







The following is the established format for referencing this article:
Guallar, S., J. Quesada, and J. C. Senar. 2026. Unscheduled plumage replacement in passerines: identification and implications for molt regulation. Journal of Field Ornithology 97(2):12.ABSTRACT
Molt is the periodic replacement of a bird’s plumage within the annual cycle. Replacements outside the known annual molt strategy of each species are considered accidental. During the winter of 2023–2024 in Barcelona city, we found evidence of a second plumage replacement in birds of three passerine species with only one known molt in their annual cycle: House Sparrow and Blue and Great tits. To evaluate how this fits our understanding of molt, we first considered whether these feathers could result from a delayed molt or from a new replacement occurring after the species’ scheduled molt. We ruled out the possibility of a delayed scheduled molt because the plumage was in fresh condition. We therefore examined whether these replacements could be treated as molt or as accidental feather replacement. To do so, we quantified two components of the molt process (intensity and sequence) and two components of the molt output (extent and asymmetry). Values of the four molt components were lowest in the House Sparrow, intermediate for the Blue Tit, and highest in the Great Tit. Using values of molt components, we propose three rules to determine whether unscheduled plumage replacements should be regarded as an actual extra molt episode or just accidental plumage replacement. From these rules, we concluded that the House Sparrow exhibited an accidental replacement or a highly asymmetrical extra molt episode, whereas both tit species were undergoing an extra molt episode, though asymmetrical in the Blue Tit. Unscheduled plumage replacement may not be an adaptive trait in some species, but could facilitate the evolution of more complex molt strategies and new plumage functions in other species.
RESUMEN
La muda es el reemplazo periódico del plumaje de un ave dentro del ciclo anual. Los reemplazos que ocurren fuera de la estrategia anual de muda conocida para cada especie se consideran accidentales. Durante el invierno de 2023–2024 en la ciudad de Barcelona, encontramos evidencia de un segundo reemplazo de plumaje en aves de tres especies de paseriformes con solo una muda conocida en su ciclo anual: Gorrión Común, Herrerillo Común y Carbonero Común. Para evaluar cómo esto se ajusta a nuestra comprensión de la muda, primero consideramos si estas plumas podrían resultar de una muda retrasada o de un nuevo reemplazo que ocurre después de la muda programada de la especie. Descartamos la posibilidad de una muda programada retrasada porque el plumaje se encontraba en condición fresca. Entonces, examinamos si estos reemplazos podían ser tratados como parte de la muda o como reemplazos accidentales de plumas. Para ello, cuantificamos dos componentes del proceso de muda (intensidad y secuencia) y dos componentes del resultado de la muda (extensión y asimetría). Los valores de los cuatro componentes de la muda fueron más bajos en el Gorrión Común, intermedios en el Herrerillo Común y más altos en el Carbonero Común. Usando los valores de los componentes de la muda, proponemos tres reglas para determinar si los reemplazos de plumaje no programados deben considerarse como un episodio adicional de muda o simplemente como reemplazos accidentales de plumas. A partir de estas reglas, concluimos que el Gorrión Común exhibió un reemplazo accidental o un episodio adicional de muda altamente asimétrico, mientras que ambas especies de páridos atravesaban un episodio adicional de muda, aunque asimétrico en el Herrerillo Común. El reemplazo no programado de plumas podría no representar un rasgo adaptativo en algunas especies, pero en otras podría favorecer la evolución de estrategias de muda más complejas y de nuevas funciones del plumaje.
INTRODUCTION
Bird molt can be defined as the non-accidental, programmed shedding and replacement of plumage (Jenni and Winkler 2020a). In passerines, it has a circannual periodicity, being partial or complete depending on age, season, and species molt strategy. Thus, in the Holarctic region, all passerine species undergo one complete molt per year as adults, usually right after breeding, although some long-distance migratory species delay this complete molt until after departing the breeding grounds. Whereas resident species typically undergo one molt per annual cycle, migratory species that winter in the tropics frequently undergo a second molt before returning to the breeding grounds (Svensson 1992, Pyle 2022). Molt timing has been evolutionarily set to optimize temporal and spatial variations in food availability (Barta et al. 2008), and fixed to use certain cues: gonadal regression and photoperiod for the complete molt of resident species (Dawson 2006, Pageau et al. 2021), breeding ground aridity for species that have a molt-migration strategy (Pageau et al. 2020), and food availability for the prealternate molt (Danner et al. 2015). The output of molt is constrained by time and resources (Bojarinova et al. 1999, Hall and Fransson 2000, Serra et al. 2024) and varies with season (Guallar and Jovani 2020).
As follows from its definition, the conceptualization of molt relies on prior knowledge of each species’ molt strategy. Any plumage growth occurring beyond the species’ scheduled molt is typically considered the result of aggression, predation attempts, or mechanical trauma, and is commonly referred to as accidental replacement (Willoughby et al. 2002). This conceptualization also relies on additional implicit assumptions. In particular, when the frequency, extent, or symmetry of plumage replacement within a population is low, the observed plumage growth is also often interpreted as accidental replacement rather than molt. However, these assumptions may fail when prior knowledge of a species’ molt strategy is limited, when there is intraspecific variability in molt strategy (Salewski et al. 2004, Lantz and Karubian 2016), or when potential evolutionary novelties emerge within the molt strategy of a population. In such cases, it is advisable to assess whether the observed plumage replacement is consistent with the elements that characterize molt. Because plumage replacement may be assessed either while it is active or after it has finished, elements of both the active molt process (e.g., intensity and sequence) and its output (e.g., extent and asymmetry) should be considered.
Intensity tends to follow a parabolic pattern as molt progresses (Wijnandts 1984, Guallar 2024) and is therefore expected to be low at the beginning and end of molt. Partial molt usually starts in body tracts, which show no clear sequence of activation (Willoughby 1986, Guallar and Quesada 2023), and is later followed by the flight-feather tracts. Whereas the sequence within tertials (divergent from the central one), alula, and rectrices (outward) is quite stable, it is less clear within the greater coverts (inward trend) and ill-defined within the median coverts (Guallar 2024). Consequently, the usefulness of sequence for assessing unscheduled plumage replacements will depend on their extent. The smaller the extent the less informative the sequence becomes. Moreover, since final extent cannot be predicted during the first stages of plumage replacement, this variable becomes informative only when extent is large enough or after the initial stages of plumage replacement. Finally, passerines usually show small lateral asymmetries during molt, and up to half of individuals in some species present asymmetries at the end of the process (Minias and Iciek 2013). Therefore, although asymmetry is usually low in scheduled partial molts, it may be higher in unscheduled plumage replacements because its regulation could be relaxed.
During the field work for two studies conducted in urban parks in the city of Barcelona (see Methods), we captured three birds that were replacing a substantial part of their plumage during the winter of 2023–2024: one House Sparrow (Passer domesticus), one Blue Tit (Cyanistes caeruleus), and one Great Tit (Parus major). These sedentary passerine species undergo one complete molt per year, although both tit species undergo a partial preformative molt during their first year of life (Svensson 1992). House Sparrow populations of the region largely complete molt in October, whereas the tit populations complete molt in September (ICO 2025). Therefore, these three birds were replacing plumage months after their scheduled molt, raising questions about how these observations fit our current understanding of molt. To address these questions, we set the following objectives:
- To evaluate whether unscheduled winter feather replacement is an overlooked phenomenon by estimating the frequency of winter plumage replacement for each species in the study area;
- To determine whether these cases represented delayed molt or a new episode of plumage replacement occurring after the species’ scheduled molt; and
- To assess whether this winter plumage replacement of plumage should be interpreted as molt by comparing intensity, sequence, extent, and asymmetry with those observed during the species’ scheduled molt.
Because our findings have implications for the regulation of molt, we also discuss the potential triggers and consequences of unscheduled episodes of plumage replacement.
METHODS
We gathered data from two ongoing population studies targeting House Sparrows (PASSERCAT project), and Great and Blue tits (MALLERENGUES project) in 16 urban parks of Barcelona city (Fig. 1). Between 1 December 2023 and 15 March 2024 (Table 1), we set up funnel traps allowing free access and ad libitum feeding of peanuts (Senar et al. 1997). The traps were deployed from approximately 08:00 until 14:00 h and checked hourly. Captured birds were banded, sexed, and aged (following Svensson 1992); wing chord and weight were measured; and the degree of feather growth on the body, wings, and tail was recorded. To assess the prevalence of winter plumage growth, we summarized its frequency among House Sparrows captured during a broader period of the PASSERCAT project (winters 2021–2025; Covas et al. 2017).
We scored the approximate percentage of feathers growing on seven body tracts (upper- and under-tail coverts, underwing coverts, scapulars, head, under- and upper-parts) and the growth stage of nine tracts on both wings and the tail of each bird (20 primaries, 12 secondaries, 6 tertials, 22 greater coverts, 16 median coverts, 18 primary coverts, 6 alula feathers, and 12 rectrices). We scored the approximate percentage of marginal coverts (expressed as a decimal) growing in the whole tract (see details in Guallar and Quesada 2021). The growth stage of each feather was scored on a continuous scale to account for differences in molt progress within feather tracts (Rohwer 2008): 0 for old feathers, 0.5 for shed feathers not yet emerging, 100 for fully grown feathers, while intermediate stages of feather growth were scored on the basis of the linear increase in feather length (Winkler et al. 1988). For example, a feather grown up to 50% of its full length was scored 50. This method is highly repeatable (Guallar and Quesada 2023).
From these data, we assessed two elements of the molt process (intensity and sequence) and two elements of the molt output (extent and asymmetry). Intensity of body was calculated as a weighted average percentage across tracts. Intensity of wing and tail feathers was computed as the percentage of growing feathers (total number = 114). Sequence profiles of wing and tail feathers were built by using growth percentages of feathers within each tract. For example, if primary P1 had 70% growth and primary P2 had 30% growth, P1 would be first in the primary molt sequence and P2 second. Extent was computed as the sheer number of feathers growing or fully grown during the current replacement of plumage. Observed asymmetry was calculated as the sum of all asymmetrically molting or molted feather positions in the wings and tail (Brommer et al. 2003). We considered only wing and tail tracts that contained either growing or fully grown feathers in each individual during the current plumage replacement. For example, if a bird had one median covert and one primary growing, and one fully grown greater covert, we included the three entire tracts in the calculations. For each observation, we calculated the expected asymmetry as the average asymmetry over 1000 random permutations by using the R package “pracma” (Borchers 2023, R Core Team 2024). Because asymmetry is a relative measure that depends on extent for each individual, it was calculated as the difference between observed and expected asymmetry. We neither kept birds in captivity nor recaptured them later on, therefore we could not infer final molt extent, which could have affected the final asymmetry.
RESULTS
During the winters of 2021–2025, the PASSERCAT project detected between 9% and 35% of House Sparrows growing body feathers (mean [range]= 47.6 [9-78] captures/winter), although only two of 238 (0.8%) were growing flight feathers. We captured 58 House Sparrows, 67 Blue Tits, 57 Great Tits, and 171 birds of seven other passerine species (i.e., bycatch) during the study period. The observed presence of winter replacement of plumage was 1.72%, 1.75%, and 1.5%, respectively. The overall percentage of winter replacement of plumage was 0.85%. Three of these birds were replacing a substantial portion of the plumage they had already replaced during their scheduled summer molt (assessment based on wear of the retained plumage).
House Sparrow D032065 was banded in Parc de la Ciutadella on 11 January 2024, a female of unknown age (fresh adult-type plumage) with a low plumage growth intensity: 12 pins growing in on the under- and upper-parts, four on the scapulars, and one on the under-tail coverts. On the right wing, the lesser tertial, secondary S2 and three greater coverts were growing, whereas none was growing on the left wing (Table 2, no photos available). Replacement did not proceed in a consecutive sequence (Fig. 2). Extent was low and asymmetry 100% (Table 2).
Blue Tit B070850 was banded in Parc del Laberint on 5 February 2024, a bird of unknown sex hatched the previous year (plumage with typical preformative molt limits on the wings and tail) showing a high plumage growth intensity: about 20% of its body feathers in a symmetrical fashion, from its cheeks, neck, and scapulars through its under- and upper-parts until the tail coverts. Also, about 10% marginal coverts were growing. Tail replacement was moderately symmetrical because the two central pairs of rectrices, plus rectrix R3 on the right side, were growing outwardly. However, replacement of wing feathers was highly asymmetrical: on the left wing, only the lesser tertial was growing, whereas on the right wing primaries P4 and P5 (P1 and P2 were fully grown), secondary S1, and the lesser alula feather were growing (Fig. 3, Table 2).
Great Tit C031451 was recaptured in Font del Racó on 20 December 2023, an adult male hatched before 2022 showing a high plumage growth intensity: about 25% of its body, 5% head, and six underwing coverts growing. Wing-feather growth was highly symmetrical and extent was large (though not finished yet), including all tracts typically molted during the species’ preformative molt: the two innermost tertials, nine inner greater coverts, the inner four median coverts, about 10% marginal coverts, and the central pair of rectrices were growing (Fig. 3, Table 2).
DISCUSSION
Unscheduled winter plumage replacement is a rare event in the Western Palearctic. It may go undetected because of the scarcity of winter banding projects and because such feather replacement is often considered accidental without further assessment. In fact, the frequency of accidental replacement may be substantial.
The three birds replacing plumage during winter had already completed their scheduled annual molt in summer-fall 2023. Therefore, this cannot be interpreted as delayed molt and should be considered either accidental replacement or an extra molt episode.
Although our sample size is limited and the molt protocol of the original projects have been discontinued, these observations provide the first documented evidence of winter plumage replacement in these species, and offer unique insights into the occurrence and regulation of unscheduled molts. Given the low frequency of the phenomenon, many years of systematic monitoring would be required to obtain additional records.
Consistency with scheduled molts
Although the very low frequency of unscheduled plumage replacement precludes the use of statistical inference, we can assess the consistency of each individual with its species’ scheduled molt. The low plumage growth intensity of House Sparrow D032065 and the presence of growing remiges and greater coverts is inconsistent with the initial stages of the molt progress in this species. Similarly, the discontinuous sequence within the greater coverts was inconsistent with the species’ typical inward sequence in this feather tract (Guallar and Quesada 2023).
Plumage replacement intensity of Blue Tit B070850 (hatched the previous breeding season) was consistent with a central stage of the molt progress. Wing-feather sequence, symmetry, and extent clearly departed from those of the species’ preformative molt, which typically does not include remiges (Fig. 2, Table 2; Jenni and Winkler 2020b). Although sequence within the rectrices was compatible with the species’ complete molt, the sequence within the primaries (right wing only) deviated from the typical complete molt because P3 was skipped. Atypically, primary molt was not accompanied by the molt of primary coverts (Fig. 3; Guallar 2024).
Plumage replacement intensity of Great Tit C031451 was consistent with a central stage of the molt progress. Sequence within the greater and median coverts, though not following the species’ overall sequence, was compatible with those of the species’ preformative molt (Fig. 2). Lack of growing alula feathers was also consistent with the species’ preformative molt, because they are the last to be activated in the molt sequence (Jenni and Winkler 2020b, Guallar 2024). Rectrix sequence (only the central pair growing) was fully compatible, whereas the outward tertial sequence was marginally compatible (exhibited by only 3 of 84 birds; Guallar 2024).
Telling apart an unscheduled molt from accidental replacement can be unfeasible during the first stages of plumage growth or when extent is very small. Accidental replacement has being characterized by asymmetrical and simultaneous plumage regrowth (Frohlich 2009, Guallar and Quesada 2021), although these assumptions should be reevaluated to take into account factors such as latitude and importance of feathers (de la Hera et al. 2015). Because distinguishing an unscheduled molt from accidental replacement can prove elusive, we propose three empirical rules to determine whether an unscheduled winter plumage replacement should be regarded as an actual extra molt episode:
- Body molt is symmetrical with an intensity > 10%.
- Sequence within wing flight-feather tracts is compatible with that of the species’ scheduled molt.
- Once plumage replacement is finished, body molt is symmetrical and affects at least 20% of plumage.
The higher these values, the greater the likelihood of the observed plumage replacement represents true molt. If multiple feather tracts are involved (especially unexposed ones like underwing coverts or axillaries), then the probability of accidental replacement is lower (Willoughby et al. 2002). Molt can be asymmetrical or anomalous (Norman 2003), and its expression might be unstable outside the scheduled period. For this reason, lateral symmetry alone might not be a diagnostic feature.
Based on these empirical rules, the unscheduled plumage replacement can be classified as an extra molt episode in the Great Tit, an extra asymmetrical molt episode in the Blue Tit, and either an accidental replacement or a highly asymmetrical extra molt episode in the House Sparrow. Applying these rules to a House Sparrow that was replacing 20% of its body plumage and 15 flight feathers on 15 February 2021 (Guallar and Quesada 2021), it should be classified as an extra anomalous molt episode.
Implications for molt regulation
Although molt is a programmed process of plumage replacement, unscheduled plumage replacement may occasionally occur. These anomalies are interesting because they can provide insight into both the triggers and the expression of molt. In this context, information on frequency of occurrence, timing, and identity of replaced feathers (i.e., the final molt phenotype; Guallar and Jovani 2020) is particularly relevant.
The triggers underlying unscheduled plumage replacement remain speculative. Given its very low frequency, they likely act in individual birds rather than at the population level. One potential congenital trigger is a mutation causing an extra zeitgeber (Gwinner 2003), which could be supported by recaptures in subsequent winters. Some studies have shown that birds begin molt earlier and exhibit reduced intensity in urban environments (Hope et al. 2016, Hutton et al. 2021). Artificial light at night can disrupt circadian and seasonal rhythms in birds, potentially advancing or delaying typical molt schedules (Dominoni et al. 2020), whereas exposure to endocrine-disrupting chemicals (e.g., in plastics; van Hassel et al. 2025) may also alter hormone-regulated processes. Severe stress might also induce feather loss; unlike scheduled molts, feather shedding is not forced by the emerging new feathers, so plumage regrowth may lag, potentially causing temporary bald patches (Kuenzel et al 2005). Birds can accidentally lose feathers from trauma, aggression by conspecifics, or predator attacks, regrowing them after a short interval (about one week in Barcelona during winter; unpublished results). The small and asymmetric extent of plumage loss in such cases is consistent with the plumage replacement observed in House Sparrow D032065.
Cases of unscheduled plumage replacement documented here are characterized by unexpected timing and by exogenous and endogenous conditions that differ markedly from those of scheduled molts. Great Tit C031451 is particularly interesting, as it was a molting adult exhibiting the intensity, sequence, and extent of the species’ preformative molt. This suggests that molt regulation under disparate conditions may trigger a stereotyped regulatory loop. However, if feather synthesis is activated at a suboptimal stage of the annual cycle, new feathers may be of lower quality, as observed for accidental replacements (de la Hera et al. 2010). Stage of the annual cycle is strongly associated with molt pattern in passerines, often differing between breeding and non-breeding stages (Jenni and Winkler 2020b, Pyle 2022), and correlates to different plumage functions (Guallar and Jovani 2020). Consequently, multiple differences arise between scheduled and unscheduled molts, including extent and pattern, likely caused by a disruption of the mechanisms of molt regulation. This may lead to strong asymmetries, as observed in Blue Tit B070850. Such asymmetries are associated with lower survival and increased flight costs (Brommer et al. 2003, Minias and Iciek 2013), whereas heat loss from highly vascularized feather quills is exacerbated during winter (Lustick 1970). Together, these two factors suggest that unscheduled plumage replacement during the cold season is not adaptive, although it may facilitate the evolution of more complex molt strategies and novel plumage functions (Terrill et al. 2020).
Our results highlight that unscheduled plumage replacement, though exceptionally rare, can occur in passerine species generally regarded as having a fixed annual molt cycle. By combining information on molt intensity, sequence, extent, and asymmetry, we provide a practical framework to distinguish between accidental feather replacement and true extra molt episodes. However, the small number of observations obtained during a single winter of surveys in a limited study area precludes drawing firm conclusions. Future studies based on long-term monitoring and repeated captures will be necessary to estimate its frequency, clarify its triggers, and assess the fitness impacts of this phenomenon in natural populations.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
S Guallar designed the study. S Guallar recorded the data. S Guallar and J Quesada analyzed the data. S Guallar wrote the paper with input from all co-authors. All authors read and approved the final manuscript. JC Senar and J Quesada contributed funding and materials.
ACKNOWLEDGMENTS
We are grateful to Pepe Carrillo and Míriam Crespo for their help during fieldwork. Comments by two anonymous reviewers greatly improved earlier versions of this article. This work was funded by CGL- 2020 PID2020- 114907GB- C21 research project to JCS from the Spanish Research Council, Ministerio de Ciencia e Innovación. Research funds were granted by the Natural Science Museum of Barcelona (PASSER-CAT project) to JQ.
DATA AVAILABILITY
The data and code that support the findings of this study are available on figshare: https://doi.org/10.6084/m9.figshare.30627578.v1.
LITERATURE CITED
Barta, Z., J. McNamara, A. I. Houston, T. P. Weber, A. Hedenström, and O. Feró. 2008. Optimal moult strategies in migratory birds. Philosophical Transactions of the Royal Society B 363:211-229. https://doi.org/10.1098/rstb.2007.2136
Bojarinova, J. G., E. Lehikoinen, and T. Eeva. 1999. Dependence of postjuvenile moult on hatching date, condition and sex in the and Great Tit. Journal of Avian Biology 30:437-446. https://doi.org/10.2307/3677016
Borchers, H. W. 2023. pracma: practical numerical math functions. R package version 2.4.4. https://doi.org/10.32614/CRAN.package.pracma
Brommer, J. E., O. Pihlajamäki, H. Kolunen, and H. Pietiäinen. 2003. Life-history consequences of partial-moult asymmetry. Journal of Animal Ecology 72(6):1057-1063. https://doi.org/10.1046/j.1365-2656.2003.00773.x
Covas, L., J. C. Senar, L. Roqué, and J. Quesada. 2017. Records of fatal attacks by Rose-banded Parakeets Psittacula krameri on native avifauna. Revista Catalana d’Ornitologia 33:45-49.
Danner, R. M., R. S. Greenberg, J. E. Danner, and J. R. Walters. 2015. Winter food limits timing of pre-alternate moult in a short-distance migratory bird. Functional Ecology 29(2):259-267. https://doi.org/10.1111/1365-2435.12322
Dawson, A. 2006. Control of molt in birds: association with prolactin and gonadal regression in starlings. General and Comparative Endocrinology 147(3):314-322. https://doi.org/10.1016/j.ygcen.2006.02.001
de la Hera, I., A. Hedenström, J. Pérez-Tris, and J. L. Tellería. 2010. Structural and mechanical differences between original and replaced feathers in Blackcaps Sylvia atricapilla. Ardeola 57(2):431-436.
de la Hera, I., J. Pérez-Tris, and J. L. Tellería. 2015. Adventitious feather replacement favours a more rapid regeneration of primaries over rectrices in two passerine bird species. Ibis 157(4):883-887. https://doi.org/10.1111/ibi.12302
Dominoni, D. M., J. A. H. Smit, M. E. Visser, and W. Halfwerk. 2020. Multisensory pollution: artificial light at night and anthropogenic noise have interactive effects on activity patterns of great tits (Parus major). Environmental Pollution 256:113314. https://doi.org/10.1016/j.envpol.2019.113314
Frohlich, D. 2009. Ageing North American landbirds by molt limits and plumage criteria. A photographic companion to the identification guide to North American birds, part I. Second edition. Slate Creek Press, Bolinas, California, USA.
Guallar, S. 2024. From partial to complete: wing- and tail-feather moult sequence and intensity depend on species, life-cycle stage, and molt completeness in passerines. Avian Research 15(1):100163. https://doi.org/10.1016/j.avrs.2024.100163
Guallar, S., and R. Jovani. 2020. Wing-feather moult phenotypes differ between the preformative and prealternate episodes and along passerine phylogeny. Ibis 162(3):778-786. https://doi.org/10.1111/ibi.12798
Guallar, S., and J. Quesada. 2021. Moulting in the wrong season? A House Sparrow replacing a significant part of its plumage in winter. Revista Catalana d’Ornitologia 37:34-41. https://doi.org/10.62102/20.8100.01.31
Guallar, S., and J. Quesada. 2023. Moult intensity constraints along the complete-moult sequence of the House Sparrow Passer domesticus. Avian Research 14(3):100125. https://doi.org/10.1016/j.avrs.2023.100125
Gwinner, E. 2003. Circannual rhythms in birds. Current Opinion in Neurobiology 13(6):770-778. https://doi.org/10.1016/j.conb.2003.10.010
Hall, K. S. S., and T. Fransson. 2000. Lesser Whitethroats under time-constraint moult more rapidly and grow shorter wing feathers. Journal of Avian Biology 31(4):583-587. https://doi.org/10.1034/j.1600-048X.2000.310419.x
Hope, S. F., F. A. Stabile, and L. K. Butler. 2016. Urban living alters moult dynamics in a passerine. Journal of Avian Biology 47:304-311. https://doi.org/10.1111/jav.00866
Hutton, P., J. McKenna, and K. J. McGraw. 2021. Urban links to molt schedule, body condition and carotenoid-based coloration in the house finch Haemorhous mexicanus. Journal of Avian Biology 52:e02761. https://doi.org/10.1111/jav.02761
ICO. 2025. SIOC: servidor d'informació ornitològica de Catalunya. ICO, Barcelona, Spain. http://www.sioc.cat
Jenni, L., and R. Winkler. 2020a. The biology of moult in birds. Helm, London, UK.
Jenni, L., and R. Winkler. 2020b. Moult and ageing of European passerines. Second edition. Bloomsbury, London, UK.
Kuenzel, W. J., R. F. Wideman, M. Chapman, C. Golden, and D. M. Hooge. 2005. A practical method for induced molting of caged layers that combines full access to feed and water, dietary thyroactive protein, and short day length. World’s Poultry Science 61(4):599-624. https://doi.org/10.1079/WPS200573
Lantz, S. M., and J. Karubian. 2016. Male Red-backed Fairywrens appear to enhance a plumage-based signal via adventitious molt. Auk 133(3):338-346. https://doi.org/10.1642/AUK-15-185.1
Lustick, S. 1970. Energy requirements of molt in cowbirds. Auk 87(4):742-746. https://doi.org/10.2307/4083708
Minias, P., and T. Iciek. 2013. Extent and symmetry of post-juvenile moult as predictors of future performance in Greenfinch Carduelis chloris. Journal of Ornithology 154:465-468. https://doi.org/10.1007/s10336-012-0911-6
Norman, D. 2003. Retrapped Nuthatch recovers from missed molt. 12 September 2003. https://www.merseysiderg.org.uk/Nuthatch%20recovers%20from%20missed%20moult.htm
Pageau, C., C. M. Tonra, M. Shaikh, N. J. Flood, and M. W. Reudink. 2020. Evolution of moult-migration is directly linked to aridity of the breeding grounds in North American passerines. Biology Letters 16(6):20200155. https://doi.org/10.1098/rsbl.2020.0155
Pageau, C., J. Sonnleitner, C. M. Tonra, M. Shaikh, and M. W. Reudink. 2021. Evolution of winter molting strategies in European and North American migratory passerines. Ecology and Evolution 11(19):13247-13258. https://doi.org/10.1002/ece3.8047
Pyle, P. 2022. Identification guide to North American birds. Part I. Second edition. Slate Creek Press, Forest Knolls, California, USA.
R Core Team. 2024. R: a language and environment for statistical computing. Vienna, R Foundation for Statistical Computing. https://doi.org/10.32614/R.manuals
Rohwer, S. 2008. A primer on summarizing molt data for flight feathers. Condor 110(4):799-806. https://doi.org/10.1525/cond.2008.8320
Salewski V., R. Altwegg, B. Erni, K. H. Falk, F. Bairlein, and B. Leisler. 2004. Molt of three Palaearctic migrants in their West African winter quarters. Journal of Ornithology 145:109-116. https://doi.org/10.1007/s10336-004-0020-2
Senar, J. C., J. Domènech, L. María, and E. Moreno. 1997. A funnel trap for the capture of tits. Butlletí del Grup Català d'Anellament 14:17-24.
Serra, L., M. Griggio, G. Casasole, S. Pirrello, L. Fusani, and A. Pilastro. 2024. Elevated testosterone levels during moult have contrasting effects on structural and carotenoid-based plumage colours in Eurasian Blue Tits. Ibis 167(1):263-276. https://doi.org/10.1111/ibi.13347
Svensson, L. 1992. Identification guide to European passerines. Fourth edition. L. Svensson, Stockholm, Sweden.
Terrill, R. S., G. F. Seeholzer, and J. D. Wolfe. 2020. Evolution of breeding plumages in birds. A multiple-step pathway to seasonal dichromatism in New World warblers (Aves: Parulidae). Ecology and Evolution 10(17):9223-9239. https://doi.org/10.1002/ece3.6606
Van Hassel, L., R. Felton, C. Tubbs, J. Beck, M. Albert, V. Baeten, J. A. Fernandez Pierna, C. Debier, and M. Finkelstein. 2025. Hormonal disruption from plastic ingestion in northern fulmars: activation and inhibition of estrogen receptors. Environmental Pollution 373:126145. https://doi.org/10.1016/j.envpol.2025.126145
Wijnandts, H. 1984. Ecological energetics of the Long-Eared Owl (Asio otus). Ardea 55(1-2):1-92. https://doi.org/10.5253/arde.v72.p1
Willoughby, E. J. 1986. An unusual sequence of molts and plumages in Cassin’s and Bachman’s sparrows. Condor 88(4):461-472. https://doi.org/10.2307/1368272
Willoughby, E. J., M. Murphy, and H. L. Gorton. 2002. Molt, plumage abrasion, and color change in Lawrence’s Goldfinch. Wilson Bulletin 114(3):380-392. https://doi.org/10.1676/0043-5643(2002)114[0380:MPAACC]2.0.CO;2
Winkler, R., W. D. Daunicht, and L. G. Underhill. 1988. Die Grossgefiedermauser von Alpendohle Pyrrhocorax graculus und Alpenkrähe Pyrrhocorax pyrrhocorax. Ornithologische Beobachter 85:245-259.
Fig. 1
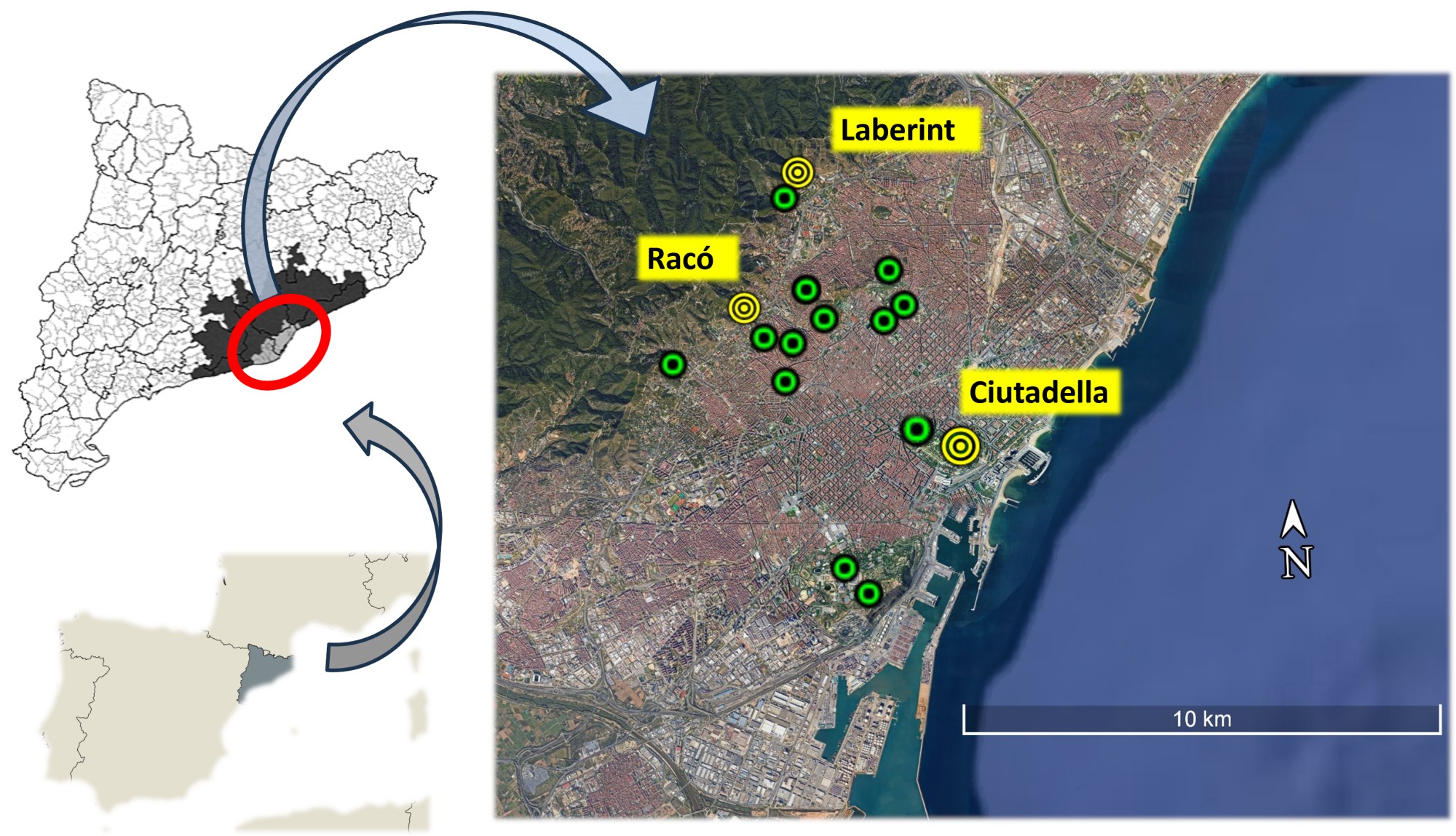
Fig. 1. Location of the 16 urban parks in the city of Barcelona (NE Iberian Peninsula) where sampling was conducted. Yellow points indicate the locations of the parks where birds undergoing plumage growth were captured during the winter of 2023–2024.
Fig. 2
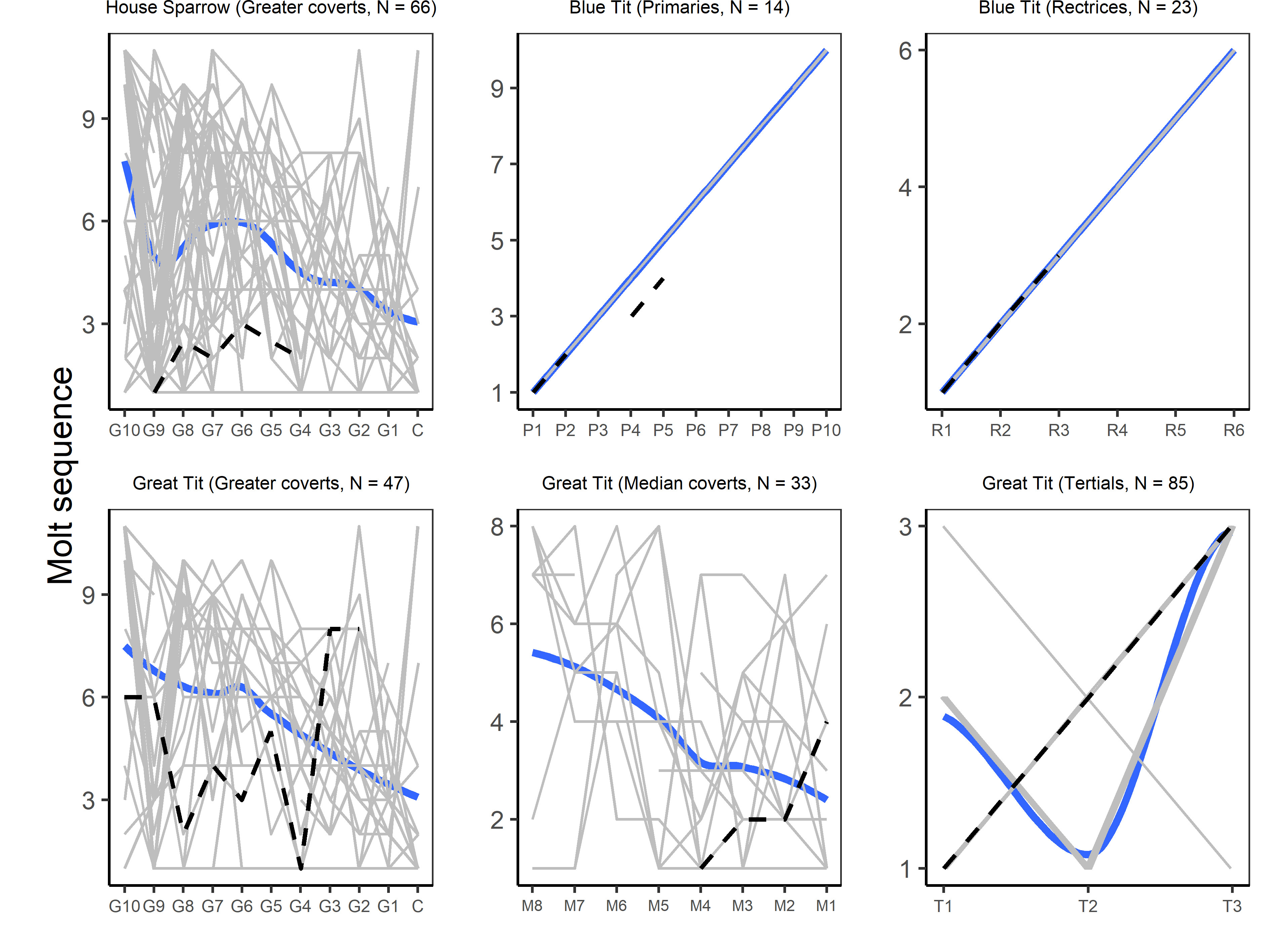
Fig. 2. Observed and fitted (polynomial regression using default parameters in base R) wing-feather sequence during scheduled molts of House Sparrows, Blue Tit, and Great Tit (data from Guallar 2024). Overlaid are the molt sequences of three individuals captured in urban parks in the city of Barcelona during the winter of 2023–2024. Primaries, tertials, and rectrices are numbered from the innermost outward on the right wing (Jenni and Winkler 2020b). Light gray lines represent the individual sequences observed in the sampled birds.
Fig. 3

Fig. 3. Winter plumage replacement in two tits captured in urban parks in the city of Barcelona. Top row: Blue Tit B070850 banded in Parc del Laberint on 5 February 2024. Primaries P4 and P5, secondary S1, and the lesser alula feather were growing (notice that primaries P1 and P2 were recently grown and that all primary coverts were juvenile). Rectrices R1 and R2 were recently grown, and R3 on the right side was growing in. Bottom row: Great Tit C031451 recaptured in Font del Racó on 20 December 2023. The two innermost tertials, nine inner greater coverts, the inner four median coverts, about 10% marginal coverts, and all rectrices were either missing or growing.
Table 1
Table 1. Frequency of site surveys in two population studies conducted in the city of Barcelona between 1 December 2023 and 15 March 2024. PASSERCAT surveyed the same two urban parks weekly during its survey period, whereas MALLERENGUES surveyed 16 parks a total of 40 times during the study period (range = 2–8 visits per park).
| Project | Sites surveyed per week | Survey period | |||||||
| PASSERCAT | 2 | 9 January–16 February | |||||||
| MALLERENGUES | 3 | 1 December–15 March | |||||||
Table 2
Table 2. Assessment of molt intensity, sequence, extent, and asymmetry for wing, tail, and body in three birds captured in urban parks in the city of Barcelona between 1 December 2023 and 15 March 2024.
| Wing/tail intensity (%) | Body intensity (%) | Sequential | Wing/tail extent (number) | Body asymmetry | Wing/tail asymmetry (%) | ||||
| House Sparrow (Passer domesticus) | 4.4 | 2 | no | 5 | yes | 100 | |||
| Blue Tit (Cyanistes caeruleus) | 10.5 | 20 | unclear | 12 | no | 79.5 | |||
| Great Tit (Parus major) | 28.9 | 20 | yes | 33 | no | 7.2 | |||