







The following is the established format for referencing this article:
Lou, Y., K. Song, Y. Hong, Y. Fang, M. Wang, H. Lloyd, and Y.-H. Sun. 2026. Seasonal differences in dietary compositions of Blood Pheasant (Ithaginis cruentus): a study focuses on bryophytes from feces analysis. Journal of Field Ornithology 97(2):10.ABSTRACT
The Blood Pheasant (Ithaginis cruentus) may be unique amongst Galliformes species in that it is believed to forage primarily on bryophytes. Yet there are few quantitative studies that have examined the species diet and how this might vary seasonally, as these data could shed some light on how such species are able to adapt to harsher, cold montane environments. To help fill this knowledge gap, we used microhistological fecal analysis to determine the dietary composition of a Blood Pheasant population at Lianhuashan Nature Reserve in Gansu Province, China. A total of 149 fresh feces samples of Blood Pheasant were collected continuously from April 2011 to October 2013 (spanning all four seasons) along 18 km of roads in the reserve. Bryophytes were the most abundant items in the fecal samples (99.8%), of which 11 families were recognized. Mniaceae was present in nearly all fecal samples and constituted the main dietary component across the population. There was significant seasonal variation in bryophyte presence in fecal samples, with the proportion of bryophytes lowest in summer, and highest during autumn. In addition, the digestibility of bryophytes was also highest in autumn compared to other seasons. Our results highlight a degree of dietary plasticity by Blood Pheasants and we discuss how this variation enables them to adapt to local seasonal climatic conditions in their montane environment.
RESUMEN
El Faisán Sangriento (Ithaginis cruentus) puede ser único entre las especies de Galliformes en el hecho de que forrajea primariamente de briófitos. Sin embargo hay pocos estudios cuantitativos que han examinado la dieta de la especie y cómo ésta puede variar estacionalmente, ya que estos datos podrían dilucidar un poco cómo tal especie se adapta a ambientes montanos más duros y fríos. Para ayudar a llenar este vacío de conocimiento, usamos análisis microhistológico fecal para determinar la composición dietética de una población de Faisán Sangriento en la Reserva Natural Lianhuashan en la Provincia Gansu, China. Un total de 149 muestras de heces de Faisán Sangriento fueron colectadas continuamente desde Abril 2011 hasta Octubre 2013 (cubriendo las 4 estaciones) a lo largo de carreteras de 18 km en la reserva. Los Briófitos fueron los ítems más abundantes en las muestras fecales (99.8%), de las cuales 11 familias fueron reconocidas. Mniaceae estuvo presente en casi todas las muestras fecales y constituyó el componente dietético principal en la población. Hubo una variación estacional significativa en la presencia de briófitos en las muestras fecales, con la menor proporción en verano y la mayor durante otoño. Además, la digestabilidad de los briófitos fue la más alta en otoño comparada con otras estaciones. Nuestros resultados resaltan un nivel de plasticidad dietética en el Faisán Sangriento y discutimos cómo esta variación les permite adaptarse a condiciones climáticas estacionales locales en su ambiente montano.
INTRODUCTION
Dietary research on animal species is crucial to further our understanding of animal ecology and evolution (Putman 1984, Sutherland et al. 2004). Diet is associated with many life history strategies of animals, such as behavioral (Levin et al. 2009) and energy intake strategies (Sih and Christensen 2001). Studies on dietary compositions can provide information on resource utilization, food abundance, and seasonal variation in resources available for a given animal species (Hammerschlag et al. 2010), and also shed some light on patterns of habitat selection, which is crucial for animal conservation (Putman 1984, Baubet et al. 2004).
For animals in general, there is a trade-off between food abundance, handling time, nutritional value, and digestibility of food resources (Bjørkvoll et al. 2009). When high-quality food resources are abundant, individuals are expected to increase their foraging activities on these food types (Emlen 1966). Among the food types of animals, bryophytes are a kind of special food resource. Several studies have revealed that bryophytes contain protein, carbohydrate, and lipid compounds (Prins 1982), and they are known to have low nutrient and caloric values (Emlen 1966). Furthermore, bryophytes contain high concentration of lignin-like compounds (Popper 2008), which hinder access to the more digestible cell content, reducing their digestibility, and resulting in very low metabolized energy content (Prins 1982). Therefore, only a few bird species forage mainly on bryophytes (Prins 1982, Glime 2017), especially in the cold environment (Russo et al. 2020). For example, in sub-Antarctic South America, White-bellied Seedsnipe (Attagis malouinus) and Upland Geese (Chloephaga picta) consume bryophytes frequently (Lázaro et al. 2021).
Given temporal and seasonal changes in food quality (e.g., nutrient content) and quantity, animals need to adjust their dietary composition accordingly (Blanco-Fontao et al. 2010, Whelan et al. 2015, Bravo et al. 2016). For example, White-eared Hummingbird (Basilinna leucotis) forage for arthropods in a higher proportion during the warm-dry season when floral nectar is scarce (Maya-García et al. 2024). Black Grouse (Lyrurus tetrix) are known to favor fruits, flowers, and seeds in summer but consume predominantly leaves during the winter period (Beeston et al. 2005). To our knowledge at least, there are few studies that have quantified the prevalence and seasonal variation of bryophytes in the diets of high-elevation montane vertebrate species.
The Blood Pheasant (Ithaginis cruentus) is a medium-sized montane Galliforme species, that inhabits coniferous and mixed forests at altitudes of 2100–4600 m above sea level. A limited number of studies (Shi and Li 1985, Yao 1992, Wang et al. 2011) have revealed that bryophytes and leaves form part of the species’ diets in winter and spring (Shi and Li 1985), whilst leaves, berries, invertebrates, and bryophytes form part of their summer and autumn diet (Li and Li 1981). Thus, Blood Pheasant is a suitable species to examine variation in the prevalence and variation of bryophytes in their diet. Yet such studies are difficult to undertake given the furtive behavior of the species and the difficulty of distinguishing different food items by observations alone (Wang et al. 2011). Here, we investigate seasonal variation in the dietary composition of a wild Blood Pheasant population, in the Lianhuashan Nature Reserve in Gansu province, China. Given that, in our study area, Blood Pheasant is very sensitive to human disturbance, and the difficulties associated with making direct observations of the species’ feeding behavior, we used microhistological fecal method to quantify the species diet (Holechek et al. 1982), which depends on the resistance of digestion. Our objectives of this study are three-fold: (1) to describe the dietary compositions of the Blood Pheasant, (2) and dietary compositions of bryophytes, (3) to determine whether the proportion and the digestibility of bryophytes varied seasonally.
METHODS
Study area
The study was conducted at the Shahetan station in the core area of the Lianhuashan Natural Reserve, southern Gansu Province, China. The elevation of the Shahetan station is 2864 m, and mean temperature of the whole year ranges from 3.2 to 4.6 °C. There are four major types of vegetation: (1) conifer forests, dominated by spruces (Picea asperata) and firs (Abies fabri); (2) deciduous forests, dominated by birches (Betula platyphylla) and willows (Salix spp.); (3) shrubs, containing willows, roses (Rosa L.), and honeysuckles (Lonicera japonica); (4) ground vegetation, dominated by moss (Bryophyte spp.), grass (Gramineae spp.), and berries (Fragaria orientalis). Detailed information on the vegetation found in the region can be found in Sun et al. (2003). According to the local climate conditions, we defined spring as occurring between April to May, summer occurs between June to August, September to October corresponded to autumn, and November to March corresponded to winter.
Fresh feces of Blood Pheasant were collected from 18 km of the main roads (the width ranges from 1 to 3 m) of our study area over four seasons. Blood Pheasant feces are easily recognizable in the wild because the fecal shape is very different to those of other sympatry Phasianidae species (Blue eared pheasant, Crossoptilon auritum, and Chinese grouse, Tetrastes sewerzowi) also found in the study area; the shape of Blue Eared Pheasant feces is the largest, the Blood Pheasant is the medium, and the Chinese Grouse is the smallest. Furthermore, the feces of Blood Pheasant typically contain visible quantities of undigested moss and grass. We collected fresh feces during four seasons, after which, we placed them in a ventilated place until they reached a constant weight and stored them in valve bags for subsequent laboratory analysis. In total, 149 fecal samples were collected at Lianhuashan Natural Reserve from April 2011 to October 2013, including 48 samples during spring, 23 during summer, 41 during autumn, and 37 during winter.
Each fecal sample was dispersed and mixed in water prior to analysis. We first took 30 μL from the mixture, and applied it to microscope slides, each of which was systematically examined at 100X magnification under a microscope (OLYMPUS CX21). Because of the high digestion of fruits, which could not be found in the feces, we only classified food items into bryophytes and spermatophytes according to the observed cell structure: bryophytes were recognizable because of their monolayer cell structure, whereas spermatophytes were recognizable from their multilayered cell structure. We estimated the percentage of different plant types in each sample while systematically counting each plant type in each sample, to avoid double counting. Because of the difficulties in identifying spermatophytes by using a microscope (Chapuis 1979, Atsalis 1999, Scheiffarth 2001), only bryophytes were identified to families from their distinctive morphological features retained in the fecal samples (Iwatsuki and Mizutani 1981). To identify the family of bryophytes, each sample was viewed under a microscope at 400X magnification (Iwatsuki and Mizutani 1981). The cells were identified by comparison with a reference collection of bryophytes from Lianhuashan Nature Reserve (unpublished data).
We also estimated the percentage of Mniaceae in the bryophytes in each sample, and based on the digestion of border cells (Fig. 1), we were able to classify the digestibility of bryophytes into four levels: first level, only the outermost cells of bryophytes were digested, which represented the lowest digestion condition. Second level, where digestion of the bryophytes included marginal border cells. For the third level, the extent of digestion included marginal border cells and partial mesophyll cells. For the final fourth level, which we considered the highest digestion condition, bryophytes were digested to the extent that only a few mesophyll cells and veins were visible, which was the highest digestion condition. To reduce the variation estimated by different observers, all identifications and estimations were done by the same person (M. Wang).
Data analysis
Kruskal-wallis tests were used to investigate seasonal differences in the consumption of bryophytes, spermatophyte, Mniaceae, and digestibility of bryophytes. The Dunn tests were used to investigate the differences between each two seasons. Spearman rank correlation was used to examine the relationship between proportion of bryophytes and Mniaceae in fecal samples. Diversity of bryophytes families from samples collected in each season was estimated using the Shannon-Wiener index. Kruskal-wallis test was used to investigate the differences of Shannon-Wiener index among seasons.
RESULTS
Diet composition of Blood Pheasant at Lianhuashan
We found that the bryophytes and spermatophytes were more frequently consumed and they presented in 99.80% and 93.36% of Blood Pheasant feces, respectively. The mean percentage of bryophytes and spermatophyte was 54.16% and 44.84%, respectively, in total feces samples. Blood Pheasant consumed 11 families of bryophytes, including Mniaceae, Thuidiacea, Hypnaceae, Brachytheciaceae, Entodontaceae, Pottiaceae, Bryaceae, Philonotis, Grimmiaceae, Sematophyllaceae, Meteoriacea (see Table 1). Furthermore, Mniaceae was the most frequently occurring bryophyte family in the samples (97.14%), and the proportion of Mniaceae intake was positively correlated with the proportion of moss intake (rs = 0.803, P < 0.001). Three families of bryophytes, Philonotis, Grimmiaceae, and Meteoriaceae, along with capsules of bryophytes, were only recorded on one occasion, and all were recorded from autumn fecal samples.
Seasonal variation of diet composition and diversity
There were significant differences in Blood Pheasant diet among seasons, both in bryophytes and spermatophytes (H = 11.630, P = 0.009; H = 11.630, P = 0.009, respectively, Fig. 2A). The proportion of bryophytes was higher in spring, autumn, and winter than summer (Table 2), while the proportion of spermatophytes was higher in summer than other three seasons (Table 2). The percentage of Mniaceae in the fecal samples was significantly different among seasons (H = 23.689, P < 0.001, Fig. 2B) and it was higher in autumn than the other three seasons (Table 3). The digestibility of bryophytes also exhibited significant seasonal variation (H = 84.311, P < 0.001, Fig. 2C), and it was higher in autumn than the other three seasons (Table 4). Diversity of bryophytes in fecal samples was fairly similar across all seasons (H = 1.663 spring; H = 1.389 summer; H = 1.794 autumn; H = 1.817 winter), and there was no significant difference among seasons (H = 3, P = 0.392).
DISCUSSION
In our study population, the dietary composition of Blood Pheasant contained 11 families of bryophytes, with the family Mniaceae being the dominant bryophyte family. In addition, the proportion of bryophytes, Mniaceae, and the digestibility of bryophytes in Blood Pheasant diet varied significantly among seasons.
Based on the occurrence and percentage of bryophytes in the fecal samples, we can conclude that bryophytes constitute a significant dietary component of Blood Pheasant population at Lianhuashan, as suggested by Wang et al. (2011). Prins (1982) suggested that animals tend to forage on bryophytes in colder, harsher environments because these bryophyte families are known to contain relatively high concentrations of arachidonic acid, and this may explain, in part at least, the prevalence of bryophytes in our Blood Pheasant fecal samples.
The families of bryophytes in our study site were different to previous studies (Shi and Li 1985, Yao 1992, Wang et al. 2011). For example, Wang et al. (2011) found that Blood Pheasant also foraged on bryophytes from 16 families at their study site at the Bangpo Monastery, such as Scorpidiaceae, Funariaceae, and Hedwigiaceae. One explanation is the noticeable differences in the environments among study sites, and the availability and abundance of bryophytes may be different.
Compared to the other 10 bryophyte families found within Blood Pheasant fecal samples, the Mniaceae may have three advantages: first, species of Mniaceae tend to have much bigger leaf cells that contain more water and nutrients (Iwatsuki and Mizutani 1981); second, the soft plastids of Mniaceae are easier to digest, because we found that the digested parts of Mniaceae bryophytes comprised only leafy gametophyte, without the rib of the leaf, capsule, or Poaceae plant, as was found by Wang et al. (2011). These authors suggest that Blood Pheasant preferred bryophytes that are softer in texture, thus perhaps limiting Blood Pheasant to forage predominantly on Mniaceae at Lianhuashan; third, a relative higher content of arachidonic acid, which could help individuals adapt to the cold (Hansen and Rossi 1999, Pejin et al. 2012, Beike et al. 2014). Arachidonic acid could help to maintain an animal’s physical mobility (Turner 1979), enzyme activity (Tallima and El Ridi 2018), and prevent blood clots (Hoak 1997). Another recent study has found similar results, in that Blood Pheasant feeds predominantly on bryophytes with higher content of arachidonic acid (Guo 2020), and that the high content of arachidonic acid is positively associated with better resistance to colder environments in this species (Guo 2020). Furthermore, this result may also help to explain why female Blood Pheasant typically leave their nests only on one occasion each day during the incubation period, averaging 6.6 hours per day away from the nest (Jia et al. 2010). Bryophytes have a lower nutrient value when compared to other food items (Wielgolaski et al. 1975), thus individuals may spend more time foraging for moss compared to other food resources, and would also necessitate spending much time searching and foraging preferentially for Mniaceae species.
There was a significant seasonal variation in the composition in Blood Pheasant fecal samples in our study, although not in diversity of bryophytes families. These patterns could reflect the abundance of specific and/or favorite food resources at Lianhuashan (Wingard et al. 2011). The peak in bryophyte occurrence in autumn fecal samples is noticeable because Blood Pheasants will need to build significant energy resources in preparation for the winter period. The proportion of bryophytes in their diet in summer was significantly lower compared to the other seasons, and may be a result of Blood Pheasants preferring fruits (Jia and Sun 2008), such as strawberry (Fragaria orientalis), which typically have higher nutrient levels, and tend to be more highly digestible and palatable (Nam et al. 2006). Foraging on these foods results in a greater energy intake over a shorter time period (Shi and Li 1985). In addition, during early spring and winter, large areas of the study site were covered with snow and this may have had some effects on the abundance and availability of bryophytes in the study area. The presence of snow in winter and early spring, could drive individuals to explore larger areas or range to lower elevation areas for food (Luo et al. 2016). Thus, further research is needed across a longer period of time to fully examine inter-annual variation in bryophyte abundance and availability, and Blood Pheasant diet in relation to weather conditions (snowfall, precipitation, ambient temperature fluctuations, etc.), and movement patterns because these all represent possible drivers of seasonal variation in dietary composition (Lou et al. 2014, Luo et al. 2016).
Bryophyte digestibility also varied seasonally in our study population. This can be explained by three reasons: first, Blood Pheasant prefer to feed on more Mniaceae species during autumn, which are much more digestible in comparison to other families; second, the patterns of digestibility might reflect the impact of seasonal changes on water content, fiber, and chemical composition of bryophytes (Dilks and Proctor 1979, Reeves 1987); and third, the physiological metabolism rates of different individual pheasants will also vary among seasons (Hoffman et al. 2001, Al-Mansour 2004), which may then affect the levels of digestibility.
Conclusions
We described both the qualitative and quantitative seasonal changes in the diet of Blood Pheasant at Lianhuashan Nature Reserve. These data fill a significant knowledge gap regarding montane Galliforme species’ dietary ecology, highlighting further avenues for ecological research for these species and their food resources, but also offering some insight as to how other vertebrates are able to adapt to seasonal variations in food availability and abundance in these topographically complex, colder montane environments.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
This research was supported by the National Natural Science Foundation of China, China (Project 32070452, 32401287). We are grateful to the staff of the Lianhuashan Nature Reserve, Gansu Province.
DATA AVAILABILITY
Data and code are available from the corresponding author upon request.
LITERATURE CITED
Al-Mansour, M. I. 2004. Seasonal variation in basal metabolic rate and body composition within individual sanderling bird Calidris alba. Journal of Biological Sciences 4:564-567. https://doi.org/10.3923/jbs.2004.564.567
Atsalis, S. 1999. Diet of the brown mouse lemur (Microcebus rufus) in Ranomafana National Park, Madagascar. International Journal of Primatology 20:193-229. https://doi.org/10.1023/A:1020518419038
Baubet, E., C. Bonenfant, S. Brandt. 2004. Diet of the wild boar in the French Alps. Galemys 16:101-113. https://doi.org/10.7325/Galemys.2004.NE.A8
Beeston, R., D. Baines, and M. Richardson. 2005. Seasonal and between-sex differences in the diet of Black Grouse Tetrao tetrix. Bird Study 52:276-281. https://doi.org/10.1080/00063650509461400
Beike, A. K., C. Jaeger, F. Zink, E. L. Decker, and R. Reski. 2014. High contents of very long-chain polyunsaturated fatty acids in different moss species. Plant Cell Reports 33:245-254. https://doi.org/10.1007/s00299-013-1525-z
Bjørkvoll, E., B. Pedersen, H. Hytteborn, S. J. Ingibjörg, and L. Rolf. 2009. Seasonal and interannual dietary variation during winter in female Svalbard reindeer (Rangifer tarandus platyrhynchus). Arctic, Antarctic, and Alpine Research 41:88-96. https://doi.org/10.1657/1523-0430-41.1.88
Blanco-Fontao, B., A. Fernández-Gil, J. R. Obeso, and M. Quevedo. 2010. Diet and habitat selection in Cantabrian Capercaillie (Tetrao urogallus cantabricus): ecological differentiation of a rear-edge population. Journal of Ornithology 151: 269-277. https://doi.org/10.1007/s10336-009-0452-9
Bravo, C., C. Ponce, L. M. Bautista, and J. C. Alonso. 2016. Dietary divergence in the most sexually size-dimorphic bird. Auk 133:178-197. https://doi.org/10.1642/AUK-15-206.1
Chapuis, J. L. 1979. Evolution saisanniere du regime alimentaire d’Oryctolagus cuniculus dans differents types d’habitats. En France. Pages 743-761 in Proceedings of the world lagomorph conference. University of Guelph, Ontario, Canada.
Dilks, T. J. K., and M. C. F. Proctor. 1979. Photosynthesis, respiration and water content in bryophytes. New Phytologist 82:97-114. https://doi.org/10.1111/j.1469-8137.1979.tb07564.x
Emlen, J. M. 1966. The role of time and energy in food preference. American Naturalist 100:611-617. https://doi.org/10.1086/282455
Glime, J. M. 2017. Birds and bryophytic food sources. Chapter 16-2 in Bryophyte ecology. Volume 2. Bryological Interaction. eBook sponsored by Michigan Technological University and the International Association of Bryologists.
Guo, L. 2020. Exploring the cold-resistant mechanism of the Blood Pheasant (Ithaginis cruentus) based on its feeding habits. Thesis. Xinjiang Agricultural University, Xinjiang, China.
Hammerschlag, N., D. Ovando, and J. E. Serafy. 2010. Seasonal diet and feeding habits of juvenile fishes foraging along a subtropical marine ecotone. Aquatic Biology 9:271-290. https://doi.org/10.3354/ab00251
Hansen, C. E., and P. Rossi. 1990. Arachidonic and eicosapentaenoic acids in brachytheciaceae and hypnaceae moss species. Phytochemistry 29:3749-3754. https://doi.org/10.1016/0031-9422(90)85325-A
Hoak, J. C. 1997. Fatty acids in animals: thrombosis and hemostasis. American Journal of Clinical Nutrition 65:1683S-1686S. https://doi.org/10.1093/ajcn/65.5.1683S
Hoffman, R. M., J. A. Wilson, D. S. Kronfeld, W. L. Cooper, L. A. Lawrence, D. Sklan, and P. A. Harris. 2001. Hydrolyzable carbohydrates in pasture, hay, and horse feeds: direct assay and seasonal variation. Journal of Animal Science 79:500-506. https://doi.org/10.2527/2001.792500x
Holechek, J. L., M. Vavra, and R. D. Pieper. 1982. Botanical composition determination of range herbivore diets: a review. Journal of Range Management 35:309-315. https://doi.org/10.2307/3898308
Iwatsuki, Z., and M. Mizutani. 1981. Coloured illustrations of bryophytes of Japan. Hoikusha, Osaka, Japan.
Jia, C. X., and Y. H. Sun. 2008. Blood pheasant. Chinese Journal of Zoology 5:143.
Jia, C. X., Y. H. Sun, and J. E. Swenson. 2010. Unusual incubation behavior and embryonic tolerance of hypothermia by the Blood Pheasant (Ithaginis cruentus). Auk 127:926-931. https://doi.org/10.1525/auk.2010.09254
Lázaro, X. A., R. Mackenzie, and J. E. Jiménez. 2021. Evidence of endozoochory in Upland Geese Chloephaga picta and White-bellied Seedsnipes Attagis malouinus in sub-Antarctic Chile. Ecology and Evolution 11:9191-9197. https://doi.org/10.1002/ece3.7725
Levin, S. A., S. R. Carpenter, H. C. J. Godfray, A. P. Kinzig, M. Loreau, J. B. Losos, and B. Walker, D. S. Wilcove, editors. 2009. The Princeton guide to ecology. Princeton University Press, Princeton, New Jersey, USA. https://doi.org/10.1515/9781400833023
Li, C. Q., and D. H. Li. 1981. Blue-eared Pheasant and Blood Pheasant in the forestry of Qi-Lian Xian, Qinghai. Zoological Research 2:77-82.
Lou, Y. Q., Y. Hong, J. Tian, and Y. H. Sun. 2014. Habitat selection of Blood Pheasant in spring and summer at Lianhuashan. Sichuan Journal of Zoology 33:229-233.
Luo, X., T. P. Wu, and A. Q. Huang. 2016. Diet analysis and foraging strategy of two sympatric pheasants at Mt. Gaoligong in winter. Chinese Journal of Ecology 35:1003-1008.
Maya-García, O., E. Maya-Elizarrarás, C. Lara, and J. E. Schondube. 2024. Arthropod foraging in a temperate mountain hummingbird ensemble. Birds 5:774-797. https://doi.org/10.3390/birds5040052
Nam, M. H., S. K. Jeong, Y. S. Lee, J. M. Choi, and H. G. Kim. 2006. Effects of nitrogen, phosphorus, potassium and calcium nutrition on strawberry anthracnose. Plant Pathology 55:246-249. https://doi.org/10.1111/j.1365-3059.2006.01322.x
Pejin, B., L. Vujisic, M. Sabovljevic, V. Tesevic, and V. Vajs. 2012. The moss Mnium hornum, a promising source of arachidonic acid. Chemistry of Natural Compounds 48:120-121. https://doi.org/10.1007/s10600-012-0175-7
Popper, Z. A. 2008. Evolution and diversity of green plant cell walls. Current Opinion Plant Biology 11:286-292. https://doi.org/10.1016/j.pbi.2008.02.012
Prins, H. H. T. 1982. Why are mosses eaten in cold environments only? Oikos 38:374-380. https://doi.org/10.2307/3544680
Putman, R. J. 1984: Facts from faeces. Mammal Review 14:79-97. https://doi.org/10.1111/j.1365-2907.1984.tb00341.x
Reeves, III, J. B. 1987. Lignin and fiber compositional changes in forages over a growing season and their effects on in vitro digestibility. Journal of Dairy Science 70:1583-1594. https://doi.org/10.3168/jds.S0022-0302(87)80186-8
Russo, N. J., M. Robertson, R. MacKenzie, B. Goffinet, and J. E. Jiménez. 2020. Evidence of targeted consumption of mosses by birds in sub‐Antarctic South America. Austral Ecology 45:399-403. https://doi.org/10.1111/aec.12858
Scheiffarth, G. 2001. The diet of Bar-tailed Godwits Limosa lapponica in the Wadden Sea: combining visual observations and feces analyses. Ardea 89:481-494.
Shi, D. C., and G. H. Li. 1985. A preliminary study on the Blood Pheasant diet at Baihe Reserve Nanping, Sichuan. Zoological Research 6:137-145.
Sih, A., and B. Christensen. 2001. Optimal diet theory: when does it work, and when and why does it fail? Animal Behaviour 61:379-390. https://doi.org/10.1006/anbe.2000.1592
Sun, Y. H., J. E. Swenson, Y. Fang, S. Klaus, and W. Scherzinger. 2003. Population ecology of the Chinese Grouse, Bonasa sewerzowi, in a fragmented landscape. Biological Conservation 110:177-184. https://doi.org/10.1016/S0006-3207(02)00187-8
Sutherland, W. J., I. Newton, and R. Green. 2004. Bird ecology and conservation: a handbook of techniques: a handbook of techniques (No. 1). Oxford University Press, New York, New York, USA.
Tallima, H., and R. El Ridi. 2018. Arachidonic acid: physiological roles and potential health benefits - a review. Journal of Advanced Research 11:33-41. https://doi.org/10.1016/j.jare.2017.11.004
Turner, J. C. 1979. Adaptive strategies of selective fatty acid deposition in the bone marrow of desert bighorn sheep. Comparative Biochemistry and Physiology Part A: Physiology 62:599-604. https://doi.org/10.1016/0300-9629(79)90110-5
Wang, N., B. C. Tan, and G. W. H. Davison. 2011. Bryophytes as a major dietary component for the Blood Pheasant, Ithaginis cruentus. Journal of Bryology 33:257-260. https://doi.org/10.1179/1743282011Y.0000000018
Whelan, C. J., Ç. H. Şekercioğlu, and D. G. Wenny. 2015. Why birds matter: from economic ornithology to ecosystem services. Journal of Ornithology 156:227-238. https://doi.org/10.1007/s10336-015-1229-y
Wielgolaski, F. E., S. Kjelvik, and P. Kallio. 1975. Mineral content of tundra and forest tundra plants in Fennoscandia. Pages 316-332 in F. E. Wielgolaski, editor. Fennoscandian tundra ecosystems. Part 1 plants and microorganisms. Springer, Berlin, Germany. https://doi.org/10.1007/978-3-642-80937-8_36
Wingard, G. J., R. B. Harris, D. H. Pletscher, D. J. Bedunah, B. Mandakh, S. Amgalanbaatar, and R. P. Reading. 2011. Argali food habits and dietary overlap with domestic livestock in Ikh Nart nature reserve, Mongolia. Journal of Arid Environments 75:138-145. https://doi.org/10.1016/j.jaridenv.2010.09.014
Yao, J. C. 1992. Blood Pheasant forage the species of bryophytes. Sichuan Journal of Zoology 3:10-13.
Fig. 1
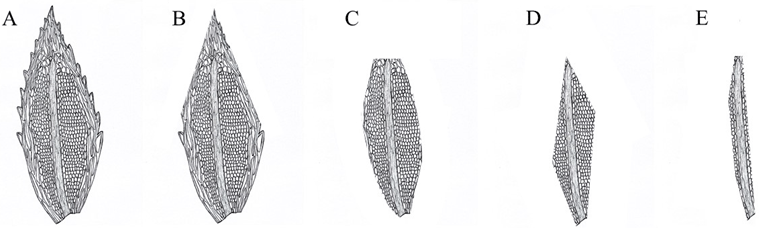
Fig. 1. Diagram illustrating the leaf structure of Mniaceae (400X magnification). A: original leaf structure; B: representation of the first level of bryophyte digestibility in the fecal samples; C: the second level of bryophyte digestibility; D: the third level of bryophyte digestibility; E: the fourth and highest level of bryophyte digestibility in the fecal samples.
Fig. 2
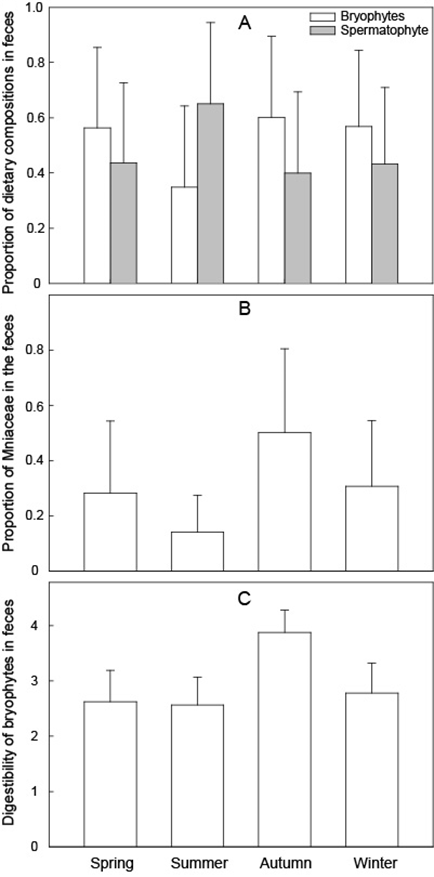
Fig. 2. Seasonal variation in dietary components in Blood Pheasant (Ithaginis cruentus) fecal samples collected at Lianhuashan from 2011 to 2013. (A) seasonal variation in the main dietary components in fecal samples. Blank bar represents bryophytes and grey represents spermatophytes; (B) Seasonal variation in the proportion of Mniaceae in fecal samples; and (C) Seasonal variation in digestibility of bryophytes in fecal samples. The upper whiskers of the bars represent the standard deviation in the figures.
Table 1
Table 1. The occurrences of main moss families in the feces of Blood Pheasant (Ithaginis cruentus) at Lianhuashan Nature Reserve from April 2011 to October 2013.
| Season | I | II | III | IV | V | VI | VII | Digestibility | |
| Spring | 97.9 | 40.4 | 29.8 | 4.3 | 66.0 | 4.3 | 10.6 | 2.65 (1–3) | |
| Summer | 95.7 | 43.5 | 34.8 | 0 | 78.3 | 8.7 | 8.7 | 2.57 (2–3) | |
| Autumn | 97.6 | 58.5 | 0 | 17.1 | 43.9 | 4.9 | 7.3 | 3.87 (2–4) | |
| Winter | 97.3 | 81.1 | 37.8 | 8.1 | 40.5 | 2.7 | 24.3 | 2.78 (2–4) | |
| Note: the number represents each family as the following: Mniaceae (I), Hypnaceae (II), Brachytheciaceae (III), Pottiaceae (IV), Thuidiacea (V), Sematophyllaceae (VI), Entodontaceae (VII) during four seasons. The digestibility was shown as mean (ranges). The values of occurrences were shown in proportions (%). | |||||||||
Table 2
Table 2. Multiple comparisons of bryophytes and spermatophytes in Blood Pheasant (Ithaginis cruentus) across seasons.
| Diet | Seasons | Z | P value | ||||||
| Bryophytes | Spring vs summer | 2.753 | 0.018 | ||||||
| Spring vs autumn | -0.750 | 1 | |||||||
| Spring vs winter | 0.016 | 1 | |||||||
| Summer vs autumn | -3.292 | 0.003 | |||||||
| Summer vs winter | -2.619 | 0.026 | |||||||
| Autumn vs winter | 0.714 | 1 | |||||||
| Spermatophytes | Spring vs summer | -2.753 | 0.018 | ||||||
| Spring vs autumn | 0.750 | 1 | |||||||
| Spring vs winter | -0.012 | 1 | |||||||
| Summer vs autumn | 3.291 | 0.003 | |||||||
| Summer vs winter | 2.619 | 0.026 | |||||||
| Autumn vs winter | -0.714 | 1 | |||||||
| Note: P values are corrected by Bonferroni, and significant values are in bold. | |||||||||
Table 3
Table 3. Multiple comparisons of percentages of Mniaceae in the fecal samples in Blood Pheasant (Ithaginis cruentus) across seasons.
| Seasons | Z | P | |||||||
| Spring vs summer | 0.844 | 1 | |||||||
| Spring vs autumn | -4.697 | < 0.001 | |||||||
| Spring vs winter | -4.656 | 0.658 | |||||||
| Summer vs autumn | -1.228 | < 0.001 | |||||||
| Summer vs winter | -1.818 | 0.207 | |||||||
| Autumn vs winter | 3.220 | 0.004 | |||||||
| Note: P values are corrected by Bonferroni, and significant values are in bold. | |||||||||
Table 4
Table 4. Multiple comparisons of bryophytes digestibility in Blood Pheasant (Ithaginis cruentus) across seasons.
| Seasons | Z | P | |||||||
| Spring vs summer | 0.466 | 1 | |||||||
| Spring vs autumn | -8.028 | < 0.001 | |||||||
| Spring vs winter | -0.827 | 1 | |||||||
| Summer vs autumn | -7.007 | < 0.001 | |||||||
| Summer vs winter | -1.126 | 0.780 | |||||||
| Autumn vs winter | 6.731 | < 0.001 | |||||||
| Note: P values are corrected by Bonferroni, and significant values are in bold. | |||||||||