







The following is the established format for referencing this article:
Mirzadi, R. E., and D. E. Gawlik. 2026. Waterbird nest survival derived from drone surveys. Journal of Field Ornithology 97(2):11.ABSTRACT
Drone surveys provide accurate estimates of nest abundance and survival for ground-nesting waterbird species, but drones have not been used to estimate survival for waterbirds nesting in a canopied swamp habitat. We compared nest survival of Snowy Egret (Egretta thula), Great Egret (Ardea alba), Tricolored Heron (Egretta tricolor), Glossy Ibis (Plegadis falcinellus), and Cattle Egret (Ardea ibis) between traditional ground surveys and drone survey methods in freshwater wetland habitats of Florida, USA, in 2021. Overall, there were minimal differences in survival estimates between methods. However, drone-derived estimates of dark-colored waterbirds had lower accuracy than drone-derived estimates of white-colored waterbirds. Our results suggest that drone surveys provide comparable estimates to traditional ground-based methods for nest survival of waterbirds breeding in swamp habitat, and they offer the advantage of causing less disturbance to birds than traditional methods if flown at appropriate altitudes and following best practices.
RESUMEN
Los muestreos con drones proporcionan estimaciones precisas de la abundancia y supervivencia de nidos para especies de aves acuáticas que anidan en el suelo; sin embargo, los drones no se habían utilizado para estimar la supervivencia de aves acuáticas que anidan en hábitats de pantanos con dosel. Durante 2021, comparamos la supervivencia de nidos de la garza dedos dorados (Egretta thula), la garza blanca (Ardea alba), la garza tricolor (Egretta tricolor), el ibis de cara blanca (Plegadis falcinellus) y la garza ganadera (Ardea ibis) mediante métodos de muestreo tradicionales en tierra y muestreos con drones en hábitats de humedales de agua dulce en Florida, EE. UU. En general, se observaron diferencias mínimas en las estimaciones de supervivencia entre ambos métodos. No obstante, las estimaciones derivadas de drones para aves acuáticas de plumaje oscuro presentaron una menor precisión en comparación con aquellas obtenidas para aves acuáticas de plumaje blanco. Nuestros resultados sugieren que los muestreos con drones proporcionan estimaciones comparables a los métodos tradicionales basados en tierra para la supervivencia de nidos de aves acuáticas que se reproducen en hábitats de pantano. Asimismo, ofrecen la ventaja de causar un menor disturbio a las aves en comparación con los métodos tradicionales, siempre que se vuele a altitudes adecuadas y se sigan las mejores prácticas.
INTRODUCTION
Nest survival, the probability that a nest is successful in rearing at least one nestling to fledging (Dinsmore et al. 2002), has traditionally been a difficult demographic parameter to measure because it requires repeated assessments of the fate of individual nests across the breeding period. This difficulty is compounded for colonial-nesting waterbirds, which often form dense aggregations for a relatively short duration (Furness and Monaghan 1987), thereby requiring frequent repetitive researcher intrusion into a colony to monitor breeding pairs.
Traditionally, waterbird populations have been surveyed using crewed aerial and ground-based monitoring methods, each of which has its own limitations and biases. Ground surveys provide accurate estimates of productivity and survival for most waterbird species; however, ground monitoring techniques are expensive and may cause disturbance resulting in negative effects to reproduction (Frederick et al. 1996, Werschkul et al. 1976, Tremblay and Ellison 1979). Fixed-wing crewed aerial surveys are crucial in monitoring colonially nesting waterbirds; however, they are expensive, have been shown to have significant observer and detection bias (Frederick et al. 1996, 2003, Rodgers et al. 2005), and have been responsible for the majority of job-related deaths for wildlife biologists (Sasse 2003). Also, fixed-wing crewed aerial surveys are not reliable for identifying and tracking individual nests, which is necessary for estimating productivity and survival.
Recently, there has been an increased interest in the use of uncrewed aircraft vehicles (UAVs), hereafter referred to as drones, for environmental and wildlife monitoring applications (Abd-Elrahman et al. 2005, Jones et al. 2006, Chabot et al. 2015). Drones have proven effective in monitoring several avian species (Bevan et al. 2018, Vermeulen et al. 2013, Abd-Elrahman et al. 2005, Jones et al. 2006, Chabot et al. 2015). Drones are less disturbing to nesting birds than traditional survey methods (Chabot et al. 2015, Vas et al. 2015) and have produced higher detection rates of some colonially nesting birds than fixed-wing crewed aerial surveys (Afán et al. 2018, Chabot et al. 2015, McEvoy et al. 2016). Fixed-wing and multirotor drones (Colomina and Molina 2014) are available to the public and often used for avian research projects. Advantages of multirotor drones over fixed-wing drones are lower purchase price and the ability to take off and land vertically, which allows them to be launched easily from small landing areas such as a boat. Multirotor drones are also easy to fly, can be easily programmed to conduct autonomous flights, and can be flown at speeds slow enough to capture side and front overlap as high as the pilot stipulates (Thamm et al. 2015, Boon et al. 2017). In contrast, an advantage of fixed-wing drones over multirotor drones is higher flight endurance, leading to a larger flight area per battery set (Boon et al. 2017, Garg 2022).
Given recent advances in drones and post-processing technology, drone-derived aerial imagery can be used to relocate individual nests on subsequent surveys, allowing researchers to track individual nests throughout the season (Lachmen et al. 2020). Because drones can be programmed to fly fully autonomous missions, capturing images in specified locations with automatic or preset camera settings (Floreano and Wood 2015), images may be combined into a single orthomosaic image of an entire waterbird colony. An orthomosaic image is an accurate photographic representation of a specific area, created from overlapping photos stitched together and geometrically corrected for accuracy (Floreano and Wood 2015). Orthomosaic images can be crafted with survey-grade accuracy when drones are used in conjunction with a high-accuracy GPS and ground control points (Agüera-Vega et al. 2017). Because orthomosaic images can be produced with survey-grade accuracy, it is now possible to identify and track individual nests and juvenile birds throughout the breeding season. This allows estimates of productivity measures like nest survival to be recorded. For example, Lachmen et al. (2020) used drones to model nest survival of Western Grebes (Aechmophorus occidentalis), and Scarton and Valle (2020) used drones to model nest survival of Gull-billed Terns (Gelochelidon nilotica). However, no studies have used drones to estimate nest success and productivity of waterbird breeding colonies in an area with a high percentage of canopy cover, which can reduce detection rates of nest. Thus, the objectives of this study were to test for differences in survival estimates of waterbirds nesting in habitat with high canopy coverage derived from drone surveys and traditional ground-based surveys.
We used ground-based methods and drone-derived imagery to estimate survival for several species of waterbirds, Snowy Egret (Egretta thula), Tricolored Heron (Egretta tricolor), and Great Egret (Ardea alba), at colonies on Lake Okeechobee, Florida, USA, during the 2021 waterbird breeding season. Previous studies using drone-derived imagery to detect waterbirds have observed high detection rates of white-colored waterbird species (Barr et al. 2018). Additionally, studies have found lower detection rates of waterbird species nesting below the canopy, especially for dark-colored waterbird species (Chabot and Bird 2012, Barr et al. 2018, Orange et al. 2023). We hypothesized that there would be no difference between survival estimates derived from drone surveys and traditional ground-based surveys for white-colored species that nest above the canopy, which in our study was only the Great Egret. We expected that survival estimates derived from drone surveys would be higher than for traditional ground-based surveys for waterbird species that nest below the canopy, especially for dark-colored species. If drone surveys can be used to accurately estimate waterbird productivity in areas with high canopy cover, it will be possible to survey a greater number of colonies and nests with a higher degree of accuracy than with traditional methods. This increased efficiency will facilitate waterbird conservation and management by providing more reliable information for the same or lower costs.
METHODS
Study area
This study was conducted on the Lake Okeechobee in south central Florida, USA (26º56′28″ N, 80º51′32″ W). Lake Okeechobee (hereafter referred to as the lake) is a large (1720 km² surface area), shallow (mean depth of 2.7 m), eutrophic lake that lies at the center of the Greater Everglades Watershed (Johnson et al. 2007). Since at least the 1930s, the lake has periodically supported large colonies of wading birds (David 1994). Whereas water management between 1978–2007 increased lake levels, reduced the availability of nesting habitat, altered the availability of prey, and led to a reduction in the overall presence of breeding waterbirds on the lake by 60%, the adoption of the Lake Okeechobee Regulation Schedule in 2008 increased the abundance of breeding waterbirds on the lake (Essian et al. 2022). The lake currently supports the second largest nesting aggregation of nesting birds in South Florida, typically producing approximately 10% of all nests in the region each year (Cook and Baranski 2021).
Waterbirds typically nest on anthropogenic islands and natural wetland within the littoral zone of the lake, which is predominantly located on the Lake Okeechobee’s western edge (Cook and Baranski 2021). Within the littoral zone, plant communities are dominated by emergent marsh (Johnson et al. 2007). Dense stands of willow (Salix caroliniana), common reed (Phragmites australis) and pond apple (Annona glabra) are scattered on elevated ridges throughout the littoral zone and on several anthropogenic islands used by nesting waterbirds on the lake (Chastant et al. 2018).
We conducted surveys on four active waterbird colonies: two on anthropogenic islands located at the southwestern extent of the lake (Clewiston and Little Bear Beach) and two in natural wetlands at the northwestern part of the lake (Eagle Bay and Moonshine Bay). Colonies were surveyed throughout the entire breeding season (February through July), beginning when birds were first observed incubating eggs and ending when birds had ceased nesting.
Ground-based nest transects
At each monitored colony, we surveyed nests along two parallel transects 50 m by 3 m in size and spaced 30 m apart (e.g., Chastant and Gawlik 2018). We recorded species, contents (i.e., eggs and nestlings), nestling age, and fate for each marked nest during each nest transect survey. We used a random bearing and distance from the boat to select the origin of the first transect. The start and direction of each transect were adjusted if a random bearing extended beyond the edges of an island or passed through unsuitable nesting habitat.
Ground-based nest surveys were conducted every two–seven days. The length of time in between surveys was dictated by logistics, weather, and the age of nests. Researchers would overview the stage and age ascribed to nests in a colony at the end of every survey day and, if chicks were determined to be close to fledging through ground-based surveys, researchers would reduce the length of time in between surveys to ensure nests were recorded at 14 days old (± 2 days). This was intended to increase the sample size of nests that were found to reach fledging. The initiation of surveys in a colony was determined through the identification of nesting focal species, either determined through ground surveys conducted with an airboat, or monthly fixed-wing aerial surveys of the study area. Because the initiation and cessation of waterbird nesting on the lake differed among colonies, breeding colonies were not surveyed over the same range of dates nor surveyed the same number of times. The Clewiston colony was surveyed 20 times, between 9 March and 13 June. The Little Bear Beach Colony was surveyed 17 times, between 23 March and 9 June. The Moonshine Beach Colony was surveyed six times, between 9 May and 10 June. The Eagle Bay Colony was surveyed eight times, between 9 April and 13 May. We flagged and assigned an identification number to all accessible nests on the transect. If the sample size was below 50 nests per species on a transect, we extended the transect width an additional 3 m and marked nests within the wider transect. Contents, nestling age, species, and fate were recorded for each marked nest. Contents were recorded as the number of eggs and chicks observed in a nest. Nestling age was derived by adding the number of days past a nest’s estimated hatch date. Hatch date was determined from the estimated age of the largest nestling termed the “A chick”. The hatch date for nestling A was back-calculated to the first survey a nestling was observed in nest at which time the nestling was considered zero days old. Nests were considered successful, and the brood size was recorded for brood size at fledging, when chicks from a nest reached at least 14 days old (± two days; Frederick and Collopy 1989, Parsons and Master 2000). Fledging was recorded at 14 days (± two days) because, after the age of fourteen days, chicks are highly mobile and cannot be attributed to any particular nest (Billerman et al. 2020). The addition of a two-day threshold around the fledging age was used because of the difficulty of assigning some 14-day old chicks to their original nest and the infrequency of colony visits, which resulted in increased data retention (Chastant et al. 2017, Frederick et al. 2003). A Leica GS07 GNSS RTK Rover (Leica Geosystems, Heinrich-Wild-Strasse, Switzerland) was used to take GPS waypoints of each nest location. The transect, colony, and number associated with each nest were denoted in the name associated with each GPS waypoint.
All GPS waypoints were later imported to the orthomosaic image produced for the survey in Agisoft Metashape. Notes of nest locations, based on their orientation in the transect and ground photographs, were also used to positively identify individual nests.
Drone transect surveys
At the start of the season, and before the drone surveys began, we placed five Ground Control Points (GCPs) throughout each monitored colony to increase the geospatial accuracy of orthomosaic images stitched together during post-processing. Ground Control Points consisted of 60 cm by 60 cm water resistant plywood that had been painted to resemble a small checkerboard target. We placed GCPs atop steel pipes (1.2 m–3.0 m long) that had been positioned vertically into the ground in areas within each colony that were unobstructed by canopy or vegetation. We recorded GPS waypoints for each GCP using a Leica GS07 GNSS RTK Rover. We regularly checked GCPs to ensure no debris was obstructing their vertical presentation.
We conducted drone transect surveys immediately prior to ground-based nest transect surveys to mitigate waterbirds flushing from their nest. Drone transect surveys were conducted with a commercially available multirotor quadcopter drone (Inspire 2, DJI, Shenzhen, Guangdong, China) paired with a 24 mega-pixel RGB camera (Znemuse x7, DJI, Shenzhen, Guangdong, China). Images collected had a pixel resolution of 0.80 cm. The Inspire 2 may be operated by a single researcher, launched from almost any surface, and is operated using a modest ground station (iPad device and remote control; Apple Inc., Cupertino, CA).
The DJI Inspire 2 is rated to fly at a maximum wind speed of 38 km/h and, to ensure the safety of researchers and drone, we intended to only operate the drone when wind speeds were ≤ 38 km/h at the altitude at which missions were flown (77 m). Gusts commonly exceeded 38 km/h during drone transect surveys and, to ensure surveys were conducted at appropriate time intervals, it was necessary to increase the maximum wind speed during surveys to ≤ 46 km/h at the mission altitude (77 m). We accessed weather projections before each survey using a third-party app (UAV Forecast) and, if the wind exceeded 46 km/h, surveys were aborted.
Prior to conducting drone surveys, we selected a home base 185–750 m from the nearest nest to launch the drone. Longer distances, albeit still within line of sight, were sometimes needed because our study area was composed of large, often inaccessible tracks of thick emergent marsh. Once a home base was selected, we programmed the drone to photograph the transect using a drone mapping app (DJI GS Pro). The drone then flew to an altitude of 76 m and captured images of the transect at intervals with the appropriate overlap (90% front overlap and 90% side overlap). The drone continued capturing images until it recorded the entire transect. Upon the completion of the survey, we imported images to Agisoft Metashape, rectified them via Digital Surface Model/Digital Elevation Model, and stitched them into Orthomosaic images. During post-processing, we marked GCPs to ensure high geospatial accuracy of the orthomosaic image.
Image interpretation
We identified and tracked waterbird nests by photointerpretation using Agisoft Metashape’s orthomosaic window feature. Upon constructing an orthomosaic image, we systematically searched each nest transect in a grid pattern at a scale of 1:20. Active nests were distinguished following Sardà-Palomera et al. (2012). Because the orthomosaic images used in this study were produced by stitching together individual images captured using a seventy-eight-degree angle, it was not always possible to positively distinguish birds below the canopy as being on nests when using an orthomosaic image alone. Instead, because of the angle of the image and the amount and location of canopy cover over a nest, it was possible that a researcher was only able to view the back of a bird and not an actual nest. We labeled these as probable nests and placed a georeferenced reference marker directly over the location of a suspected nesting bird. We then selected the reference marker of the probable nest using the function Filter Photos by Marker in Agisoft Metashape. This function automatically selects every photo containing a referenced marker and zooms to the same magnification in every image. We examined every image containing a referenced marker and recorded whether the bird could be confirmed as on a nest from an individual image captured during the survey or in the same location in images captured throughout the survey. If it was not possible to confirm nesting through an individual image captured during the survey, we assumed a nest to be active if the individual did not move locations in consecutive images taken during the survey, at an interval between 15–45 minutes between the time images were captured. We tracked nests classified as active throughout the season. There were 29 instances when new nests were constructed on top of old nests after not being detected during previous weeks. We considered those nests to be new nests and marked them with a unique identification number.
Once a nest was confirmed as active through ground-based nest transects, we imported GPS coordinates of the nest’s location recorded with the RTK Rover into Agisoft Metashape. We then matched nests observed in Agisoft Metashape to nests marked through ground-based nest transects. The GPS coordinates of nests observed during ground-based nest transects but not detected through drone surveys were marked in the orthomosaic image, ensuring that the nest had been accounted for and had not been mistaken for another nest. We tracked active nests observed through drone surveys throughout the nesting season using the Filter Photos by Marker function in Agisoft Metashape. While tracking nests throughout the season, we recorded whether a nest was detected, bird species, stage of nesting, contents, nestling age, and fate. Nest detection, bird species, stage of nesting, contents, nestling age, and fate were recorded after looking through every image containing a nest and selecting the image with the clearest view of the contents of a nest and the highest resolution.
We used the contents of a nest observable through drone transect surveys to determine nest stage. A nest was classified as being in the incubation (I) or nest building (NB) stage until nestlings were visible in an image of a particular nest. A nest was classified as being in the nestling (N) stage once a nestling was observed in a nest, until the nestlings were believed to be at least 14 days old (± two days).
To determine the age of nests, we tracked nests from the initial date they were observed. If a nest was abandoned or had failed before chicks were observed, we marked the initial date the nest was observed as day zero. The number of days between subsequent surveys was then added to its initial age. If a nestling was recorded within a nest, we marked the date of the survey and used this date to back-calculate the age of the nest. Given the resolution of the imagery, we were unable to positively ascribe a nestling to a certain age. Therefore, we marked nestlings as 0 days old during their first observation, and the age of the nest was back-calculated based on incubation periods for each species, as follows: Great Egret (Ardea alba) = 25 days, Snowy Egret (Egretta thula) = 22 days, Tricolored Heron (E. tricolor) = 22 days, Cattle Egret (Bubulcus ibis) = 23 days, White Ibis (Eudocimus albus) = 22 days, and Glossy Ibis (Plegadis falcinellus) = 21 days (Billerman et al. 2020). If the age of a nest in previous surveys was ascribed to a negative number because of back-calculating, we marked the initial date the nest was detected as zero days old. This may have occurred if nestlings were not observed in a nest until they were several days old and/or the initial date a nest was detected and tracked was close to the date the first egg was laid. Past the age of fourteen days, chicks are highly mobile and may leave the nest before researchers can record them (Billerman et al. 2020). Therefore, we considered birds to have fledged at fourteen days ± two days (e.g., Chastant et al. 2018, Frederick et al. 2003). Because researchers do not have to directly approach nests while conducting drone surveys, chicks could reliably be associated with a nest past 14 days and up to 21 days old. If a nest was determined to have reached fledging, we marked the nest as successful and no longer tracked it in surveys.
During the 2021 waterbird breeding season, we followed the fates of 563 different nests (307 Snowy Egret, 64 Great Egret, 83 Tricolor Heron, 24 Cattle Egret, 49 Glossy Ibis, 30 White Ibis, and 6 Little Blue Heron) using ground-based surveys for a total of 2813 individual observations (sum of all observations of all nests; Table 1).
The dataset used for either method differed because each dataset comprised only those nests observed through the associated survey method. During the 2021 waterbird breeding season, we followed the fates of 510 different nests (300 Snowy Egret, 64 Great Egret, 65 Tricolor Heron, 24 Cattle Egret, 25 Glossy Ibis, 28 White Ibis, and 6 Little Blue Heron) using drone nest transect surveys, with a total of 2432 individual observations (sum of all observations of all nests; Table 1).
Statistical analysis
To account for biases resulting from variation in the initial nest ages at monitored nests, we calculated daily nest survival using logistic exposure models (Shaffer 2004). These logistic exposure models use generalized linear models with a bimodal response distribution for the fate of a nest during an interval of time (fate = 1 if the nest survived, fate = 0 if the nest failed), logistic equations, and modifications to the logit link function such that it accounts for variation in the length of an observation period. The model converts probabilities of survival during an observation interval into daily probabilities of survival (daily survival rate: DSR). DSR can be converted to overall survival rate (DSRinterval length) by exponentiating the interval length. The assumptions of this method are that the fates of each nest are independent and that daily survival rate remains consistent within each observation interval (Rotella et al. 2004, Shaffer 2004).
We compared differences in survival between survey methods for all species pooled, each plumage color, and some single species (all species pooled, white-colored birds, dark-colored birds, Great Egret, Snowy Egret, and Tricolored Heron), at each colony surveyed (Clewiston, Little Bear Beach, Eagle Bay, and Moonshine Bay). Only a single focal species of white-colored waterbird, Great Egret, constructed nests above the canopy. Thus, only Great Egret nests were used to compare differences in survival between survey methods for white-colored species that nest above the canopy. The range of differences in the mean overall survival rate between survey methods for all species pooled, each plumage color, and species, among colonies, were presented as the lowest and greatest difference in absolute value of survival estimates between ground-based and drone-based survival estimates. To visually display the difference of survival estimates between survey methods, we paired a box plot of means and 95% confidence intervals of estimates of daily survival rate for each species, survey method (Fig 1.) and colony (Fig 2.). All analyses and plotting were performed in program R (version 4.2.0; R Core Team 2022).
RESULTS
The difference in the overall survival rate between survey methods (drone-based survival estimates minus ground-based survival estimates) for all species pooled, across colonies, was low: 0.59%, 95% CI [0.20, 0.91], but variation was not constant among species and plumage color groups (Fig. 1). The difference in the overall survival rate, between survey methods, across colonies, for white and dark-colored birds, was low, with noticeable variation among species (difference calculated as drone minus ground based method) between survival estimate for white-colored waterbirds (1.41%, 95% CI [9.46, 2.26]), Great Egret (-1.37%, 95% CI [-0.90%, -1.89%]), and Snowy Egret (-0.51%, 95% CI [3.19%, -1.36%; Fig. 1). The difference in the overall survival rate, between survey methods, across colonies, for Tricolored Heron and dark-colored waterbirds, was greater than that observed with white-colored waterbird species, as follows: difference between survival estimate for Tricolored Heron (2.68%, 95% CI [9.46%, 2.26%]), and dark-colored waterbirds (-5.73, 95% CI [3.77%, 7.33%]). The difference in the overall survival rate between survey methods and across colonies, for the white-colored species that nests above the canopy (Great Egret) was lower than any other species or color grouping included in the study.
The range of differences in the mean overall survival rate between survey methods for all species pooled, among colonies, was low (0.38–2.89%, 95% CI [1.07–3.03, 0.02–3.83]), with the variation between plumage color and species differing by colony (Fig. 2, Table 2). The range of differences in the mean overall survival rate between survey methods for white-colored species pooled, among colonies (0.24–8.61%, 95% CI [0.24–8.61, 0.31–10.52]), was greater than the range of differences in the mean overall survival rate between survey methods for Great Egret among colonies (0–1.46%, 95% CI [0–2.04, 0–1.0]) and Snowy Egret among colonies (0–2.49%, 95% CI [0–3.13, 0–2.01]; Fig. 2). The range of differences in the mean overall survival rate between survey methods for dark-colored species pooled, among colonies, was high (5.05–10.29%, 95% CI [2.54–12.06, 4.33–9.17]; Fig. 2). Tricolored Herons displayed a similar range of differences in the mean overall survival rate between survey methods among colonies, with the exception of a single colony (0–10.29%, 95% CI [2.54–12.06, 4.33–9.17]; Fig. 2). The range of differences among colonies in the mean overall survival rate between survey methods for the Great Egret was lower than any other species or color grouping.
DISCUSSION
Although variation between survival estimates of ground-based and drone nest transect surveys was low, the overall survival rate between survey methods for dark-colored waterbirds was greater than for Snowy Egret, Great Egret, and white-colored waterbirds pooled. This pattern of nest survival differences between plumage color groups was similar to patterns of nest detection where the nests of dark-colored species were detected less than those of white-colored species (Frederick et al. 2003, Barr et al. 2018). The discrepancy between drone-derived dark and white-colored waterbird estimates was reportedly the result of the low contrast of dark colored birds under dense canopy cover (Chabot and Bird 2012, Barr et al. 2018). Whereas dark-colored waterbirds may show high contrast to the green emergent vegetation, below the canopy, they may blend into the darker substrate of the lake and be mistaken for shadows. Similarly, it was more difficult to view dark-colored nestlings under high canopy cover than white-colored nestlings because nestlings could be mistaken for shadows from the canopy and/or the attending parent.
Whereas we found relatively low variation between the overall survival estimates of ground-based and drone nest transect surveys for white-colored waterbirds pooled, Great Egret, and Snowy Egret across colonies, there was a greater range of variation observed for white-colored waterbirds among colonies (Fig. 2). This increase in variation was likely due to birds at a single colony (Moonshine Bay) with greater canopy cover than other colonies and reduced detection from a drone.
The use of drones provided a non-invasive means to estimate nest productivity and nest survival. This is notable because the ability to accurately estimate daily and overall nest survival is dependent on the ability to record and reconstruct nest histories (Rotella 2017). This study demonstrated that our methodology coupled with current drone technology provided estimates of nest survival for several colonial-nesting waterbird species with lower variation that estimates derived from traditional survey methods.
The machine characteristics of the model of drone used in this study (DJI Inspire 2) was instrumental in conducting the individual surveys necessary to estimate survival. Waterbird breeding colonies are often located far distances apart and are inaccessible by land (Frederick 1996). The greater flight time and transmission distance of the drone used in the study was necessary to conduct drone transect surveys given the distance between the home base and breeding colony (up to 750 m). Additionally, the use of a multirotor quadcopter, as opposed to a fixed-wing drone, was essential in capturing images with the resolution needed to identify individual nests to species. Previous studies have noted difficulties in identifying nesting birds to species with lower resolution orthomosaic images (Barr et al. 2018). In this study, a pixel resolution of 0.80 cm allowed for the identification of nests to species. Because multirotor drones have the flexibility and maneuverability to fly at lower speeds and altitudes than a fixed-wing models (Siebert and Teizer 2014), they can capture images with greater magnification and overlap. Greater magnification of individual images increases the resolution of the resulting orthomosaic images, whereas greater overlap is necessary to stitch together images during the initial stages of post-processing. Multirotor drones are also generally smaller and lighter than most fixed-wing models, which gives them the capability to take off and land vertically; these features permit greater diversity of launch locations than many fixed-wing drones (Siebert and Teizer 2014), which was necessary to conduct missions on the lake. Additionally, the ability of the camera’s integrated gimbal allowed for the capture of oblique images. The angle of the images taken allowed researchers to see the contents of nests at angles that might otherwise be obstructed by an attending adult.
Our methods provided several advantages compared to previous studies employing the use of drones or traditional manned aerial surveys to examine waterbird reproductive measures. Our ability to visit transects with a greater frequency allowed us to track the fates of individual nests and estimate nest survival with high relative accuracy. Using a multirotor quadcopter drone equipped with a high-resolution camera, as opposed to the fixed-wing drones (e.g., Chabot et al. 2015, Sarda-Palomera 2017), allowed us to fly at the low speeds and altitudes needed to construct orthomosaic images with greater temporal and spatial resolution than has been reported in prior studies (Sarda-Palomera 2017, Lachman et al. 2020). Given the high canopy cover at many of the breeding colonies we monitored and the use of images captured at different angles and times to observe nest contents, high front and side overlap was necessary to construct accurate orthomosaics and observe nest contents. The overlap employed in this study (90% front and side overlap) would likely be unachievable using a fixed-wing drone paired with the same camera, unless flight altitude was increased and pixel resolution decreased. The use of a multirotor drone also provided several advantages over traditional crewed fixed-wing aerial survey methods. The high cost associated with procuring manned fixed-wing aircraft for aerial surveys would have greatly reduced the frequency at which we could conduct surveys throughout the breeding season; significant observer and detection bias associated with crewed aerial surveys would likely have reduced the accuracy and precision of nest count estimates (Frederick et al. 1996, 2003, Rodgers et al. 2005) and, because that methodology has not been proven to reliably identify and track individual nests, we would have been unable to estimate daily nest survival and fledging success.
Additionally, our methods allowed us to observe nests from a greater number of images across a wider degree of angles than previous studies examining waterbird breeding colonies. Rather than tracking nests using a single orthomosaic image (Chabot and Bird 2015, Sandra-Palomera et al. 2017, Barr et al. 2018), we examined every individual image containing the nest that was incorporated into the orthomosaic image. The identification of those images was automated by using the post-processing software (Agisoft Metashape) and creating a reference marker for a particular nest using the function Filter Photos by Marker.
Although the methods employed for this study offered several advantages over previous studies, there was one notable limitation. Unless an unobstructed clear view of a nest and its contents was observed in one of the pictures, the researcher had to manually inspect every picture containing the nest during the associated survey. This often included several dozen individual pictures. Therefore, our study of nest survival took longer than traditional survey methods, whereas the opposite was true for studies using drones to count nests.
Drones are a rapidly advancing technology that can be advantageous over traditional ground-based methods for estimating nest survival for the species under the conditions in our study area and more broadly across avian conservation and research. Drones can be employed to survey large areas at high resolution and can be used to estimate survival throughout the breeding season. This would not be possible with traditional fixed-wing aerial methodology and very difficult to achieve using traditional ground-based methodology given the associated time and cost. This may allow researchers to survey a greater number of breeding colonies and increase the breadth of survival estimates across study areas. The advantages of using drones over other methods depends on the habitat being surveyed, target species, and parameters (e.g., nest counts versus nest survival) that researchers wish to record. If drone surveys are conducted at an appropriate altitude and species nesting traits are considered (Cantu de Leija et al. 2023), drones can disturb birds less than ground-based surveys, adding to their advantage as an emerging tool for avian researchers.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
Funding was provided by the Harte Research Institute for Gulf of Mexico Studies at Texas A&M University—Corpus Christi and Florida Atlantic University. I would like to thank Michelle Petersen for her expertise and guidance during field work. Finally, I would like to thank David Essian, Antonio Cantu de Leija, Alex Sharp, Jacqueline Evans, Lars Nelson, and all the volunteers, technicians, fellow graduate students, and members of the Harte Research Institute’s Conservation and Biodiversity Lab who contributed to this project.
DATA AVAILABILITY
Data and code are available from the author upon request.
LITERATURE CITED
Abd-Elrahman, A., L. Pearlstine, and F. Percival. 2005. Development of pattern recognition algorithm for automatic bird detection from unmanned aerial vehicle imagery. Surveying and Land Information Science 65(1):37-45. https://www.researchgate.net/publication/289617088_Development_of_pattern_recognition_algorithm_for_automatic_bird_detection_from_unmanned_aerial_vehicle_imagery
Afán, I., Máñez, M. and R. Díaz-Delgado. 2018. Drone monitoring of breeding waterbird populations: the case of the glossy ibis. Drones 2(4):42. https://doi.org/10.3390/drones2040042
Agüera-Vega, F., F. Carvajal-Ramírez, and P. Martínez-Carricondo. 2017. Assessment of photogrammetric mapping accuracy based on variation ground control points number using unmanned aerial vehicle. Measurement 98:221-227. https://doi.org/10.1016/j.measurement.2016.12.002
Barr, J. R., M. C. Green, S. J. DeMaso, and T. B. Hardy. 2018. Detectability and visibility biases associated with using a consumer‐grade unmanned aircraft to survey nesting colonial waterbirds. Journal of Field Ornithology 89(3):242-257. https://doi.org/10.1111/jofo.12258
Bevan, E., Whiting, S., T. Tucker, M. Guinea, A. Raith, and R. Douglas. 2018. Measuring behavioral responses of sea turtles, saltwater crocodiles, and crested terns to drone disturbance to define ethical operating thresholds. PLoS ONE 13(3):e0194460. https://doi.org/10.1371/journal.pone.0194460
Billerman, S. M., B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg. 2020. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://birdsoftheworld.org
Boon, M. A., A. P. Drijfhout, and S. Tesfamichael. 2017. Comparison of a fixed-wing and multi-rotor UAV for environmental mapping applications: a case study. International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences XLII-2/W6:47-54. https://doi.org/10.5194/isprs-archives-XLII-2-W6-47-2017
Cantu de Leija, A., R. E. Mirzadi, J. Randall, M. Portmann, E. Mueller, and D. E. Gawlik. 2023. A meta-analysis of disturbance caused by drones on nesting birds. Journal of Field Ornithology 94:3. https://doi.org/10.5751/JFO-00259-940203
Chabot, D., and D. M. Bird. 2012. Evaluation of an off-the-shelf unmanned aircraft system for surveying flocks of geese. Waterbirds 35(1):170-174. https://doi.org/10.1675/063.035.0119
Chabot, D., S. R. Craik, and D. M. Bird. 2015. Population census of a large common tern colony with a small unmanned aircraft. PLoS ONE 10(4):e0122588. https://doi.org/10.1371/journal.pone.0122588
Chastant, J. E., and D. E. Gawlik. 2018. Water level fluctuations influence wading bird prey availability and nesting in a subtropical lake ecosystem. Waterbirds 41(1):35-45. https://doi.org/10.1675/063.041.0105
Cook, M. I., and M. Baranski. 2021. Systemwide summary. South Florida Wading Birding Report 26:1-7. https://www.sfwmd.gov/sites/default/files/documents/SFWBR_2020_Final.pdf
Colomina, I., and P. Molina. 2014. Unmanned aerial systems for photogrammetry and remote sensing: a review. ISPRS Journal of photogrammetry and remote sensing 92:79-97. https://doi.org/10.1016/j.isprsjprs.2014.02.013
David, P. G. 1994. Wading bird use of Lake Okeechobee relative to fluctuating water levels. Wilson Bulletin 106(4):719-732. https://www.jstor.org/stable/4163488
Dinsmore, S. J., G. C. White, and F. L. Knopf. 2002. Advanced techniques for modeling avian nest survival. Ecology 83(12):3476-3488. https://doi.org/10.1890/0012-9658(2002)083[3476:ATFMAN]2.0.CO;2
Essian, D. A., R. Paudel, and D. E. Gawlik. 2022. Predicting effects of water management on breeding abundance of three wading bird species. Journal of Wildlife Management 86(1):e22155. https://doi.org/10.1002/jwmg.22155
Furness, R. W., and P. Monaghan. 1987. Seabird feeding ecology. Pages 23-34 in Seabird ecology. Springer, Boston, Massachusetts, USA. https://doi.org/10.1007/978-1-4613-2093-7_3
Floreano, D., and R. J. Wood. 2015. Science, technology and the future of small autonomous drones. Nature 521(7553):460-466. https://doi.org/10.1038/nature14542
Frederick, P. C., and M. W. Collopy. 1989. Researcher disturbance in colonies of wading birds: effects of frequency of visit and egg-marking on reproductive parameters. Colonial Waterbirds 12:152-157. https://doi.org/10.2307/1521335
Frederick, P. C., B. Hylton, J. A. Heath, and M. Ruane. 2003. Accuracy and variation in estimates of large numbers of birds by individual observers using an aerial survey simulator. Journal of Field Ornithology 74(3):281-287. https://doi.org/10.1648/0273-8570-74.3.281
Frederick, P. C., T. Towles, R. J. Sawicki, and G. T. Bancroft. 1996. Comparison of aerial and ground techniques for discovery and census of wading bird (Ciconiiformes) nesting colonies. Condor: Ornithological Applications 98(4):837-841. https://doi.org/10.2307/1369865
Garg, P. K. 2022. Characterisation of fixed-wing versus multirotors uavs/drones. Journal of Geomatics 16(2):152-159. https://doi.org/10.58825/jog.2022.16.2.44
Hodgson, J. C., S. M. Baylis, R. Mott, A. Herrod, and R. H. Clarke. 2016. Precision wildlife monitoring using unmanned aerial vehicles. Scientific Reports 6(1):22574. https://doi.org/10.1038/srep22574
Hodgson, J. C., R. Mott, S. M. Baylis, T. T. Pham, S. Wotherspoon, A. D. Kilpatrick, R. Raja Segaran, I. Reid, A. Terauds, and L. P. Koh. 2018. Drones count wildlife more accurately and precisely than humans. Methods in Ecology and Evolution 9(5):1160-1167. https://doi.org/10.1111/2041-210X.12974
Jehle, G., A. A. Yackel Adams, J. A. Savidge, and S. K. Skagen. 2004. Nest survival estimation: a review of alternatives to the Mayfield estimator. Condor: Ornithological Applications 106(3):472-484. https://doi.org/10.1093/condor/106.3.472
Johnson, K. G., M. S. Allen, and K. E. Havens. 2007. A review of littoral vegetation, fisheries, and wildlife responses to hydrologic variation at Lake Okeechobee. Wetlands 27(1):110-126. https://doi.org/10.1672/0277-5212(2007)27[110:AROLVF]2.0.CO;2
Jones, G. P., L. G. Pearlstine, and H. F. Percival. 2006. An assessment of small unmanned aerial vehicles for wildlife research. Wildlife Society Bulletin 34(3):750-758. https://doi.org/10.2193/0091-7648(2006)34[750:AAOSUA]2.0.CO;2
Lachman, D., C. Conway, K. Vierling, and T. Matthews. 2020. Drones provide a better method to find nests and estimate nest survival for colonial waterbirds: a demonstration with Western Grebes. Wetlands Ecology and Management 28(5):837-845. https://doi.org/10.1007/s11273-020-09743-y
Lyons, M., K. Brandis, C. Callaghan, J. McCann, C. Mills, S. Ryall, and R. Kingsford. 2018. Bird interactions with drones, from individuals to large colonies. Australian Field Ornithology 35:51-56. https://afo.birdlife.org.au/afo/index.php/afo/article/view/2109
McEvoy, J. F., G. P. Hall, and P. G. McDonald. 2016. Evaluation of unmanned aerial vehicle shape, flight path and camera type for waterfowl surveys: disturbance effects and species recognition. PeerJ 4:e1831. https://doi.org/10.7717/peerj.1831
Orange, J. P., R. R. Bielefeld, W. A. Cox, and A. L. Sylvia. 2023. Impacts of drone flight altitude on behaviors and species identification of marsh birds in Florida. Drones 7(9):584. https://doi.org/10.3390/drones7090584
Parsons, K. C., and T. L. Master. 2020. Snowy Egret (Egretta thula), version 1.0. In A. F. Poole and F. B. Gill, editors. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.snoegr.01
R Core Team. 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rodgers Jr, J. A., P. S. Kubilis, and S. A. Nesbitt. 2005. Accuracy of aerial surveys of waterbird colonies. Waterbirds 28(2):230-237. https://doi.org/10.1675/1524-4695(2005)028[0230:AOASOW]2.0.CO;2
Rotella, J. 2017. Birds of Montana. Condor: Ornithological Applications 119(1):172-173. https://doi.org/10.1650/CONDOR-16-174.1
Rotella, J. J., S. J. Dinsmore, and T. L. Shaffer. 2004. Modeling nest-survival data: a comparison of recently developed methods that can be implemented in MARK and SAS. Animal biodiversity and Conservation 27(1):187-205. https://doi.org/10.32800/abc.2004.27.0187
Sasse, D. B. 2003. Job-related mortality of wildlife workers in the United States, 1937-2000. Wildlife Society Bulletin 31(4):1015-1020. https://www.jstor.org/stable/3784446
Sardà‐Palomera, F., G. Bota, N. Padilla, L. Brotons, and F. Sardà. 2017. Unmanned aircraft systems to unravel spatial and temporal factors affecting dynamics of colony formation and nesting success in birds. Journal of Avian Biology 48(9):1273-1280. https://doi.org/10.1111/jav.01535
Sardà‐Palomera, F., G. Bota, C. Viñolo, O. Pallarés, V. Sazatornil, L. Brotons, S. Gomáriz, and F. Sarda. 2012. Fine‐scale bird monitoring from light unmanned aircraft systems. Ibis 154(1):177-183. https://doi.org/10.1111/j.1474-919X.2011.01177.x
Scarton, F., and R. Valle. 2020. Drone assessment of habitat selection and breeding success of Gull-billed Tern Gelochelidon nilotica nesting on low-accessibility sites: a case study. Rivista Italiana di Ornitologia 90(2):69-76. https://doi.org/10.4081/rio.2020.475
Shaffer, T. L. 2004. A unified approach to analyzing nest success. Auk 121(2):526-540. https://doi.org/10.1642/0004-8038(2004)121[0526:AUATAN]2.0.CO;2
Siebert, S., and J. Teizer. 2014. Mobile 3D mapping for surveying earthwork projects using an Unmanned Aerial Vehicle (UAV) system. Automation in Construction 41:1-14. https://doi.org/10.1016/j.autcon.2014.01.004
South Florida Water Management District. 2021. South Florida wading bird report 2020. West Palm Beach, Florida: South Florida Water Management District. https://www.sfwmd.gov/sites/default/files/documents/SFWBR_2019.pdf
Thamm, F. P., N. Brieger, K. P. Neitzke, M. Meyer, R. Jansen, and M. Mönninghof. 2015. Songbird—an innovative UAS combining the advantages of fixed wing and multi rotor UAS. International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences 40:345-349. https://doi.org/10.5194/isprsarchives-XL-1-W4-345-2015
Tremblay, J., and L. N. Ellison. 1979. Effects of human disturbance on breeding of Black-crowned Night Herons. Auk 96(2):364-369. https://doi.org/10.1093/auk/96.2.364
Vas, E., A. Lescroël, O. Duriez, G. Boguszewski, and D. Grémillet. 2015. Approaching birds with drones: first experiments and ethical guidelines. Biology Letters 11(2):20140754. https://doi.org/10.1098/rsbl.2014.0754
Vermeulen, C., P. Lejeune, J. Lisein, P. Sawadogo, and P. Bouché. 2013. Unmanned aerial survey of elephants. PLoS ONE 8(2):e54700. https://doi.org/10.1371/journal.pone.0054700
Werschkul, D. F., E. McMahon, and M. Leitschuh. 1976. Some effects of human activities on the Great Blue Heron in Oregon. Wilson Bulletin 88(4):660-662. https://digitalcommons.usf.edu/wilson_bulletin/vol88/iss4/20
Williams, K. A., P. C. Frederick, P. S. Kubilis, and J. C. Simon. 2008. Bias in aerial estimates of the number of nests in White Ibis and Great Egret colonies. Journal of Field Ornithology 79(4):438-447. https://doi.org/10.1111/j.1557-9263.2008.00197.x
Fig. 1
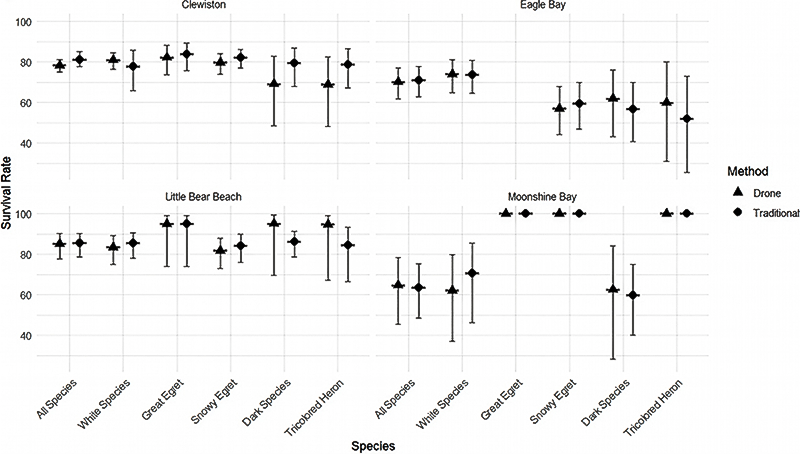
Fig. 1. Overall survival rate (± 95% CI) for traditional survey methods and drone survey methods of all species (Great Egret [Ardea alba], Snowy Egret [Egretta thula], White Ibis [Eudocimus albus], Tricolored Heron [Egretta tricolor], Glossy Ibis [Plegadis falcinellus], and Little Blue Heron [Egretta caerulea]), pooled; white-colored species (Great Egret, Snowy Egret, White Ibis), pooled; Great Egret; Snowy Egret; dark-colored species (Tricolored Heron, Glossy Ibis, Little Blue Heron), pooled; and Tricolored Heron at each colony surveyed (Clewiston, Eagle Bay, Little Bear Beach, and Moonshine Bay).
Fig. 2
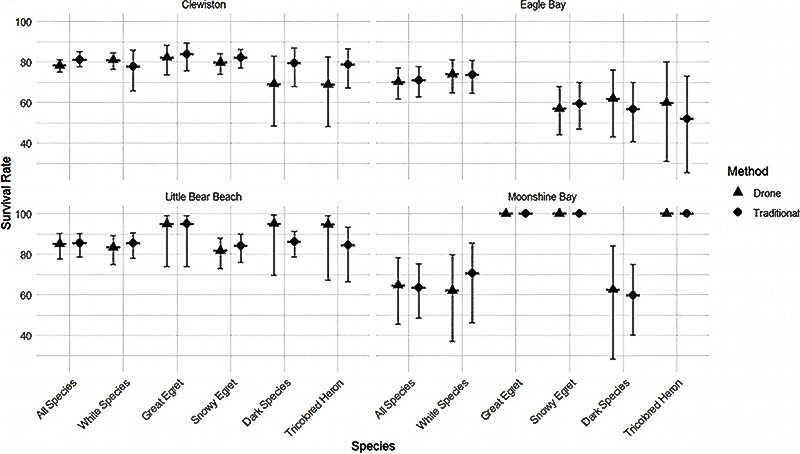
Fig. 2. Daily survival rate (± 95% CI) for traditional survey methods and drone survey methods of all species (Great Egret [Ardea alba], Snowy Egret [Egretta thula], White Ibis [Eudocimus albus], Tricolored Heron [Egretta tricolor], Glossy Ibis [Plegadis falcinellus], and Little Blue Heron [Egretta caerulea]), pooled; white-colored species (Great Egret, Snowy Egret, and White Ibis), pooled; Great Egret; Snowy Egret; and dark-colored species (Tricolored Heron, Glossy Ibis, and Little Blue Heron), pooled; and Tricolored Heron at each colony surveyed (Clewiston, Eagle Bay, Little Bear Beach, and Moonshine Bay).
Table 1
Table 1. Total number of nests observed (Nests), total number of observations of all nests (Total observations), and mean detections per nest (Mean detections per nest) for drone surveys and ground-based surveys for all species, pooled (All species pooled); white-colored species (Great Egret [Ardea alba], Snowy Egret [Egretta thula], Cattle Egret [Ardea ibis], and White Ibis [Eudocimus albus]), pooled (White species pooled); dark-colored species (Tricolored Heron [Egretta tricolor], Glossy Ibis [Plegadis falcinellus], and Little Blue Heron [Egretta caerulea]), pooled (Dark species pooled); Great Egret; Snowy Egret; Cattle Egret; White Ibis; Tricolored Heron; Glossy Ibis; and Little Blue Heron. SD denotes the standard deviation for mean detections per nest.
| Grouping | Method | Nests | Total observations | Mean detections per nest | SD | ||||
| All species pooled | Drone | 512 | 2432 | 4.8 | 2.43 | ||||
| Traditional | 563 | 2813 | 5.0 | 2.43 | |||||
| White species pooled | Drone | 416 | 2080 | 5.0 | 2.43 | ||||
| Traditional | 425 | 2224 | 5.2 | 2.4 | |||||
| Dark species pooled | Drone | 96 | 352 | 3.7 | 2.17 | ||||
| Traditional | 138 | 582 | 4.2 | 2.39 | |||||
| Great Egret | Drone | 64 | 389 | 6.1 | 2.63 | ||||
| Traditional | 64 | 390 | 6.1 | 2.65 | |||||
| Snowy Egret | Drone | 300 | 1482 | 4.9 | 2.35 | ||||
| Traditional | 307 | 1613 | 5.3 | 2.32 | |||||
| Cattle Egret | Drone | 24 | 140 | 5.8 | 1.67 | ||||
| Traditional | 24 | 141 | 5.9 | 1.62 | |||||
| White Ibis | Drone | 28 | 68 | 2.4 | 0.74 | ||||
| Traditional | 30 | 79 | 2.6 | 1.03 | |||||
| Tricolored Heron | Drone | 65 | 249 | 3.8 | 2.2 | ||||
| Traditional | 83 | 407 | 4.9 | 2.49 | |||||
| Glossy Ibis | Drone | 25 | 76 | 3.0 | 2.16 | ||||
| Traditional | 49 | 148 | 3.0 | 1.81 | |||||
| Little Blue Heron | Drone | 6 | 28 | 4.7 | 1.37 | ||||
| Traditional | 6 | 28 | 4.7 | 1.37 | |||||
Table 2
Table 2. Overall survival rate (Overall survival %), lower 95% CI (LWR 95% CI), and upper 95% CI (UPR 95% CI) for traditional survey methods and drone survey methods (Method) of all species (Great Egret [Ardea alba], Snowy Egret [Egretta thula], White Ibis [Eudocimus albus], Tricolored Heron [Egretta tricolor], Glossy Ibis [Plegadis falcinellus], and Little Blue Heron [Egretta caerulea]), pooled; white-colored species (Great Egret, Snowy Egret, and White Ibis), pooled; Great Egret; Snowy Egret; dark-colored species (Tricolored Heron, Glossy Ibis, and Little Blue Heron), pooled; and Tricolored Heron (species) at each colony surveyed (Clewiston, Eagle Bay, Little Bear Beach, and Moonshine Bay).
| Colony | Species | Method | Overall survival % | LWR 95% CI | UPR 95% CI | ||||
| Clewiston | White species | Drone | 80.9 | 76.41 | 84.62 | ||||
| Clewiston | White species | Traditional | 77.79 | 65.89 | 85.97 | ||||
| Eagle Bay | White species | Drone | 73.91 | 64.88 | 80.97 | ||||
| Eagle Bay | White species | Traditional | 73.67 | 64.57 | 80.79 | ||||
| Little Bear Beach | White species | Drone | 83.43 | 75.18 | 89.14 | ||||
| Little Bear Beach | White species | Traditional | 85.62 | 78.07 | 90.74 | ||||
| Moonshine Bay | White species | Drone | 62.07 | 37.11 | 79.6 | ||||
| Moonshine Bay | White species | Traditional | 70.68 | 46.18 | 85.64 | ||||
| Clewiston | Great Egret | Drone | 82.21 | 73.53 | 88.28 | ||||
| Clewiston | Great Egret | Traditional | 83.67 | 75.57 | 89.28 | ||||
| Little Bear Beach | Great Egret | Drone | 95.02 | 74.04 | 99.03 | ||||
| Little Bear Beach | Great Egret | Traditional | 95.02 | 74.04 | 99.03 | ||||
| Moonshine Bay | Great Egret | Drone | 100 | 100 | 100 | ||||
| Moonshine Bay | Great Egret | Traditional | 100 | 100 | 100 | ||||
| Clewiston | Snowy Egret | Drone | 79.65 | 73.99 | 84.22 | ||||
| Clewiston | Snowy Egret | Traditional | 82.14 | 77.01 | 86.23 | ||||
| Eagle Bay | Snowy Egret | Drone | 57.04 | 44.26 | 67.98 | ||||
| Eagle Bay | Snowy Egret | Traditional | 59.26 | 46.73 | 69.8 | ||||
| Little Bear Beach | Snowy Egret | Drone | 81.83 | 72.91 | 88.05 | ||||
| Little Bear Beach | Snowy Egret | Traditional | 84.22 | 76.04 | 89.81 | ||||
| Moonshine Bay | Snowy Egret | Drone | 100 | 100 | 100 | ||||
| Moonshine Bay | Snowy Egret | Traditional | 100 | 100 | 100 | ||||
| Clewiston | Snowy Egret | Drone | 79.65 | 73.99 | 84.22 | ||||
| Clewiston | Snowy Egret | Traditional | 82.14 | 77.01 | 86.23 | ||||
| Eagle Bay | Snowy Egret | Drone | 57.04 | 44.26 | 67.98 | ||||
| Eagle Bay | Snowy Egret | Traditional | 59.26 | 46.73 | 69.8 | ||||
| Little Bear Beach | Snowy Egret | Drone | 81.83 | 72.91 | 88.05 | ||||
| Little Bear Beach | Snowy Egret | Traditional | 84.22 | 76.04 | 89.81 | ||||
| Moonshine Bay | Snowy Egret | Drone | 100 | 100 | 100 | ||||
| Moonshine Bay | Snowy Egret | Traditional | 100 | 100 | 100 | ||||
| Clewiston | Tricolored Heron | Drone | 68.79 | 48.41 | 82.51 | ||||
| Clewiston | Tricolored Heron | Traditional | 78.63 | 67.1 | 86.55 | ||||
| Eagle Bay | Tricolored Heron | Drone | 59.87 | 30.83 | 80.11 | ||||
| Eagle Bay | Tricolored Heron | Traditional | 51.81 | 25.69 | 72.94 | ||||
| Little Bear Beach | Tricolored Heron | Drone | 94.62 | 67.32 | 99.24 | ||||
| Little Bear Beach | Tricolored Heron | Traditional | 84.56 | 66.58 | 93.35 | ||||
| Moonshine Bay | Tricolored Heron | Drone | 100 | 100 | 100 | ||||
| Moonshine Bay | Tricolored Heron | Traditional | 100 | 100 | 100 | ||||
| Clewiston | Dark species | Drone | 69.02 | 48.75 | 82.65 | ||||
| Clewiston | Dark species | Traditional | 79.31 | 68.05 | 86.98 | ||||
| Eagle Bay | Dark species | Drone | 61.9 | 43.26 | 76.07 | ||||
| Eagle Bay | Dark species | Traditional | 56.85 | 40.72 | 70.02 | ||||
| Little Bear Beach | Dark species | Drone | 95.08 | 69.69 | 99.3 | ||||
| Little Bear Beach | Dark species | Traditional | 86.2 | 78.91 | 91.12 | ||||
| Moonshine Bay | Dark species | Drone | 62.56 | 28.24 | 84.2 | ||||
| Moonshine Bay | Dark species | Traditional | 59.93 | 40.3 | 75.03 | ||||