night. Furthermore, the behavioral category of flying above canopy was excluded for this sampling technique because the observer followed focal individuals within an observable distance of 10 m from the observer.
Vocalization rates
Given that multiple individuals often called simultaneously, repeatedly, and while flying, it was difficult to determine the precise number of calls per minute. Therefore, we use the term “vocalization rate” to indicate the percentage of the sampling duration in which calls were detected, rather than an estimate of number of calls per minute. During each session of ad libitum sampling outlined above, the observer listened for calls given by Kākā within a 50 m radius. When a call was heard, the observer noted the time and call type (Table 1). If the same call type was heard within the same “sampling minute,” i.e., within 60 seconds of the previous call, it was not documented again for that sampling minute. However, if a different call type was heard within the same sampling minute, the observer noted it. Calls of the same type were noted again only if they occurred in the subsequent sampling minute. We then estimated the mean percentage of sampling time in which each call was detected over the total sampling period for day and night separately.
Statistical analysis
We first calculated the percentage of Kākā encounters that occurred at night from the total number of encounters for the entire sampling period. Then, to assess whether the behavioral estimates of Kākā differed between night and day, we estimated the percentage of time that birds were observed engaging in each behavior during day and night separately, using the data obtained from ad libitum sampling. Given that we had multinomial data with six behavioral categories, we calculated simultaneous confidence intervals (Goodman 1964). Simultaneous confidence intervals help control the family-wise error rate across multiple comparisons, reducing the risk of Type-I errors that may arise from calculating individual confidence intervals for each category (Goodman 1964, Sison and Glaz 1995, Glaz and Sison 1999). We then used mixed-effects logistic regressions to compare the percentage of each behavior between night and day, treating time of day (night vs. day) as a fixed effect and sampling date as the random effect to account for the non-independence of observations within the same sampling session. For the sake of simplicity, cloud cover and wind were not included in the model. All analyses were performed in the R environment using the packages MultinomialCI and lme4 (Bates et al. 2015, R Core Team 2022).
Estimates of observed behaviors may vary depending on the sampling method (Harrison et al. 2009, Amato et al. 2013, Munita et al. 2016). Therefore, we used the activity budgets obtained from instantaneous sampling to estimate the percentage of time that birds were observed engaging in each behavior and examine whether they varied across night and day. Given that we had multinomial data with six behavioral categories, we calculated simultaneous confidence intervals (Goodman 1964). To account for the non-independence of observations from the same focal bird, we used mixed-effects logistic regressions to compare the percentage of each behavior between night and day, treating time of day (night vs. day) as a fixed effect and individual focal bird as the random effect. Kākā were not observed engaging in social interactions at night; we excluded that category from the mixed-effects logistic regression analyses because there was a complete separation in the data between night and day (i.e., zero observations of social interactions at night resulting in a large standard error). All analyses were performed in the R environment using the packages MultinomialCI and lme4 (Bates et al. 2015, R Core Team 2022). Because of the inherent differences in data collection between ad libitum and instantaneous sampling techniques (Altmann 1974, Mann 1999), we made descriptive rather than empirical comparisons between the behavioral estimates obtained from the two methods.
Finally, to address whether vocalizations differed between night and day, we calculated the percentage of the sampling duration in which calls were detected, averaged over all the sampling sessions (defined here as the vocalization rate). This was calculated for each call type separately for day and night sampling sessions. Given the low sample size for some call types at night, we compared these calls descriptively rather than empirically. Additionally, we assessed whether Kākā called more often on nights with a certain moon phase. We calculated the total vocalization rate of all call types and compared them between the four moon phases (New moon, First quarter, Full moon, Third quarter) using the Kruskal-Wallis test. All analyses were performed in the R environment using the coin package (Hothorn et al. 2008, R Core Team 2022).
RESULTS
Diurnal and nocturnal encounters
We encountered Kākā a total of 326 times during the study period, of which approximately 17.5% encounters (n = 57) occurred at night, and 82.5% encounters occurred during the day (n = 269). Most encounters were of solitary birds, with the average group size of 1.05 ± 0.23 birds at night, and 1.27 ± 0.76 birds during the day. Behaviors from all six categories, namely flying above canopy, foraging, grooming, moving through canopy, resting, and social interaction (Table 1, Fig. 3), were detected during both day and night observations between the two sampling techniques.
How does observed behavior differ between night and day?
During ad libitum sampling in the daytime, the majority of Kākā encounters occurred while they were foraging (42.4%), followed by flying above canopy (22.3%), moving through the canopy (16.4%), grooming (7.1%), resting (5.9%), and engaging in social interactions (5.9%; Fig. 4). At night, the majority of Kākā encounters occurred while they moved through the canopy (42.1%) and flew above the canopy (40.4%), whereas foraging (6.9%), grooming (6.9%), and resting (3.5%) were detected relatively rarely (Fig. 4). Social interactions were not detected for Kākā at night using the ad libitum sampling technique.
The time of day (night vs. day) significantly influenced the probability of observing Kākā engaging in certain behaviors during ad libitum sampling. Kākā were more likely to be encountered while flying above the canopy at night compared to the day (β = 0.962, p = 0.0073). Kākā were also more likely to be encountered while moving through the canopy night than in the daytime (β = 1.333, p < 0.001). In contrast, Kākā were more likely to be encountered while foraging during the day than at night (β = -2.316, p < 0.001). There was no significant difference in the detection of grooming (β = -0.007, p = 0.99) or resting (β = -0.553, p = 0.469) between night and day (Appendix Table A1).
For the instantaneous sampling in the daytime, focal birds (N = 8) were observed resting for majority of the pooled observation time (52.5%), followed by grooming (19.6%), foraging (13.7%), moving through the canopy (7.5%), and engaging in social interactions (6.7%; Fig. 5). At night, focal Kākā (N = 12) across pooled observations were mostly observed moving through the canopy (38.6%), and resting (33.8%), followed by foraging (13.6%), and grooming (13.1%). Social interactions among Kākā were rarely detected at night (0.9%) (Fig. 5).
At the individual level, the time of day (night vs. day) influenced the probability of observing Kākā engaging in certain behaviors during instantaneous sampling. Kākā were more likely to be observed moving through the canopy at night than during daytime (β = 2.808, p < 0.001), whereas there was no significant difference in the probability of observing Kākā engaged in foraging (β = -0.038, p = 0.980), resting (β = -1.576, p = 0.119), grooming (β = -0.420, p = 0.351) and social interactions (β = -2.053, p = 0.075) between night and day (Appendix Table S2). There was high variability between individual birds engaging in each behavioral category, likely resulting in a non-significant difference in the probability of resting between night and day despite the seemingly variable estimates obtained from the simultaneous confidence intervals (Fig. 5).
Do vocalizations vary within and between night and day?
All five call types, namely whistle, screech, long, croak, and warble, were detected during the day, while only whistle, screech, long, and croak calls were also detected at night. In the daytime, the whistle call was uttered most frequently (42.9% of the sampling period), followed by screech (27.0%), long (7.5%), croak (5.28%), and warble (0.9%; Fig. 6). Nocturnal vocalizations followed a similar pattern, except that warble calls were not detected during night-time observations. At night, the whistle call was uttered most frequently (8.6%), followed by screech (5.8%), long (1.7%), and croak (0.1%; Fig. 6). Because certain call types were rarely uttered, and different call types often occurred in the same sampling minute, the total percentages do not add up to 100%. Vocalization rates differed between call types during the day. Overall, vocalization rates were significantly higher in the daytime compared to the night for all call types. Moreover, the total vocalization rate did not vary significantly between nights with different moon phases (Χ² = 0.373, df = 3, p = 0.946, Appendix Fig. A1).
DISCUSSION
Kākā engaged in a variety of behaviors at night in addition to resting, such as flying above the canopy, foraging, grooming, and moving through the canopy by hopping, swinging, walking, and flying short distances. Nearly one-fifth of all Kākā encounters occurred at night, when Kākā were chiefly encountered while they moved through the forest, either moving through or flying above the canopy. Contrary to our expectations of observing Kākā foraging on nocturnal invertebrates such as tree wētā, we did not observe Kākā foraging on plant resources or invertebrates at night. Instead, nocturnal foraging observations consisted of consuming pellets from supplementary feeders and drinking water from a puddle. Foraging was the most observed activity during daytime. Vocalizations were detected during the day and at night, with Kākā producing all but one of the call types from their vocal repertoire at night. Furthermore, while we encountered the truly nocturnal birds like the Little Spotted Kiwi (Apteryx owenii) and Ruru (Ninox novaeseelandiae) during our nighttime observations, no other diurnal bird species that are present and common in Zealandia were visually detected or heard calling at night.
Vocalizations may play a vital role in creating and maintaining social bonds among Kākā, although social groups and possible fission-fusion dynamics are not well understood for this species (Van Horik et al. 2007, Silk et al. 2014). Kākā vocalized prominently at night. The whistle and screech calls were loud, high-pitched calls that likely functioned as long-distance contact calls. The long call was given when Kākā flew above the canopy. In the field, the prolonged duration of the long call often provided the observer with information about the bird’s direction of flight. Correspondingly, the long call may serve a similar purpose for conspecifics, to indicate direction of flight and aid in flock formation, possibly assisting in fission-fusion-like social dynamics (Penndorf et al. 2025). The croak and warble calls were low-pitched calls given while perched, facilitating short-distance communication, with the warble call likely associated with courtship behavior (Van Horik et al. 2007). The absence of warble calls at night, along with the low sightings of nocturnal social interactions, suggests that courtship may be restricted to daylight hours. However, because our study was conducted primarily during the non-breeding season, it remains unclear whether this pattern persists during the breeding season.
Nocturnal calls were detected at similar rates across all four moon phases, suggesting that Kākā vocalized similarly during full moon and new moon nights, as well as during the intermediate partial moon phases. This was contrary to the previous anecdotal evidence that Kākā vocalized mainly on moonlit nights (see Higgins 1999), although we did not specifically measure the light intensity at night. Cloud cover was also not included in the analysis, which could alter moonlight intensity, and consequently affect the activity of birds. Lunar phase has varying effects on nocturnal calls of birds. For instance, Eastern Ground Parrots (P. wallicus wallicus) vocalized more often during full moon periods (Prior et al. 2023). Among passerines, Ovenbirds (Seiurus aurocapilla) and White-throated Sparrows (Zonotrichia albicollis) sing less often on nights with a full moon (Foote et al. 2017), while nocturnal singing in the Willie Wagtail (Rhipidura leucophrys) increases with lunar illumination (Dickerson et al. 2020).
Parrots are generally considered to be diurnal birds that do not fly at night; however, flexible activity patterns may be more common than often assumed (Gilardi and Munn 1998, Boyes and Perrin 2009, Renton et al. 2015). Species such as the Night Parrot and Kākāpō exhibit specialized anatomical adaptations to facilitate their nocturnal lifestyle (Corfield et al. 2011, Iwaniuk et al. 2020, Shute et al. 2023). Nevertheless, some Kākāpō individuals, particularly females with dependent offspring, may forage during the daytime (Higgins 1999). Bourke’s Parrots are typically crepuscular and use watering holes at dawn and dusk (Fisher et al. 1972). The nocturnal activity observed in Kākā may also indicate adaptations for enhanced night vision (see Corfield et al. 2011, Lind et al. 2012). Similarly, the assumption of strict diurnal activity in the New Zealand Kea warrants investigation, given anecdotal evidence of partial nocturnality, and the shared ancestry of Kea with the Kākāpō and Kākā (King and Kemp 2021). This behavioral flexibility raises important questions about the patterns of rest and sleep in Kākā.
Diurnal birds typically roost and sleep at night (Martin 1990). Sleep in animals serves several functions, including physical and neurological recovery, energy conservation, and risk reduction (Amlaner and Ball 1983). Avian sleep may also play a role in maintaining and developing vocal communication (Gaviraghi Mussoi et al. 2022). In our study, we encountered Kākā less frequently at night than in the daytime, leading to the reasonable assumption that most Kākā sleep at night. Many parrot species roost communally at night to maintain social cohesion, increase proximity to food or water resources, or reduce predation risk (Weatherhead 1983, Beauchamp 1999). Communally roosting species exhibit varying degrees of fidelity to roost locations (Harms and Eberhard 2003, Boyes and Perrin 2009). We did not observe large nocturnal congregations of Kākā or identify specific trees or locations with frequent nocturnal activity. This suggests that Kākā are mostly solitary or occur in small groups at night, which may reflect an even distribution of resources or low predation pressure compared with species in which communal roosting provides greater benefits. Instantaneous sampling resulted in relatively high resting observations during the day and at night. Thus, it is possible that the individuals that are active nocturnally may be sleeping during the day to mitigate the cognitive and physiological impairment associated with sleep deprivation (Aulsebrook et al. 2021, Johnsson et al. 2022). Further investigation of sleep in Kākā would provide deeper insight into individual variation in activity levels between night and day.
The purpose of nocturnal activity in diurnal birds is poorly understood (see La 2012). There may be several non-mutually exclusive explanations for partial nocturnality in Kākā. The evolution of nocturnality is often associated with reduced predation risk (Kahlert et al. 1996, Rahlfs and Fichtel 2010, Monterroso et al. 2013, de Matos Dias et al. 2018). Historically, Kākā may have been hunted by large avian predators in New Zealand, such as the extinct Eyles’s Harrier (Circus eylesi; Holdaway 1999). Predation pressure could have led to the extension of certain activities into the night. Some birds are known to adopt vastly different strategies against perceived threats depending on time of day; for example, the Indian Peafowl (Pavo cristatus) exhibits different call intensities and body postures in response to a similar threat between day and night (Yorzinski and Platt 2012). Alternatively, partial nocturnality may be driven by ecological factors, such as optimizing reproductive success, dispersal, and social learning (La 2012). For instance, juvenile Eurasian Reed Warblers (Acrocephalus scirpaceus), a migratory passerine species, engage in nocturnal exploratory flights outside of the migration season to aid in post-fledging dispersal (Mukhin 2004, Mukhin et al. 2005, 2009). In Nightingales (Luscinia megarhynchos), nocturnal activity is linked to sex-specific mate searching and territory prospecting (Roth et al. 2009), while Yellow-breasted Stonechats (Icteria virens) engage in extra-territorial forays and mate-guarding at night (Ward et al. 2014). Similarly, nocturnal activity in Kākā may indicate birds exploring their habitat, learning or practicing vocalizations, and developing social bonds. Seasonal and demographic variations in nocturnal activity should be investigated using tagged individuals with known life history traits and studied over multiple annual cycles to understand ontogenetic variation in nocturnal activity.
Anthropogenic disturbances that alter the nocturnal environment, such as artificial lights and noise, can severely impact avian behavioral patterns (Kronfeld-Schor et al. 2013, Nordt and Klenke 2013, Aulsebrook et al. 2020). These factors are linked with the early onset of dawn chorus in diurnal passerines like the European Blackbird (Turdus merula; Nordt and Klenke 2013), and Saffron Finch (Sicalis flaveola; Marín Gómez 2022). The song structure of male Vermillion Flycatchers (Pyrocephalus rubinus) is altered in the presence of bright lights at night (Nakamura-Garcia and Ríos-Chelén 2022), while nocturnal vocalizations of the diurnal Willie Wagtails decrease in areas with more sky glow and streetlights (Dickerson et al. 2022). Similar changes may impact Kākā populations in Wellington city as they disperse into urban areas (Charles and Linklater 2013). A comparative investigation of nocturnal behavior of Kākā over an urbanization gradient would help in determining how anthropogenic disturbances influence their activities after dark.
Human-induced disturbances, such as habitat modification, urbanization, and population declines, erode the cultural and behavioral diversity in non-human animals (van Schaik 2002, Laiolo and Tella 2007, Gruber et al. 2019, Roncero et al. 2023, Bolcato and Aplin 2025). According to Māori lore, Kākā flocks comprising hundreds of birds were once known to congregate on seasonally flowering rātā trees (Metrosideros spp.; Riley 2001). In recent times, Kākā numbers have declined drastically across their historical range (Johnson et al. 2025), which may have resulted in the loss of behavioral diversity, including their vocal repertoire, movements, and social behavior, like fission-fusion flocking dynamics (van Schaik 2002, Silk et al. 2014, Aplin 2019, Kühl et al. 2019). Although our findings align with mātauranga, which states that Kākā are “semi-nocturnal” (Riley 2001), their present-day nocturnal behavior may differ considerably from the time when their populations flourished, largely because of human-induced habitat loss and urbanization.
Night-time field observations pose many practical challenges because of the difficulty of studying organisms at night (Gaston 2019). Consequently, nocturnal activity of Kākā may be underestimated in this study compared to day-time activity because of the limited visual range and the variable conspicuousness of certain behaviors for a human observer (Morelli et al. 2022). Additionally, individuals engaged in certain inconspicuous behaviors at night, such as resting and grooming, may have gone undetected. For instance, we did not encounter Kākā sleeping at night using ad libitum sampling, possibly because of the inconspicuousness of the behavior, and the limited visual field afforded by the flashlight. Similarly, Kākā flying over the canopy at night were only detected using auditory cues, hence there would be potential loss of data if the birds did not call while in flight. Monitoring individuals using a combination of human observation and technology, such as GPS tracking devices on backpack harnesses as demonstrated by Recio et al. (2016) and Forrest et al. (2022), radio transmitters, acoustic recorders, and trail cameras (Ellis and Marsland 2022) could optimize the detection and tracking of birds over long distances and extended time frames.
An additional challenge in this study was implementing behavioral sampling methods in field conditions. We used ad libitum sampling for the flexibility it provided in locating birds and documenting their behaviors at night (Altmann 1974). Ad libitum sampling yielded higher estimates of conspicuous behaviors, such as foraging during the day, and moving through or flying above the canopy at night. In contrast, instantaneous sampling yielded higher estimates of resting at night and lower estimates of daytime foraging. Activity budgets ascertained from instantaneous sampling may more accurately reflect the true activity patterns (Mann 1999, Pöysä 1991). However, the precision of activity budget estimates is influenced by both the number of individuals sampled from a population and the number of observations recorded per individual (Morrison 1984, Pöysä 1991, Hepworth and Hamilton 2001, Wilson et al. 2008). In our study, individual identification was not possible, preventing repeated observations per focal bird. As a result, although we provide a baseline estimate of the activity budget of wild Kākā using instantaneous sampling, our findings may not represent accurate estimates because of the small sample size. Still, we considered it valuable to report these results to illustrate how different sampling methods influence behavioral estimates (Altmann 1974, Rose 2000, Harrison et al. 2009, Amato et al. 2013, Karniski et al. 2015, Munita et al. 2016, Tesar et al. 2023). Future studies on the nocturnal behavior of birds would benefit from higher sample sizes and extended sampling effort (Pöysä 1991, Wilson et al. 2008).
CONCLUSION
Diel activity patterns of parrot species may reflect behavioral flexibility similar to that observed in their diets and habitat use (McFarland 1991, Renton et al. 2015). Kākā exhibited a range of behaviors at night in addition to resting, including nocturnal movements, vocalizations, and, to a lesser extent, foraging and grooming. If the function of nocturnal activity in Kākā is similar to those in other bird species that are partially nocturnal, these behaviors may facilitate long-distance communication, mate prospecting, and the maintenance of social bonds. For juvenile birds, nocturnal activity may promote post-fledging dispersal and vocal learning. Our findings provide baseline information regarding the nocturnal activities of Kākā, which may offer valuable insights into their habitat use, social dynamics, and reproductive success and inform future conservation strategies. The purpose of partial nocturnality in Kākā warrants further exploration to enhance our understanding of flexible diel activity patterns, a phenomenon rarely reported in parrots.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Tirth Vaishnav, Rachael Shaw, and K.C. Burns conceptualized the study. Tirth Vaishnav collected and analyzed the data, and drafted the manuscript. All authors reviewed and edited the manuscript.
ACKNOWLEDGMENTS
The authors would like to thank the staff of Zealandia Te Māra A Tāne ecosanctuary, particularly Jo Ledington and Ellen Irwin, for their help with the research permit and logistical support for this study. Thanks to Lisa Woods for providing statistical advice, and to Mark Hauber and the anonymous reviewers for their insights that helped improve the manuscript. Thanks also to Riccardo Ciarle and the field volunteers for helping with the nocturnal outings. The authors acknowledge the support of the Tararua Tramping Club in funding the field expenses for this study.
DATA AVAILABILITY
Dataset is available online in the Figshare data repository: https://doi.org/10.6084/m9.figshare.31438240.
LITERATURE CITED
Alonso, J. C., I. Abril-Colón, and C. Palacín. 2021. Moonlight triggers nocturnal display in a diurnal bird. Animal Behavior 171:87-98. https://doi.org/10.1016/j.anbehav.2020.11.011
Altmann, J. 1974. Observational study of behavior: sampling methods. Behavior 49(3-4):227-266. https://doi.org/10.1163/156853974X00534
Amato, K. R., S. Van Belle, and B. Wilkinson. 2013. A comparison of scan and focal sampling for the description of wild primate activity, diet and intragroup spatial relationships. Folia Primatologica 84(2):87-101. https://doi.org/10.1159/000348305
Amlaner Jr., C. J., and N. J. Ball. 1983. A synthesis of sleep in wild birds. Behavior 87(1-2):85-119. https://doi.org/10.1163/156853983X00138
Aplin, L. M. 2019. Culture and cultural evolution in birds: a review of the evidence. Animal Behaviour 147:179-187. https://doi.org/10.1016/j.anbehav.2018.05.001
Aulsebrook, A. E., R. D. Johnsson, and J. A. Lesku. 2021. Light, sleep and performance in diurnal birds. Clocks and Sleep 3(1):115-131. https://doi.org/10.3390/clockssleep3010008
Aulsebrook, A. E., J. A. Lesku, R. A. Mulder, W. Goymann, A. L. Vyssotski, and T. M. Jones. 2020. Streetlights disrupt night-time sleep in urban black swans. Frontiers in Ecology and Evolution 8:131. https://doi.org/10.3389/fevo.2020.00131
Baker, M. C. 2011. Geographic variation of three vocal signals in the Australian ringneck (Aves: Psittaciformes): do functionally similar signals have similar spatial distributions? Behavior 148(3):373-402. https://doi.org/10.1163/000579511X559751
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67:1-48. https://doi.org/10.18637/jss.v067.i01
Beauchamp, G. 1999. The evolution of communal roosting in birds: origin and secondary losses. Behavioral Ecology 10(6):675-687. https://doi.org/10.1093/beheco/10.6.675
Beggs, J. R., and P. R. Wilson. 1991. The kaka Nestor meridionalis, a New Zealand parrot endangered by introduced wasps and mammals. Biological Conservation 56(1):23-38. https://doi.org/10.1016/0006-3207(91)90086-O
Blick, R., R. Bartholomew, T. Burrell, and K. C. Burns. 2008. Successional dynamics after pest eradication in the Karori Wildlife Sanctuary. New Zealand Natural Sciences 33:3-14.
Bolcato, S., and L. Aplin. 2025. The effect of habitat health and environmental change on cultural diversity and richness in animals. Philosophical Transactions of the Royal Society B: Biological Sciences 380(1925):20240141. https://doi.org/10.1098/rstb.2024.0141
Boyes, R. S., and M. R. Perrin. 2009. Flocking dynamics and roosting behavior of Meyer’s parrot (Poicephalus meyeri) in the Okavango Delta, Botswana. African Zoology 44(2):181-193. https://doi.org/10.3377/004.044.0206
Bradbury, J. W., K. A. Cortopassi, J. R. Clemmons, and D. Kroodsma. 2001. Geographical variation in the contact calls of orange-fronted parakeets. Auk 118(4):958-972. https://doi.org/10.1093/auk/118.4.958
Broughton, D., and K. McBreen. 2015. Mātauranga Māori, tino rangatiratanga and the future of New Zealand science. Journal of the Royal Society of New Zealand 45(2):83-88. https://doi.org/10.1080/03036758.2015.1011171
Celis-Murillo, A., T. J. Benson, J. R. Sosa-López, and M. P. Ward. 2016a. Nocturnal songs in a diurnal passerine: attracting mates or repelling intruders? Animal Behavior 118:105-114. https://doi.org/10.1016/j.anbehav.2016.04.023
Celis-Murillo, A., K. W. Stodola, B. Pappadopoli, J. M. Burton, and M. P. Ward. 2016b. Seasonal and daily patterns of nocturnal singing in the Field Sparrow (Spizella pusilla). Journal of Ornithology 157:853-860. https://doi.org/10.1007/s10336-015-1318-y
Chambers, G. K., and T. H. Worthy. 2013. Our evolving view of the kakapo (Strigops habroptilus) and its allies. Notornis 60(3):197-200. https://doi.org/10.63172//277926uikyof
Chapman, C. A., L. J. Chapman, and L. Lefebvre. 1989. Variability in parrot flock size: possible functions of communal roosts. Condor 91:842-847. https://doi.org/10.2307/1368068
Charles, K. E., and W. L. Linklater. 2013. Behavior and characteristics of sap-feeding North Island kākā (Nestor meridionalis septentrionalis) in Wellington, New Zealand. Animals 3(3):830-842. https://doi.org/10.3390/ani3030830
Corfield, J. R., A. C. Gsell, D. Brunton, C. P. Heesy, M. I. Hall, M. L. Acosta, and A. N. Iwaniuk. 2011. Anatomical specializations for nocturnality in a critically endangered parrot, the kakapo (Strigops habroptilus). PLoS ONE 6(8):e22945. https://doi.org/10.1371/journal.pone.0022945
Cox, D. T., and K. J. Gaston. 2024. Cathemerality: a key temporal niche. Biological Reviews 99(2):329-347. https://doi.org/10.1111/brv.13024
Daan, S. 1981. Adaptive daily strategies in behavior. Pages 275-298 in J. Aschoff, editor. Biological rhythms. Springer, New York, New York, USA. https://doi.org/10.1007/978-1-4615-6552-9_15
de Matos Dias, D., C. B. de Campos, and F. H. Guimarães Rodrigues. 2018. Behavioral ecology in a predator-prey system. Mammalian Biology 92:30-36. https://doi.org/10.1016/j.mambio.2018.04.005
Dickerson, A. L., M. L. Hall, and T. M. Jones. 2022. The effect of natural and artificial light at night on nocturnal song in the diurnal Willie Wagtail. Science of the Total Environment 808:151986. https://doi.org/10.1016/j.scitotenv.2021.151986
Dickerson, A. L., M. L. Hall, and T. M. Jones. 2020. The effect of variation in moonlight on nocturnal song of a diurnal bird species. Behavioral Ecology and Sociobiology 74:109. https://doi.org/10.1007/s00265-020-02888-z
Ellis, S., and S. Marsland. 2022. Sounding out the nest. New Zealand Journal of Ecology 46(1):1-11.
Finley, Jr, R. B. 1959. Observation of nocturnal animals by red light. Journal of Mammalogy 40(4):591-594. https://doi.org/10.2307/1376280
Fisher, C. D., E. Lindgren, and W. R. Dawson. 1972. Drinking patterns and behavior of Australian desert birds in relation to their ecology and abundance. Condor 74(2):111-136. https://doi.org/10.2307/1366276
Foote, J. R., L. K. Nanni, and R. Schroeder. 2017. Seasonal patterns of nocturnal singing by Ovenbirds and White-throated Sparrows. Behavior 154(13-15):1275-1295. https://doi.org/10.1163/1568539X-00003468
Forrest, S. W., M. R. Recio, and P. J. Seddon. 2022. Moving wildlife tracking forward under forested conditions with the SWIFT GPS algorithm. Animal Biotelemetry 10:19. https://doi.org/10.1186/s40317-022-00289-9
Fuller, R. A., P. H. Warren, and K. J. Gaston. 2007. Daytime noise predicts nocturnal singing in urban robins. Biology Letters 3(4):368-370. https://doi.org/10.1098/rsbl.2007.0134
Gaston, K. J. 2019. Nighttime ecology: the “nocturnal problem” revisited. American Naturalist 193(4):481-502. https://doi.org/10.1086/702250
Gaviraghi Mussoi, J., M. C. Stanley, and K. E. Cain. 2022. Importance of sleep for avian vocal communication. Biology Letters 18(8):20220223. https://doi.org/10.1098/rsbl.2022.0223
Geary, M., C. J. Brailsford, L. I. Hough, F. Baker, S. Guerrero, Y. M. Leon, N. J. Collar, and S. J. Marsden. 2021. Street-level green spaces support a key urban population of the threatened Hispaniolan parakeet Psittacara chloropterus. Urban Ecosystems 24(6):1371-1378. https://doi.org/10.1007/s11252-021-01119-1
Gibbs, G. W. 1998. Why are some weta (Orthoptera: Stenopelmatidae) vulnerable yet others are common? Journal of Insect Conservation 2(3):161-166. https://doi.org/10.1023/A:1009660200402
Gilardi, J. D., and C. A. Munn. 1998. Patterns of activity, flocking, and habitat use in parrots of the Peruvian Amazon. Condor 100(4):641-653. https://doi.org/10.2307/1369745
Gillings, S., and C. Scott. 2021. Nocturnal flight calling behaviour of thrushes in relation to artificial light at night. Ibis 163(4):1379-1393. https://doi.org/10.1111/ibi.12955
Glaz, J., and C. P. Sison. 1999. Simultaneous confidence intervals for multinomial proportions. Journal of Statistical Planning and Inference 82(1-2):251-262. https://doi.org/10.1016/S0378-3758(99)00047-6
Goodman, L. A. 1964. Simultaneous confidence intervals for contrasts among multinomial populations. Annals of Mathematical Statistics 35:716-725. https://doi.org/10.1214/aoms/1177703569
Groom, C. J., P. R. Mawson, J. D. Roberts, and N. J. Mitchell. 2014. Meeting an expanding human population’s needs whilst conserving a threatened parrot species in an urban environment. WIT Transactions on Ecology and the Environment 191:1199-1212.
Gruber, T., L. Luncz, J. Mörchen, C. Schuppli, R. L. Kendal, and K. Hockings. 2019. Cultural change in animals: a flexible behavioural adaptation to human disturbance. Palgrave Communications 5:64. https://doi.org/10.1057/s41599-019-0271-4
Halle, S., and N. C. Stenseth, editors. 2000. Activity patterns in small mammals: an ecological approach. Springer-Verlag, Berlin, Germany. https://doi.org/10.1007/978-3-642-18264-8
Harms, K. E., and J. R. Eberhard. 2003. Roosting behavior of the Brown-throated Parakeet (Aratinga pertinax) and roost locations on four southern Caribbean islands. Ornitologia Neotropical 14(1):79-89.
Harrison, M. E., E. R. Vogel, H. C. Morrogh-Bernard, and M. A. Van Noordwijk. 2009. Methods for calculating activity budgets compared: a case study using orangutans. American Journal of Primatology 71(4):353-358. https://doi.org/10.1002/ajp.20655
Hepworth, G., and A. Hamilton. 2001. Scan sampling and waterfowl activity budget studies: design and analysis considerations. Behavior 138(11-12):1391-1405. https://doi.org/10.1163/156853901317367654
Higgins, P. J. 1999. Handbook of Australian, New Zealand and Antarctic Birds. Volume 4, Parrots to Dollarbird. Oxford University Press, Melbourne, Australia.
Holdaway, R. N. 1999. Introduced predators and avifaunal extinction in New Zealand. Pages 189-238 in R. D. E. MacPhee, editor. Extinctions in near time: causes, contexts, and consequences. Springer, New York, New York, USA. https://doi.org/10.1007/978-1-4757-5202-1_9
Hothorn, T., K. Hornik, M. A. van de Wiel, and A. Zeileis. 2008. Implementing a class of permutation tests: the coin package. Journal of Statistical Software 28(8):1-23. https://doi.org/10.18637/jss.v028.i08
Iwaniuk, A. N., A. R. Keirnan, H. Janetzki, K. Mardon, S. Murphy, N. P. Leseberg, and V. Weisbecker. 2020. The endocast of the Night Parrot (Pezoporus occidentalis) reveals insights into its sensory ecology and the evolution of nocturnality in birds. Scientific Reports 10(1):9258. https://doi.org/10.1038/s41598-020-65156-0
Johnson, F. N., R. C. Shaw, and P. M. Wehi. 2025. Supporting biocultural connections in conservation translocations. Biological Conservation 302:110937. https://doi.org/10.1016/j.biocon.2024.110937
Johnsson, R. D., F. Connelly, J. Gaviraghi Mussoi, A. L. Vyssotski, K. E. Cain, T. C. Roth, and J. A. Lesku. 2022. Sleep loss impairs cognitive performance and alters song output in Australian magpies. Scientific Reports 12(1):6645. https://doi.org/10.1038/s41598-022-10162-7
Kahlert, J., A. D. Fox, and H. Ettrup. 1996. Nocturnal feeding in moulting Greylag Geese Anser anser—an anti-predator response. Ardea 84(1-2):15-22.
Karniski, C., E. M. Patterson, E. Krzyszczyk, V. Foroughirad, M. A. Stanton, and J. Mann. 2015. A comparison of survey and focal follow methods for estimating individual activity budgets of cetaceans. Marine Mammal Science 31(3):839-852. https://doi.org/10.1111/mms.12198
King, H., and C. Kemp. 2021. The impact of enrichment on the behaviour of kea (Nestor notabilis) at Auckland Zoo. Environmental and Animal Sciences, Unitec Institute of Technology, Auckland, New Zealand. https://hdl.handle.net/10652/5582
Kronfeld-Schor, N., and T. Dayan. 2003. Partitioning of time as an ecological resource. Annual Review of Ecology, Evolution, and Systematics 34(1):153-181. https://doi.org/10.1146/annurev.ecolsys.34.011802.132435
Kronfeld-Schor, N., D. Dominoni, H. De la Iglesia, O. Levy, E. D. Herzog, T. Dayan, and C. Helfrich-Forster. 2013. Chronobiology by moonlight. Proceedings of the Royal Society B: Biological Sciences 280(1765):20123088. https://doi.org/10.1098/rspb.2012.3088
Kühl, H. S., C. Boesch, L. Kulik, F. Haas, M. Arandjelovic, P. Dieguez, G. Bocksberger, M. B. McElreath, A. Agbor, and A. Angedakin, et al. 2019. Human impact erodes chimpanzee behavioral diversity. Science 363(6434):1453-1455. https://doi.org/10.1126/science.aau4532
La, V. T. 2012. Diurnal and nocturnal birds vocalize at night: a review. Condor 114(2):245-257. https://doi.org/10.1525/cond.2012.100193
Laiolo, P., and J. L. Tella. 2007. Erosion of animal cultures in fragmented landscapes. Frontiers in Ecology and the Environment 5(2):68-72. https://doi.org/10.1890/1540-9295(2007)5[68:EOACIF]2.0.CO;2
Lind, O., T. Sunesson, M. Mitkus, and A. Kelber. 2012. Luminance-dependence of spatial vision in budgerigars (Melopsittacus undulatus) and Bourke’s parrots (Neopsephotus bourkii). Journal of Comparative Physiology A 198(1):69-77. https://doi.org/10.1007/s00359-011-0689-7
Loepelt, J., R. C. Shaw, and K. C. Burns. 2016. Can you teach an old parrot new tricks? Cognitive development in wild kaka (Nestor meridionalis). Proceedings of the Royal Society B: Biological Sciences 283(1832):20153056. https://doi.org/10.1098/rspb.2015.3056
Mann, J. 1999. Behavioral sampling methods for cetaceans: a review and critique. Marine Mammal Science 15(1):102-122. https://doi.org/10.1111/j.1748-7692.1999.tb00784.x
Marín Gómez, O. H. 2022. Artificial light at night drives earlier singing in a neotropical bird. Animals 12(8):1015. https://doi.org/10.3390/ani12081015
Martin, G. 1990. Birds by night. T. and A. D. Poyser, London, UK.
Martin, G. R. 2017. The sensory ecology of birds. Oxford University Press, Oxford, UK. https://doi.org/10.1093/oso/9780199694532.001.0001
McFarland, D. C. 1991. The biology of the Ground Parrot, Pezoporus wallicus, in Queensland. I. Microhabitat use, activity cycle and diet. Wildlife Research 18(2):169-184. https://doi.org/10.1071/WR9910169
Mennill, D. J. 2014. Variation in the vocal behavior of Common Loons (Gavia immer): insights from landscape-level recordings. Waterbirds 37(sp1):26-36. https://doi.org/10.1675/063.037.sp105
Miskelly, C., R. Empson, and K. Wright. 2005. Forest birds recolonising Wellington. Notornis 52(1):21-26. https://doi.org/10.63172/195201kvvyml
Mitchell, W. F., and R. H. Clarke. 2019. Using infrared thermography to detect night-roosting birds. Journal of Field Ornithology 90(1):39-51. https://doi.org/10.1111/jofo.12285
Monterroso, P., P. C. Alves, and P. Ferreras. 2013. Catch me if you can: diel activity patterns of mammalian prey and predators. Ethology 119(12):1044-1056. https://doi.org/10.1111/eth.12156
Moorhouse, R. J. 1997. The diet of the North Island kaka (Nestor meridionalis septentrionalis) on Kapiti Island. New Zealand Journal of Ecology 21(2):141-152.
Morelli, F., V. Brlík, Y. Benedetti, R. Bussière, L. Moudrá, J. Reif, and M. Svitok. 2022. Detection rate of bird species and what it depends on: tips for field surveys. Frontiers in Ecology and Evolution 9:671492. https://doi.org/10.3389/fevo.2021.671492
Morrison, M. L. 1984. Influence of sample size and sampling design on analysis of avian foraging behavior. Condor 86(2):146-150. https://doi.org/10.2307/1367029
Mukhin, A. 2004. Night movements of young reed warblers (Acrocephalus scirpaceus) in summer: is it post-fledging dispersal? Auk 121(1):203-209. https://doi.org/10.1093/auk/121.1.203
Mukhin, A., V. Grinkevich, and B. Helm. 2009. Under cover of darkness: nocturnal life of diurnal birds. Journal of Biological Rhythms 24(3):225-231. https://doi.org/10.1177/0748730409335349
Mukhin, A., V. Kosarev, and P. Ktitorov. 2005. Nocturnal life of young songbirds well before migration. Proceedings of the Royal Society B: Biological Sciences 272(1572):1535-1539. https://doi.org/10.1098/rspb.2005.3120
Munita, C., T. A. Tadich, and C. Briceño. 2016. Comparison of two behavioral sampling methods to establish a time budget in a captive female cheetah (Acinonyx jubatus). Journal of Veterinary Behavior 13:1-5. https://doi.org/10.1016/j.jveb.2016.03.003
Nakamura-Garcia, M. T., and A. A. Ríos-Chelén. 2022. More than noise: light, moon phase, and singing behavior in a passerine. Urban Ecosystems 25(1):291-303. https://doi.org/10.1007/s11252-021-01142-2
NIWA. 2024. Climate summaries. NIWA, New Zealand. https://niwa.co.nz/climate-and-weather/climate-summaries
Nordt, A., and R. Klenke. 2013. Sleepless in town—drivers of the temporal shift in dawn song in urban European blackbirds. PLoS ONE 8(8):e71476. https://doi.org/10.1371/journal.pone.0071476
Penndorf, J., D. R. Farine, J. M. Martin, and L. M. Aplin. 2025. Parrot politics: social decision-making in wild parrots relies on both individual recognition and intrinsic markers. Royal Society Open Science 12(5):241542. https://doi.org/10.1098/rsos.241542
Pollard, K. A., and D. T. Blumstein. 2008. Time allocation and the evolution of group size. Animal Behavior 76(5):1683-1699. https://doi.org/10.1016/j.anbehav.2008.08.006
Powlesland, R. G., T. C. Greene, P. J. Dilks, R. J. Moorhouse, L. R. Moran, G. Taylor, A. Jones, D. E. Wills, C. K. August, and A. C. August. 2009. Breeding biology of the New Zealand kaka (Nestor meridionalis) (Psittacidae, Nestorinae). Notornis 56(1):11-33. https://doi.org/10.63172/399927iwpuxt
Pöysä, H. 1991. Measuring time budgets with instantaneous sampling: a cautionary note. Animal Behavior 42(2):317-318. https://doi.org/10.1016/S0003-3472(05)80564-0
Prior, L., K. Aland, A. L. Levengood, and D. A. Potvin. 2023. Vocal activity of the Eastern Ground Parrot (Pezoporus wallicus wallicus) and implications for acoustic monitoring efforts. Emu-Austral Ornithology 123(4):364-369. https://doi.org/10.1080/01584197.2023.2269575
R Core Team. 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://doi.org/10.32614/R.manuals
Rahlfs, M., and C. Fichtel. 2010. Anti-predator behavior in a nocturnal primate, the grey mouse lemur (Microcebus murinus). Ethology 116(5):429-439. https://doi.org/10.1111/j.1439-0310.2010.01756.x
Recio, M. R., K. Payne, and P. J. Seddon. 2016. Emblematic forest dwellers reintroduced into cities: resource selection by translocated juvenile kaka. Current Zoology 62(1):15-22. https://doi.org/10.1093/cz/zov010
Reilly, M. P. 2011. Māori studies, past and present: a review. Contemporary Pacific 23(2):340-370. https://doi.org/10.1353/cp.2011.0039
Renton, K., A. Salinas-Melgoza, M. Á. De Labra-Hernández, and S. M. de la Parra-Martínez. 2015. Resource requirements of parrots: nest site selectivity and dietary plasticity of Psittaciformes. Journal of Ornithology 156:73-90. https://doi.org/10.1007/s10336-015-1255-9
Riede, S. J., V. van der Vinne, and R. A. Hut. 2017. The flexible clock: predictive and reactive homeostasis, energy balance and the circadian regulation of sleep-wake timing. Journal of Experimental Biology 220(5):738-749. https://doi.org/10.1242/jeb.130757
Riley, M. 2001. Māori bird lore: an introduction. Viking Sevenseas NZ Ltd., Paraparaumu, New Zealand.
Robertson, H. A., K. A. Baird, G. Elliott, R. Hitchmough, N. McArthur, T. Makan, C. M. Miskelly, C. F. J. O’Donnell, P. M. Sagar, R. P. Scofield, G. A. Taylor, and P. Michel. 2021. Conservation status of birds in Aotearoa New Zealand, 2021. Department of Conservation, Wellington, New Zealand.
Roncero, P., O. de Mendonca-Furtado, and P. Izar. 2023. Human-induced rapid environmental change: a case study showing negative impact on animal culture. Journal for Nature Conservation 74:126424. https://doi.org/10.1016/j.jnc.2023.126424
Rose, L. 2000. Behavioral sampling in the field: continuous focal versus focal interval sampling. Behavior 137(2):153-180. https://doi.org/10.1163/156853900502006
Roth, T., P. Sprau, R. Schmidt, M. Naguib, and V. Amrhein. 2009. Sex-specific timing of mate searching and territory prospecting in the nightingale: nocturnal life of females. Proceedings of the Royal Society B: Biological Sciences 276(1664):2045-2050. https://doi.org/10.1098/rspb.2008.1726
Salinas-Melgoza, A., V. Salinas-Melgoza, and T. F. Wright. 2013. Behavioral plasticity of a threatened parrot in human-modified landscapes. Biological Conservation 159:303-312. https://doi.org/10.1016/j.biocon.2012.12.013
Santos, C. D., P. M. Lourenço, A. C. Miranda, J. P. Granadeiro, and J. M. Palmeirim. 2008. Birds after dark: an efficient and inexpensive system for making long-range observations at night. Journal of Field Ornithology 79(3):329-335. https://doi.org/10.1111/j.1557-9263.2008.00168.x
Shute, E. R., A. M. Clement, and G. J. Prideaux. 2023. Cranial adaptations of the Night Parrot (Psittaculidae: Pezoporus occidentalis), a cryptic nocturnal bird. Emu-Austral Ornithology 123(2):129-139. https://doi.org/10.1080/01584197.2023.2181185
Silk, M. J., D. P. Croft, T. Tregenza, and S. Bearhop. 2014. The importance of fission-fusion social group dynamics in birds. Ibis 156(4):701-715. https://doi.org/10.1111/ibi.12191
Sison, C. P., and J. Glaz. 1995. Simultaneous confidence intervals and sample size determination for multinomial proportions. Journal of the American Statistical Association 90(429):366-369. https://doi.org/10.1080/01621459.1995.10476521
Sourav, M. S. H., P. M. Thompson, and K. F. Biswas. 2018. Population and behavioural ecology of Alexandrine Parakeet Psittacula eupatria in Dhaka city, Bangladesh. Forktail 34:22-28.
Tesar, K., E. W. Heymann, and S. Dolotovskaya. 2023. A comparison of scan and focal sampling in estimating activity budgets, diet composition, and proximity patterns of a wild pair-living primate. Folia Primatologica 94(4-6):249-263. https://doi.org/10.1163/14219980-bja10016
Thomas, R. J. 2002. Seasonal changes in the nocturnal singing routines of Common Nightingales Luscinia megarhynchos. Ibis 144(2):E105-E112. https://doi.org/10.1046/j.1474-919X.2002.00070_7.x
van der Veen, D. R., S. J. Riede, P. D. Heideman, M. Hau, V. van der Vinne, and R. A. Hut. 2017. Flexible clock systems: adjusting the temporal programme. Philosophical Transactions of the Royal Society B: Biological Sciences 372(1734):20160254. https://doi.org/10.1098/rstb.2016.0254
Van Horik, J., B. Bell, and K. C. Burns. 2007. Vocal ethology of the North Island kaka (Nestor meridionalis septentrionalis). New Zealand Journal of Zoology 34(4):337-345. https://doi.org/10.1080/03014220709510093
Van Schaik, C. P. 2002. Fragility of traditions: the disturbance hypothesis for the loss of local traditions in orangutans. International Journal of Primatology 23(3):527-538. https://doi.org/10.1023/A:1014965516127
Vickery, P. D. 1996. Grasshopper Sparrow (Ammodramus savannarum), version 2.0. In A. F. Poole and F. Gill, editors. The birds of North America. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bna.239
Ward, M. P., M. Alessi, T. J. Benson, and S. J. Chiavacci. 2014. The active nightlife of diurnal birds: extraterritorial forays and nocturnal activity patterns. Animal Behavior 88:175-184. https://doi.org/10.1016/j.anbehav.2013.11.024
Weatherhead, P. J. 1983. Two principal strategies in avian communal roosts. American Naturalist 121(2):237-243. https://doi.org/10.1086/284053
Wilson, R. R., B. D. Jansen, and P. R. Krausman. 2008. Planning and assessment of activity budget studies employing instantaneous sampling. Ethology 114(10):999-1005. https://doi.org/10.1111/j.1439-0310.2008.01544.x
Wood, A. G. 1986. Diurnal and nocturnal territoriality in the Grey Plover at Teesmouth, as revealed by radio telemetry. Journal of Field Ornithology 57(3):213-221.
Wright, T. F. 1996. Regional dialects in the contact call of a parrot. Proceedings of the Royal Society of London. Series B: Biological Sciences 263(1372):867-872. https://doi.org/10.1098/rspb.1996.0128
Wright, T. F., and C. R. Dahlin. 2018. Vocal dialects in parrots: patterns and processes of cultural evolution. Emu-Austral Ornithology 118(1):50-66. https://doi.org/10.1080/01584197.2017.1379356
Wu, Y. 2020. Evolutionary origin of nocturnality in birds. Encyclopedia of Life Sciences 1:483-489. https://doi.org/10.1002/9780470015902.a0029073
Yambem, S. D., and M. Jain. 2023. Temporal variation in the behavior of a cooperatively breeding bird, Jungle Babbler (Argya striata). Tropical Ecology 64(1):133-145. https://doi.org/10.1007/s42965-022-00254-w
Yorzinski, J. L., and M. L. Platt. 2012. The difference between night and day: antipredator behavior in birds. Journal of Ethology 30:211-218. https://doi.org/10.1007/s10164-011-0318-5
Fig. 1
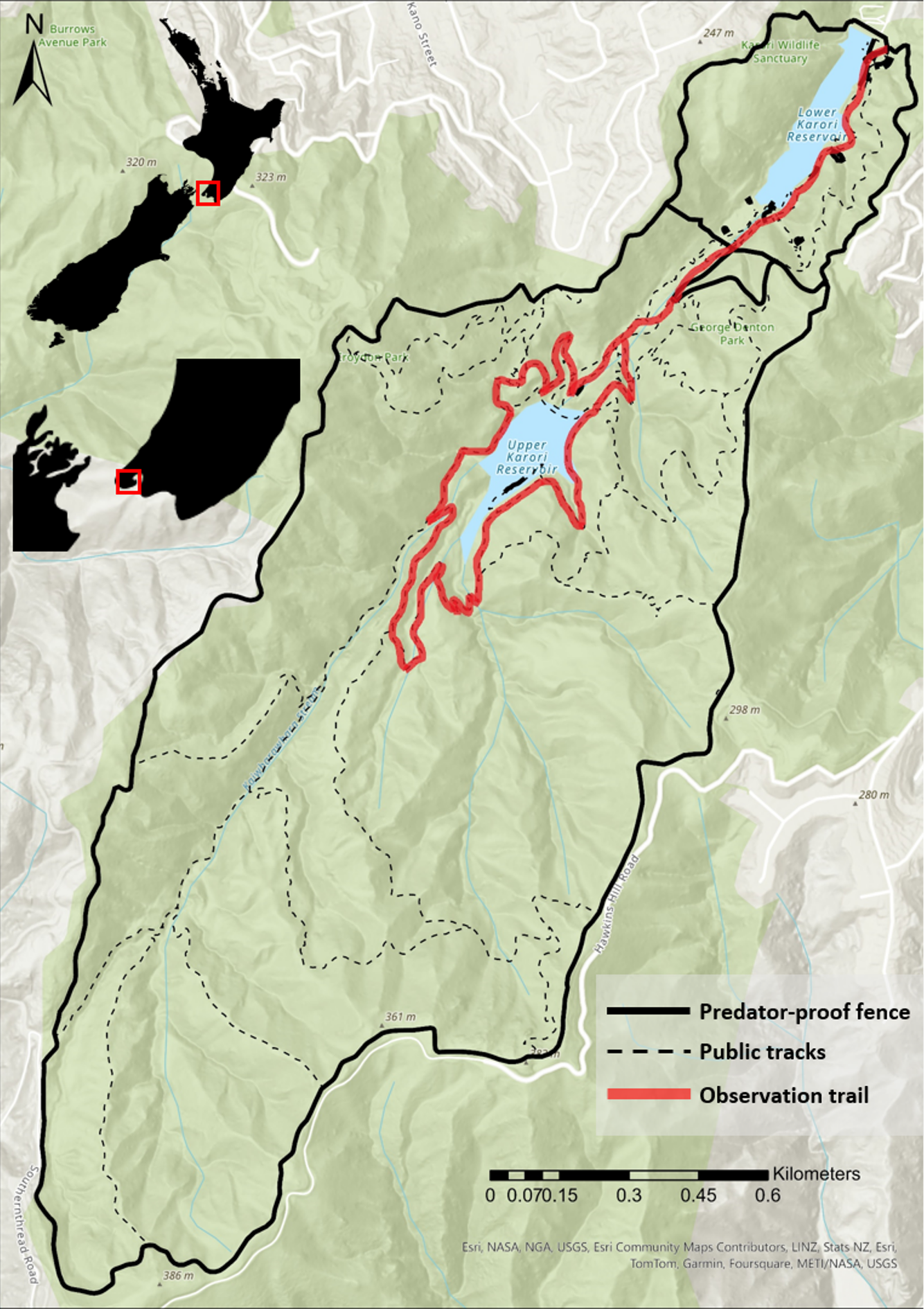
Fig. 1. Map of the observation trail within Zealandia Te Māra a Tāne, a predator-free wildlife sanctuary in Wellington city. Insets on the left indicate Zealandia’s location in New Zealand’s North Island.
Fig. 2
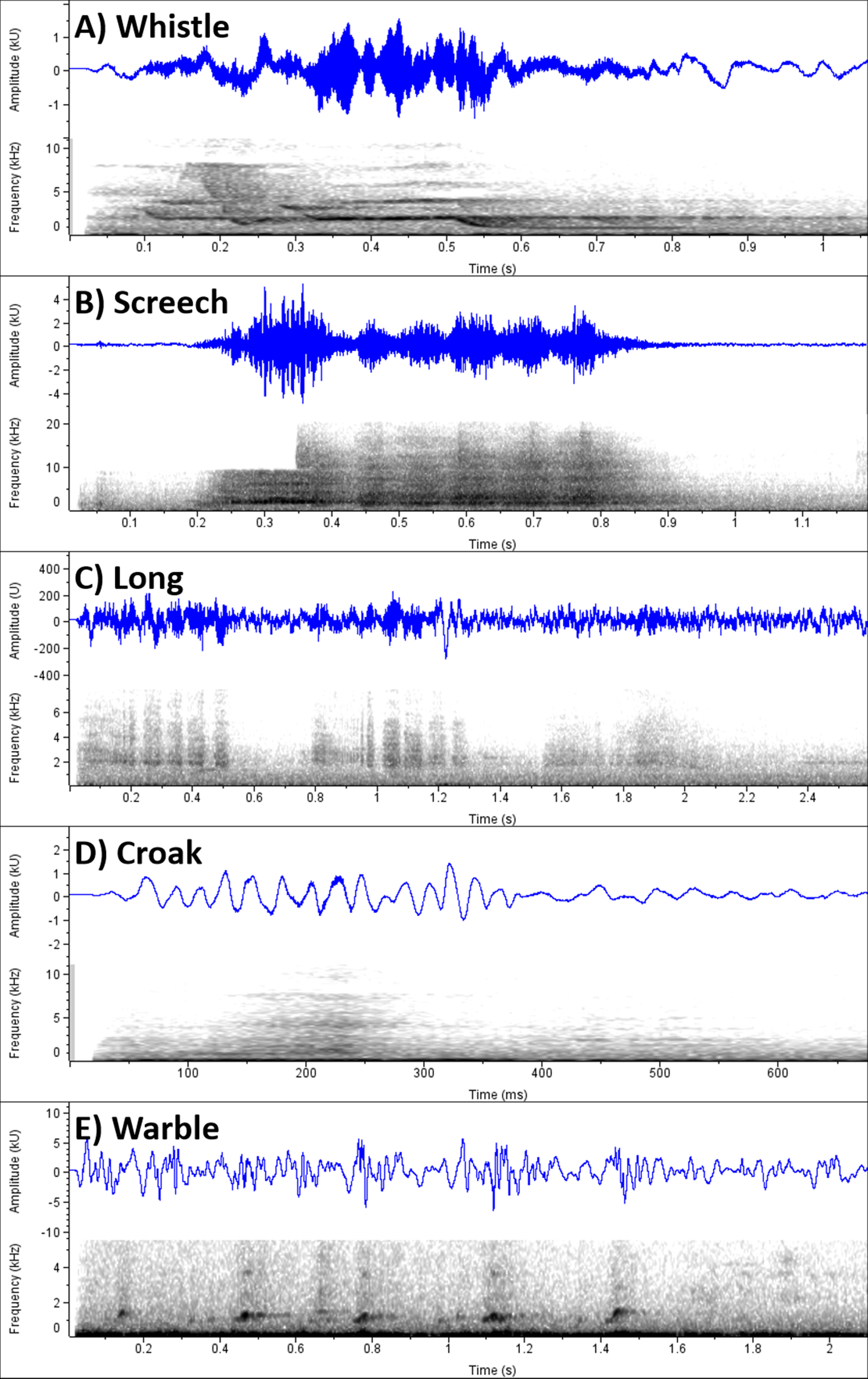
Fig. 2. Example spectrograms of the five call types identified in the field observations. Differences in the amplitude (blue line) and frequency (spectrograms) are shown separately for whistle (A), screech (B), long (C), croak (D), and warble (E) calls. The calls were recorded using a smartphone app and spectrograms were created using the software Raven Lite 2 (K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology 2023).
Fig. 3

Fig. 3. Examples of behaviors undertaken by Kākā (Nestor meridionalis) in Zealandia. Kākā foraging during day (A) and at night (B), grooming during the day (C) and at night (D), moving through the canopy during the day (E) and at night (F), and sleeping during the day (G) and at night (H).
Fig. 4

Fig. 4. Estimated percentage of time that Kākā (Nestor meridionalis) in Zealandia ecosanctuary were encountered engaging in the six behavioral categories at night and during daytime using ad libitum sampling. Error bars denote the simultaneous 95% confidence intervals.
Fig. 5
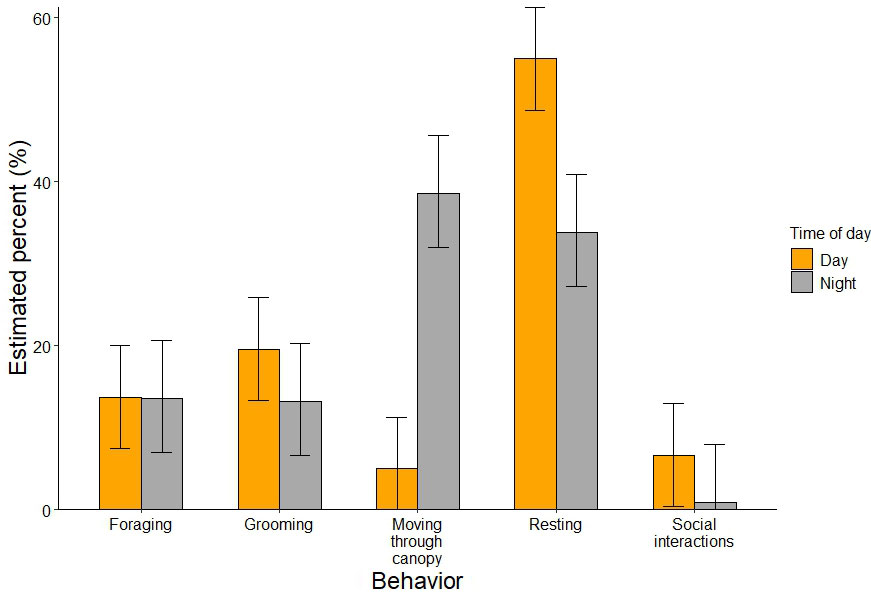
Fig. 5. Estimated activity budget of Kākā (Nestor meridionalis) in Zealandia ecosanctuary for the six behavioral categories at night and during daytime using instantaneous sampling. Error bars denote the simultaneous 95% confidence intervals. The category flying over canopy was excluded for this analysis.
Fig. 6
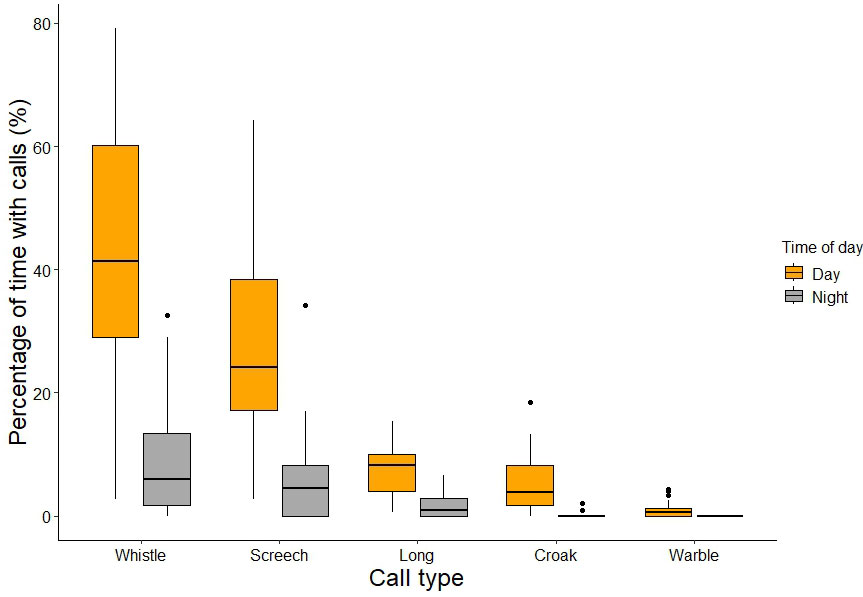
Fig. 6. Vocalization rate, defined as the percentage of sampling time in which calls were present, for each call type at night and during the day for Kākā (Nestor meridionalis) in Zealandia ecosanctuary. Because different call types often occur during the same sampling minute, the percentages do not add up to 100%.
Table 1
Table 1. Ethogram of behaviors of Kākā (Nestor meridionalis) observed in Zealandia ecosanctuary, Wellington, New Zealand, between February 2024 and October 2024. Behaviors are classified in six categories, and a description of the different call types is provided.
| Category | Behavior | Description | |||||||
| Flying above canopy | Flying above canopy | Flying above the canopy, often over a height of 50 m | |||||||
| Foraging | Drinking | Drinking water from a source on the ground | |||||||
| Eating pellets | Eating pellets from supplementary feeders | ||||||||
| Feeding | Engaging in feeding behavior: digging for wood-boring invertebrates, eating fruits or seeds, sap-feeding, visiting flowers for nectar, or eating pine seeds from pinecones | ||||||||
| Grooming | Bill-wiping | Wiping bill on branch with a figure-of-eight motion of the head | |||||||
| Head-scratching | Using claws to scratch the back of the head | ||||||||
| Self-preening | Cleaning feathers with beak | ||||||||
| Moving through canopy | Moving through canopy | Moving through the canopy using various methods in quick succession: walking, hopping, swinging with beak or claws, low flight within canopy | |||||||
| Resting | Perching | Perched at rest on branch with both feet and open eyes, not sleeping | |||||||
| Sleeping | Sleeping with eyes closed, head to one side, and perched on one foot | ||||||||
| Social interaction | Allopreening | Using bill to preen feathers of conspecifics | |||||||
| Calling (heard and seen) | Making calls while perched | ||||||||
| Courting | Male chasing the female in courtship ritual | ||||||||
| Copulating | Copulation where male has mounted the female | ||||||||
| Playing | Engaged in play behavior with conspecifics | ||||||||
| Chasing away Toutouwai (Petroica longipes) | Antagonistic altercation, disturbed by Toutouwai while foraging, chased it away | ||||||||
| Fighting with Tūī (Prosthemadera novaeseelandiae) | Antagonistic altercation with Tūī | ||||||||
| Call types (heard only) | Croak call | Low croaking call uttered while perched | |||||||
| Long call | Loud, harsh screech calls uttered in flight, mostly greater than 3 seconds in duration | ||||||||
| Screech call | Loud, grating single screech uttered mainly while perched but sometimes in flight | ||||||||
| Warble call | Repeating low-pitched whistles uttered in a sequence of 5 to 6 at a time | ||||||||
| Whistle call | Loud, high-pitched whistle uttered while perched or in flight uttered singly but may be repeated | ||||||||