







The following is the established format for referencing this article:
Covino, K. M., F. D. Titmuss, J. E. Brosseau, D. C. Donacik, M. E. Ellms, A. E. Fisante, O. K. Maday, A. R. Nowicki, D. Okuyama, J. M. Robinson, K. A. Schickfus, E. L. Stanley, and M. E. Everett. 2023. Brilliant bird brains: object recognition in Herring Gulls (Larus argentatus smithsonianus). Journal of Field Ornithology 94(4):7.ABSTRACT
The ability to recognize and associate objects with prior experiences has been demonstrated in several avian taxa. Corvids, for example, recognize visual stimuli associated with negative previous experiences and will modify their behavioral responses according to the level of perceived threat. However, the extent to which similar memory-based behaviors exist in seabirds is poorly understood. Our study investigated object recognition by Herring Gulls (Larus argentatus smithsonianus) on their breeding grounds, the site of a long-term monitoring project. When handling gulls and conducting field research, researchers wear standard bicycle helmets as a protective measure against physical aggression by nesting gulls. We tested whether previously banded gulls vary their behavioral responses based on the headgear worn by human researchers. Herring Gulls were significantly more aggressive when approached by helmeted observers compared to those wearing neutral headwear or even holding a helmet, thus indicating a potential association between wearing bike helmets and the negative experience with researchers. Our study demonstrates that Herring Gulls are capable of object memory and object recognition within context, and display different behaviors toward different objects according to their levels of perceived threat. This information is key when designing studies with nesting gulls to ensure that behavioral biases are not unintentionally created due to gull prior experiences or researcher safety gear.
RESUMEN
La habilidad para reconocer y asociar objetos con experiencias anteriores ha sido demostrada en varios grupos de aves. Los Cuervos, por ejemplo, reconocen estímulos visuales asociados con experiencias negativas en el pasado y modifican sus respuestas comportamentales de acuerdo con el nivel de amenaza percibido. Sin embargo, el nivel al cual comportamientos similares, basados en la memoria, en aves marinas ha sido poco comprendido. Nuestro estudio investigó el reconocimiento de objetos en Larus argentatus smithsonianus en su zona de reproducción, el sitio de un proyecto de monitoreo a largo plazo. Cuando se manipulan los individuos y se realiza la investigación de campo, los investigadores usan cascos de bicicleta como una medida de protección contra las agresiones físicas de gaviotas anidando. Evaluamos si las gaviotas que han sido anilladas anteriormente varían en sus respuestas comportamentales basados en el tipo de preda en la cabeza utilizada por los investigadores humanos. Los individuos de L.a.smithsonianus fueron significativamente más agresivos cuando fueron aproximados por investigadores utilizando cascos comparado con investigadores utilizando prendas neutrales en la cabeza o incluso con los cascos en las manos, indicando una asociación potencial entre el uso de cascos y la experiencia negativa con los investigadores. Nuestro estudio demuestra que los individuos de L.a.smithsonianus son capaces de generar una memoria de los objetos y de reconocimiento de los objetos con un contexto, y desplegar diferentes comportamientos hacia diferentes objetos de acuerdo con los niveles de amenaza percibidos. Esta información es clave para el diseño de estudios en anidación de gaviotas para asegurar que los sesgos comportamentales no son creados sin intención debido a las experiencias previas de las gaviotas o las prendas de seguridad de los investigadores.
INTRODUCTION
Cognition and memory have long been topics of ornithological interest, and the effects of memory on avian behavior have been relatively well-studied (Emery 2006). Past research has demonstrated that multiple species of birds possess episodic memory, indicating the ability to have a detailed recollection of past personal experiences. For example, Common Ravens (Corvus corax) have a demonstrated ability to store, categorize, and recall information (Müller et al. 2017); likewise, American Crows (C. brachyrhynchos) can form positive or negative associations with an object based on their prior experiences with that object (Marzluff et al. 2010). Additionally, both Rock Pigeons (Columba livia) and Keas (Nestor notabilis) have similar recognition and association skills, and display an ability to recognize objects in pictures and associate them with positive stimuli (Soto and Wasserman 2014, Wein et al. 2015).
Although the field of animal behavior was initially established with the study of gull behaviors (Tinbergen 1953, 1960), research on memory and object recognition and their associated behavioral effects in gulls (Laridae) is sparse. Studies of avian memory and cognition tend to focus on relatively few taxa—primarily Corvidae (e.g., Marzluff et al. 2010, Smirnova et al. 2020), Columbiformes (Peissig et al. 2006, Soto and Wasserman 2014), and Psittaciformes (Pepperberg 1994, 2006, Wein et al. 2015). Even with the taxonomically skewed research on avian memory, some researchers have drawn inferences on the similarities between avian and human cognition (e.g., Soto and Wasserman 2012, 2014, Darby et al. 2018). That said, several studies have explored a variety of behaviors in gulls, including detailing their breeding and foraging behaviors (e.g., Tinbergen 1953, 1959, Annett and Pierotti 1989), their ability to recognize and distinguish the auditory signals of threats from non-threats (MacLean and Bonter 2013), individual recognition between adults, chicks, and family groups (e.g., Beer 1969, Miller and Emlen 1975), and even the ability to solve small puzzles (Neibuhr 1983). Given this wealth of behavioral studies in gulls, and that gulls can serve as a sentinel or early indicator of the effects of climate change (Cimino et al. 2022), it is unclear why few, if any studies have directly explored the possibility of cognition and memory in this taxonomic group.
Banding studies of gulls are crucial for both basic and applied inquiries into their biology, and must include research across species and years. Long-term studies of gull nesting colonies provide a unique opportunity to study a wide variety of biological phenomena, including breeding biology, longevity, and population trends (e.g., Pierotti and Annett 2001, Ellis and Good 2006, Bonter et al. 2016). One such long-term banding and monitoring project takes place along the northwest Atlantic coast, off the Maine–New Hampshire border, where researchers routinely band and study the biology of breeding American Herring Gulls (Larus argentatus smithsonianus) and Great Black-backed Gulls (L. marinus) (Ellis et al. 2005, 2007; MacLean and Bonter 2013, Crisologo and Bonter 2016). Prior research on both this and other study systems has indicated that breeding gulls may be distressed by researcher presence, which has the potential to negatively affect breeding success (Brown and Morris 1994, MacLean and Bonter 2013). Anecdotal evidence suggests that the breeding gulls of Appledore recognize researchers versus non-researchers (Gulls of Appledore Research Group, personal observation). While other avian species have been shown to recognize researchers after research activities such as capture and banding have been conducted (Marzluff et al. 2010, 2012), this has yet to be demonstrated in gulls. In addition to the safety of the gulls, researchers’ safety is a high priority, and since nesting gulls may respond defensively to perceived threats, field researchers frequently wear bicycle helmets when performing research tasks, including banding.
We sought to determine whether Herring Gulls respond specifically to the sight of a particular recognizable object—bicycle helmets—which are worn by gull researchers on Appledore Island for personal safety purposes. We hypothesized that Herring Gulls would exhibit different behavioral responses depending on the presence of our focal object, the helmet. Specifically, we predicted that relative to control trials, a researcher wearing a helmet would elicit the greatest intensity of behavioral response due to stressful past experiences involving helmeted researchers.
METHODS
Study site
This study was performed on Appledore Island, Maine (42.9887°, -70.6135°). Appledore is a 38-ha (95-acre) island within the Isles of Shoals, a small archipelago approximately 13 km (8 miles) east of Rye, New Hampshire, and is the site of various educational and research activities, each of which exposes nesting gulls in the central portion of the island to regular human interaction throughout their breeding season. In particular, the island is home to the Shoals Marine Laboratory and a seasonal songbird banding station (Appledore Island Migration station), and serves as the field site for the Gulls of Appledore Research Group. Herring Gulls and Great Black-backed Gulls breed on Appledore from late April through August (Ellis and Good 2006). From 2004 to the present, the Gulls of Appledore Research Group has conducted banding of Great Black-backed Gulls each season, and through 2017, the group also regularly banded both adult and pre-fledgling Herring Gulls; only a small number of additional Herring Gulls have been banded since 2017.
Adult banding has typically taken place early during incubation (mid-May), and birds have been captured with nest traps or by hand at the nest. Banding of chicks has typically occurred during mid-July; pre-fledgling chicks have been captured by hand prior to their ability to fly. All individuals have been banded with stainless steel USGS and a 3-digit field-readable PVC leg band. Morphometric measurements, including mass, head–bill (distance from the occipital to the tip of the bill), and sometimes wing chord length have been taken and recorded. During banding and research activities throughout the summer, researchers wear helmets, typically standard bicycle helmets, but occasionally snowboarding or skateboarding helmets. Due to the danger of physical flighted attacks by gulls (particularly the Great Black-backed Gulls) defending their nests, the practice of wearing a helmet as standard field protective equipment began circa 2008 and has been a consistent requirement of field researchers since 2010.
Study species
This study focused on Herring Gulls nesting near the buildings of Shoals Marine Laboratory (Fig. 1). Selection of focal individuals around the campus buildings ensured that gulls had regular interactions with non-researchers (students at field dorms), in addition to past interactions with researchers during banding and ongoing monitoring projects. Nesting initiation peaks around 14 May for Appledore’s Herring Gulls (M. Everett, unpublished data), and incubation of their typical clutch of three eggs lasts 30–32 days (Weseloh et al. 2020). While male Herring Gulls tend to be slightly larger than females, the sexes are otherwise similar, and both males and females participate in incubation, brooding, and chick provisioning (Gulls of Appledore Research Group, unpublished data; Weseloh et al. 2020). The 25 individuals included in this study were banded by the Gulls of Appledore Research Group between 1 and 17 years prior to our study (mean ± SD = 8.5 ± 3.37 years). Nineteen of the gulls used in this study were banded as hatch-year chicks and ranged in age from 5 to 17 years (mean ± SD = 9.3 ± 3.28 years, median = 8). The remaining six were banded as fully mature, after-fourth-year adults and ranged in age from at least 5 years to at least 11 years. Using only previously banded Herring Gulls allowed for repeated testing of individuals in our randomized experimental trials.
Experimental trials
We tested the Herring Gulls between 31 May 2022 and 4 June 2022, while all focal individuals were incubating eggs (mean ± SD = 2.76 ± 0.44 eggs; range 1–3 eggs). Focal gulls experienced each of our four experimental trials in a randomized order, with a minimum of 2 hours between trials. The four trials were as follows: (1) researcher wearing a standard black bicycle helmet on their head, (2) researcher carrying the same helmet by the straps at their side, (3) researcher wearing a solid grey baseball cap, and (4) researcher with no headwear. Field researchers wore neutral, dark-colored clothing (blacks, browns, blues) for every trial. Semi-randomized trial assignments across the seven field researchers guaranteed that each gull experienced only one trial by an individual researcher, none of whom were personally involved in the prior banding or prior year’s research on the Appledore gulls. Each focal individual experienced the four trials over a maximum of 5 days (mean = 2.76 days) to minimize the effects of seasonal changes in behavioral response (e.g., Crisologo and Bonter 2016). All trials were conducted between 0630 and 1935 hours and were limited to fair weather conditions.
In each trial, field researchers donned their headwear, if applicable, before entering the focal gull’s field of vision. Researchers walked past the focal nest (1–2 m from the nest’s edge) at a slow, steady pace (0.5–0.6 m/s) while remaining quiet and keeping a neutral expression. The researcher’s gaze was directed at the ground throughout the trial except for a glance upward to assess the gull’s reaction, thus controlling for the effect of eye contact on behavior (Goumas et al. 2019, 2020) (Fig. 2). If the researcher could not read the band on the gull while conducting a trial, they returned immediately after completing the trial to identify the incubating gull by noting the field-readable band code. To assess the behavioral response of each gull, we used a categorical scale (Table 1) (Covino et al., unpublished, as modified from MacLean and Bonter 2013), with categories ranging from no response (score of 0) to a response indicating significant agitation (score of 8). Each gull was accordingly scored numerically in each trial based on its maximum response exhibited. Researchers documented the number of eggs present and whether the focal gull’s mate was present at the time of each trial.
The eggs of one focal individual (Gull ID C17) (Fig. 3) began to hatch before attempting our fourth experimental trial, so we did not complete the hat trial on that individual. Additionally, because both male and female Herring Gulls brood, the focal individual in each pair was not always present at its nest when researchers attempted a trial. In these instances, field researchers waited a minimum of 2 hours before revisiting the nest to test the focal individual. In one case, a focal individual (Gull ID 68J) (Fig. 3) was continually absent during trial attempts; as such, we could complete only three trials and were unable to conduct the hat trial with this individual.
Data analyses
We ran a general linear model to investigate the variation in behavioral response score across individuals and the potential effect of (and interaction with) mate presence. We also ran a general linear model that tested for the effects of trial order and researcher on response score to test whether our semi-random experimental design successfully precluded these variables from influencing behavioral response. We ran a linear mixed-effects model to examine the effect of trial type on response score, with Gull ID, trial order, and researcher as separate random factors. Finally, we also tested for any effects of the number of eggs at the time of each trial by using a separate general linear model (ran separately to prevent overfitting our main mixed effects model). The results of all tests were considered significant at the α = 0.05 level. All analyses were run using R (R Core Team 2021). Linear mixed models were run using packages “lme4” (Bates et al. 2015) and “lmerTest” (Kuznetsova et al. 2017), and multiple comparisons were run using package “multcomp” (Hothorn et al. 2008). For linear mixed models, we used the Satterthwaite’s method to estimate denominator degrees of freedom.
RESULTS
We conducted a total of 98 trials on 25 gulls; as noted previously, the fourth trial was not completed for two individuals. There was a significant amount of variation in scores across individual gulls (F24,55 = 8.63, P < 0.001) (Fig. 3).
There was no effect of the individual researcher (F6,88 = 0.37, P = 0.90) or trial order (F3,88 = 0.48, P = 0.70) on response score. There was a significant effect of headwear on response score (F3,68.9 = 8.13, P < 0.001). Specifically, scores for both wearing a hat and no headwear were approximately 1.6 (± 0.4, SE) lower on our response scale than for wearing a helmet (Fig. 4) (wearing helmet versus hat: z = 4.18, P < 0.001; wearing helmet versus no headwear: z = 4.29, P < 0.001). Further, responses for holding a helmet were approximately 1.1 (± 0.4, SE) lower than wearing a helmet (z = -3.11, P < 0.01). All other pairwise comparisons were non-significant (Fig. 4) (holding helmet versus hat: z = 1.23, P = 0.61; no headwear versus hat: z = 0.08, P = 1.00; holding helmet versus no headwear: z = 1.17, P = 0.64). We found no effect on response score due to mate presence (F1,55 = 1.12, P = 0.33) or the number of eggs in the nest (F1,94 = 1.32, P = 0.25).
DISCUSSION
As evidenced by their varying responses to researchers wearing or carrying different types of headwear, Herring Gulls displayed memory vis-à-vis object recognition. Individual gulls exhibited different behavioral responses when a helmet was worn compared to when a helmet was carried, when a hat was worn, or when there was no headwear worn or present. Since all Herring Gulls in this study were previously banded and had experienced field researchers in helmets during banding, subsequent field projects, or both, our study suggests that Herring Gulls associate the helmets with the field researchers specifically, over general human presence. Future investigations into the effect of the helmet on gull behavioral responses should incorporate more specific tests of helmet position; while all researchers held the helmet at their side in this study, the particular orientation with respect to the focal gull (whether it was held on the side of the researcher nearest the gull/nest) was not standardized. The gulls exhibited greater response intensities when researchers wore a helmet versus when they carried the helmet, which suggests that the object’s position may influence object recognition. However, the potential inconsistency of helmet position may be responsible for the relatively high variation observed in the “holding helmet” trial results as a product of varying helmet visibility (Fig. 4).
Our results are similar to a prior study on American Crows which responded aggressively to a mask that researchers previously wore during the banding process (Marzluff et al. 2010). Much like the crows, the Herring Gulls in this study exhibited a level of precision in their ability to discriminate between a learned stimulus (a helmet) and a similar but distinct alternative stimulus (a hat). In addition, Nováková et al. (2020) recently found that Red-backed Shrikes (Lanius collurio) did not attack a potential threat—a dummy of a predator—when its features were scrambled. However, they were more wary of the scrambled predator than the neutral control dummy. While the Herring Gulls in this study had similar responses to researchers holding the helmet compared to researchers wearing a hat or not wearing any headwear, we contend that the inconsistencies in helmet position preclude our ability to directly compare this particular result; this should be investigated further.
Maclean and Bonter (2013) demonstrated that both Herring Gulls and Great Black-backed Gulls are alarmed by human presence. In their study, which was also conducted on Appledore Island, gulls responded similarly or more intensely to human presence compared to predator audio recordings. Most nests in our study were located in areas that experienced regular human activity throughout the 5-day study period; thus, the possibility that our focal gulls also encountered the presence of other humans over the course of the study was high if not ubiquitous. However, given that most gulls in this study (22 of 25) responded with some level of alarm (response score ≥ 3) in at least one of the four experimental trials (Fig. 3), it is clear based on both this study and that of Maclean and Bonter (2013) that the gulls on Appledore Island are alerted to human presence. Moreover, some focal nests were located near nests of other Herring Gulls and/or Great Black-backed Gulls, which also presented a potential confounding factor for behavioral response intensity. Nest density may influence nest success in gulls, as the predation risk by other gulls intraspecifically and interspecifically may depend on both nest density and whether the neighbors are conspecific (Watanuki 1988, Ellis and Good 2006). While these effects of additional human presence and proximity of conspecifics and heterospecifics may have influenced an individual gull’s response intensity or likelihood, these effects were likely consistent within each focal individual’s experience and thus would not have substantially affected the results of this study.
Although banding projects with Appledore Island Herring Gulls began in 2004, in recent years (since 2017) the gull have only occasionally been banded. The time period between initial banding and our trials ranged from 1 to 17 years (8.5 ± 3.37 years). However, there have been consistent banding and monitoring efforts on the island, which more recently concentrated on breeding Great Black-backed Gulls. Thus, while a large span of time had passed for some individuals since their initial banding experience, because of their nesting locations, our focal Herring Gulls have regularly experienced helmeted researchers conducting their work. Similarly, the age of our focal gulls varied across both those banded as hatch-year chicks (5–17 years) and those banded as adults (at least 5 years to at least 11 years). The unknown precise age of all focal gulls (banded adults were at least 4 years old) precluded our ability to investigate the effects of age on nest defense and object recognition. Future research should explore whether gulls’ responses to helmet-clad researchers vary depending on the time elapsed since banding and/or gull age, but to do so, a study design would have to include a unique helmet or other stimulus used solely during banding and not during other ongoing research efforts.
CONCLUSION
We demonstrated that Herring Gulls recognized a particular object—a bicycle helmet—that is associated with researchers during banding and other studies. The responses of the gulls in this study suggest that they are associating researchers wearing helmets with greater threats of danger. These findings are important because they further expand our understanding of the ways in which birds recognize certain objects or stimuli, and expand the taxa for which this capability has been demonstrated. To our knowledge, no other study has documented object recognition within the Larus genus.
As the environment is continuously modified by humans, bird species are being pressured to adapt their life history strategies to better survive. Appledore Island represents just one example of an active breeding colony of birds coexisting alongside human activity. Human-caused nest disturbance negatively affects reproductive success across a vast taxonomic breadth of seabirds (Fetterolf 1983, Carney and Sydman 1999); however, with the development and implementation of protocols that are mindful of nesting phenology and bird stress when monitoring a breeding colony, the detrimental effects of human involvement can be mitigated (Brown and Morris 1994). For this reason, it is important to understand the factors that may induce stress and activity interruption in breeding seabirds to minimize any harm caused by population monitoring and banding. Researchers should continue to optimize banding and observation protocols with special consideration of the particular breeding biology of their focal species. In examining the potentially negative effects of monitoring and banding on individuals, we may become better able to implement protocols that reduce our impact on our study species overall.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
All co-authors contributed to project development, at least one component of the study itself, data management, and draft composition. FDT supervised field work, conducted database research for focal gull data, and assisted with editing the final manuscript. KMC and MEE supervised the project development, data collection, analyses, and manuscript preparation. KMC edited and revised the final version of the manuscript.
ACKNOWLEDGMENTS
We thank the Shoals Marine Laboratory for allowing access to Appledore Island, its facilities, and the staff support that we relied on to conduct our study. For setting the precedent and providing a body of work and resources, we thank the Gulls of Appledore Research Group, including Sarah J. Courchesne, Julie C. Ellis, and all volunteers past and present. We are grateful for the anonymous donors that support the Gulls of Appledore Research Group, without which the ongoing banding and research would not be possible. Students in the Shoals Marine Laboratory Field Ornithology course received financial support from a variety of sources, including the Art Borror Fund, the Rowley Ornithology Award, and the Shoals Scholars Program. KMC and the Loyola Marymount University Physiology, Hormones, & Avian Biology Lab are supported by the Santa Monica Bay Audubon Society. Finally, we thank Shoals Marine Laboratory Gull Research Intern Ry Andruk for their assistance in experimental design, as well as Field Ornithology student Samuel Munguia Ponce for his contributions to experimental design, data collection, and analysis. Finally, we dedicate this research to L. William Clark who has continually inspired and supported the Appledore Gull research. This is contribution #200 of the Shoals Marine Laboratory.
DATA AVAILABILITY
The data/code that support the findings of this study are openly available in ResearchGate at https://doi.org/DOI:10.13140/RG.2.2.13628.00646. Ethical approval for this research study was granted by the Shoals Marine Lab and University of New Hampshire IACUC (Approval number 210503).
LITERATURE CITED
Annett, C., and R. Pierotti. 1989. Chick hatching as a trigger for dietary switching in the Western Gull. Colonial Waterbirds 12:4-11. https://doi.org/10.2307/1521306
Bates, D., M. Mäechler, B. M. Bolker, and S. C. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67:1-48. https://doi.org/10.18637/jss.v067.i01
Beer, C. G. 1969. Laughing Gull chicks: recognition of their parents’ voices. Science 166:1030-1032. https://doi.org/10.1126/science.166.3908.1030
Bonter, D. N., M. C. Moglia, and L. E. DeFisher. 2016. Sons do not take advantage of a head start: parity in Herring Gull offspring sex ratios despite greater initial investment in males. Journal of Avian Biology 47:121-128. https://doi.org/10.1111/jav.00649
Brown, K. M., and R. D. Morris. 1994. The influence of investigator disturbance on the breeding success of Ring-billed Gulls (Larus delawarensis). Colonial Waterbirds 17:7-17. https://doi.org/10.2307/1521376
Carney, K. M., and W. J. Sydman. 1999. A review of human disturbance effects on nesting colonial waterbirds. Waterbirds 22:68-79. https://doi.org/10.2307/1521995
Cimino, M. A., S. A. Shaffer, H. Welch, J. A. Santora, P. Warzybok, J. Jahncke, I. Schroeder, E. L. Hazen, and S. J. Bograd. 2022. Western Gull foraging behavior as an ecosystem state indicator in coastal California. Frontiers in Marine Science 8:790559. https://doi.org/10.3389/fmars.2021.790559
Crisologo, T. L., and D. N. Bonter. 2016. Defending the weak: parental defense peaks when chick vulnerability is greatest in the Herring Gull (Larus argentatus). Ethology 123:113-122. https://doi.org/10.1111/eth.12578
Darby, K. P., L. Castro, E. A. Wasserman, and V. M. Sloutsky. 2018. Cognitive flexibility and memory in pigeons, human children, and adults. Cognition 177:30-40. https://doi.org/10.1016/j.cognition.2018.03.015
Ellis, J. C., W. Chen, B. O’Keefe, M. J. Shulman, and J. D. Witman. 2005. Predation by gulls on crabs in rocky intertidal and shallow subtidal zones of the Gulf of Maine. Journal of Experimental Marine Biology and Ecology 324:31-43. https://doi.org/10.1016/j.jembe.2005.04.001
Ellis, J. C., and T. P. Good. 2006. Nest attributes, aggression, and breeding success of gulls in single and mixed species subcolonies. Condor 108:211-219. https://doi.org/10.1093/condor/108.1.211
Ellis, J. C., M. J. Shulman, M. Wood, J. D. Witman, and S. Lozyniak. 2007. Regulation of intertidal food webs by avian predators on New England rocky shores. Ecology 88:853-863. https://doi.org/10.1890/06-0593
Emery, N. J. 2006. Cognitive ornithology: the evolution of avian intelligence. Philosophical Transactions of the Royal Society B 361:23-43. https://doi.org/10.1098/rstb.2005.1736
Fetterolf, P. M. 1983. Effects of investigator activity on Ring-Billed Gull behavior and reproductive performance. Wilson Bulletin 95:23-41.
Goumas, M., I. Burns, L. A. Kelley, and N. J. Boogert. 2019. Herring Gulls respond to human gaze direction. Biology Letters 15:20190405. https://doi.org/10.1098/rsbl.2019.0405
Goumas, M., T. R. Collins, L. Fordham, L. A. Kelley, and N. J. Boogert. 2020. Herring Gull aversion to gaze in urban and rural human settlements. Animal Behavior 168:83-88. https://doi.org/10.1016/j.anbehav.2020.08.008
Hothorn, T., F. Bretz, and P. Westfall. 2008. Simultaneous inference in general parametric models. Biometrical Journal 50:346-363. https://doi.org/10.1002/bimj.200810425
Kuznetsova, A., P. B. Brockhoff, and R. H. B. Christensen. 2017. ImerTest package: tests in linear mixed effects models. Journal of Statistical Software 82:1-26. https://doi.org/10.18637/jss.v082.i13
MacLean, S. A., and D. N. Bonter. 2013. The sound of danger: threat sensitivity to predator vocalizations, alarm calls, and novelty in gulls. PLoS ONE 8:e82384. https://doi.org/10.1371/journal.pone.0082384
Marzluff, J. M., R. Miyaoka, and D. J. Cross. 2012. Brain imaging reveals neuronal circuitry underlying the crow’s perception of human faces. Proceedings of the National Academy of Sciences 109:15912-15917. https://doi.org/10.1073/pnas.1206109109
Marzluff, J. M., J. Walls, H. N. Cornell, J. C. Withey, and D. P. Craig. 2010. Lasting recognition of threatening people by wild American Crows. Animal Behavior 79:699-707. https://doi.org/10.1016/j.anbehav.2009.12.022
Miller, D. E., and J. T. Emlen. 1975. Individual chick recognition and family integrity in the Ring-Billed Gull. Behaviour 52:124-144.
Müller, J. J. A., J. J. M. Massen, T. Bugnyar, and M. Osvath. 2017. Ravens remember the nature of a single reciprocal interaction sequence over 2 days and even after a month. Animal Behavior 128:69-78. https://doi.org/10.1016/j.anbehav.2017.04.004
Niebuhr, V. 1983. Feeding strategies and incubation behaviour of wild Herring Gulls: an experiment using operant feeding boxes. Animal Behavior 31:708-717. https://doi.org/10.1016/S0003-3472(83)80226-7
Nováková, N., P. Veselý, and R. Fuchs. 2020. Object categorization by wild-ranging birds in nest defense. Animal Cognition 23: 203-213. https://doi.org/10.1007/s10071-019-01329-3
Peissig, J. J., M. E. Young, E. A. Wasserman, and I. Biederman. 2006. The role of edges in object recognition by pigeons. Perception 34:1353-1374. https://doi.org/10.1068/p5427
Pepperberg, I. M. 1994. Numerical competence in an African Gray Parrot. Journal of Comparative Psychology 108:36-44. https://doi.org/10.1037/0735-7036.108.1.36
Pepperberg, I. M. 2006. Ordinality and inferential abilities of a Gray Parrot (Psittacus erithacus). Journal of Comparative Psychology 120:205-216. https://doi.org/10.1037/0735-7036.120.3.205
Pierotti, R., and C. Annett. 2001. The ecology of Western Gulls in habitats varying in degree of urban influence. Pages 307-329 in J. M. Marzluff, R. Bowman, and R. Donnelly, editors. Avian ecology and conservation in an urbanizing world. Springer. https://doi.org/10.1007/978-1-4615-1531-9_15
R Core Team. 2021. R: a language and environment for statistical computing. Version 4.1.2. R Foundation for Statistical Computing, Vienna, Austria.
Smirnova, A. A., Y. A. Kalashnikova, M. V. Samuleeva, and Z. A. Zorina. 2020. Evaluating the capability of mirror self-recognition in Hooded Crows (Corvus cornix). Biology Bulletin 47:836-843. https://doi.org/10.1134/S106235902007016X
Soto, F. A., and E. A. Wasserman. 2012. Visual object categorization in birds and primates: integrating behavioral, neurobiological, and computational evidence within a “general process” framework. Cognitive, Affective, & Behavioral Neuroscience 12: 220-240. https://doi.org/10.3758/s13415-011-0070-x
Soto, F. A., and E. A. Wasserman. 2014. Mechanisms of object recognition: what we have learned from pigeons. Frontiers in Neural Circuits 8:1-22. https://doi.org/10.3389/fncir.2014.00122
Tinbergen, N. 1953. The Herring Gull’s world: a study of the social behaviour of birds. Frederick A. Praeger, Oxford, UK.
Tinbergen, N. 1959. Comparative studies of the behaviour of gulls (Laridae): a progress report. Behavior 15:1-70. https://doi.org/10.1163/156853960X00098
Tinbergen, N. 1960. The evolution of behavior in gulls. Scientific American 203:118-133.
Watanuki, Y. 1988. Intraspecific predation and chick survival: comparison among colonies of Slaty-Backed Gulls. Oikos 53:194-202. https://doi.org/10.2307/3566062
Wein, A., G. K. Gajdon, and R. Schwing. 2015. Picture – object recognition in Kea (Nestor notabilis). Ethology 121:1059-1070. https://doi.org/10.1111/eth.12423
Weseloh, D. V., C. E. Hebert, M. L. Mallory, A. F. Poole, J. C. Ellis, P. Pyle, and M. A. Patten. 2020. Herring Gull (Larus argentatus), version 1.0. In S. M. Billerman, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA.
Fig. 1
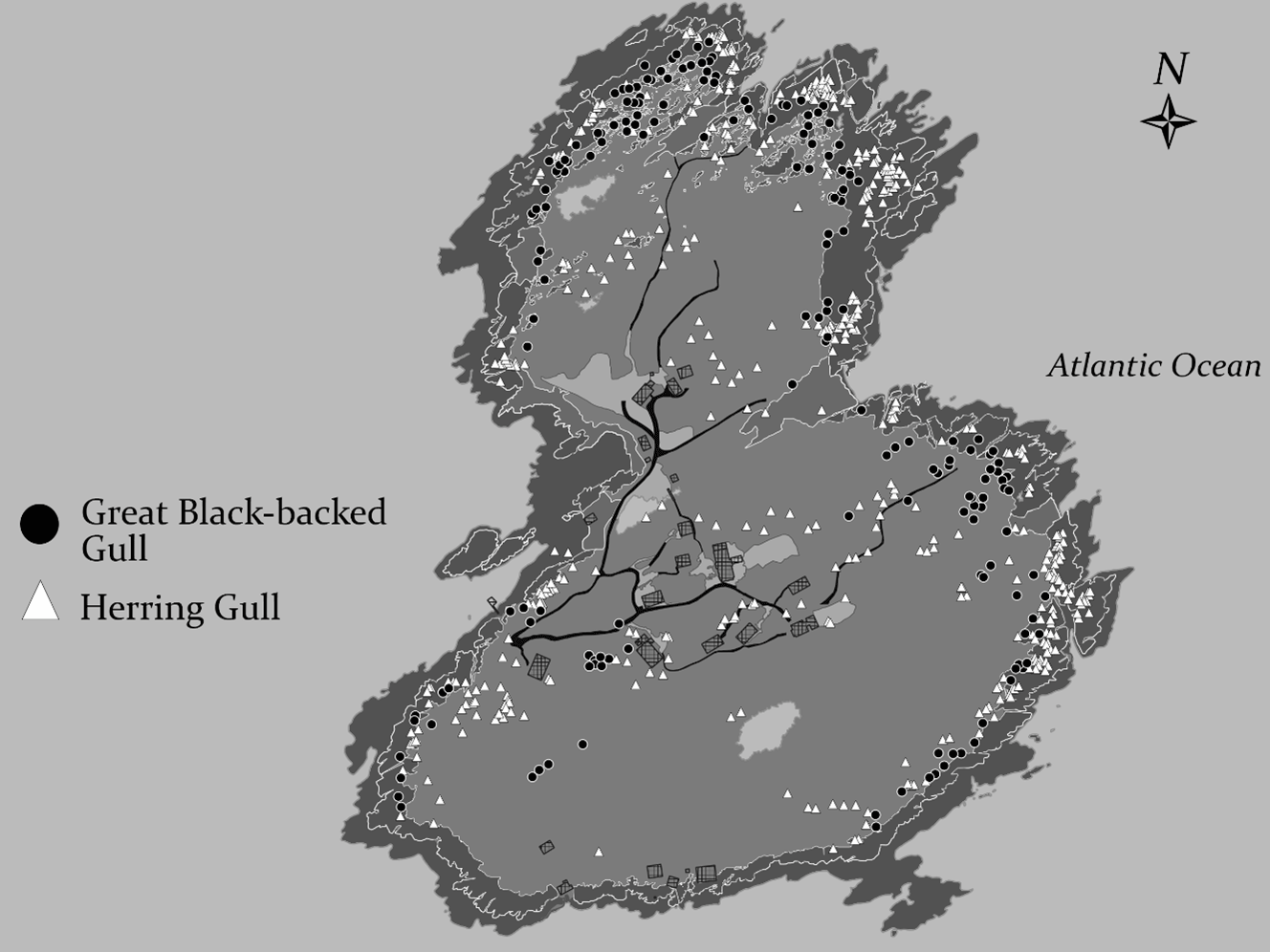
Fig. 1. Map of Appledore Island (42.98°, -70.61°) showing locations of all the island’s gull nests from a 2018 census. Gray hatched boxes represent island buildings, including Shoals Marine Laboratory facilities and private homes.
Fig. 2
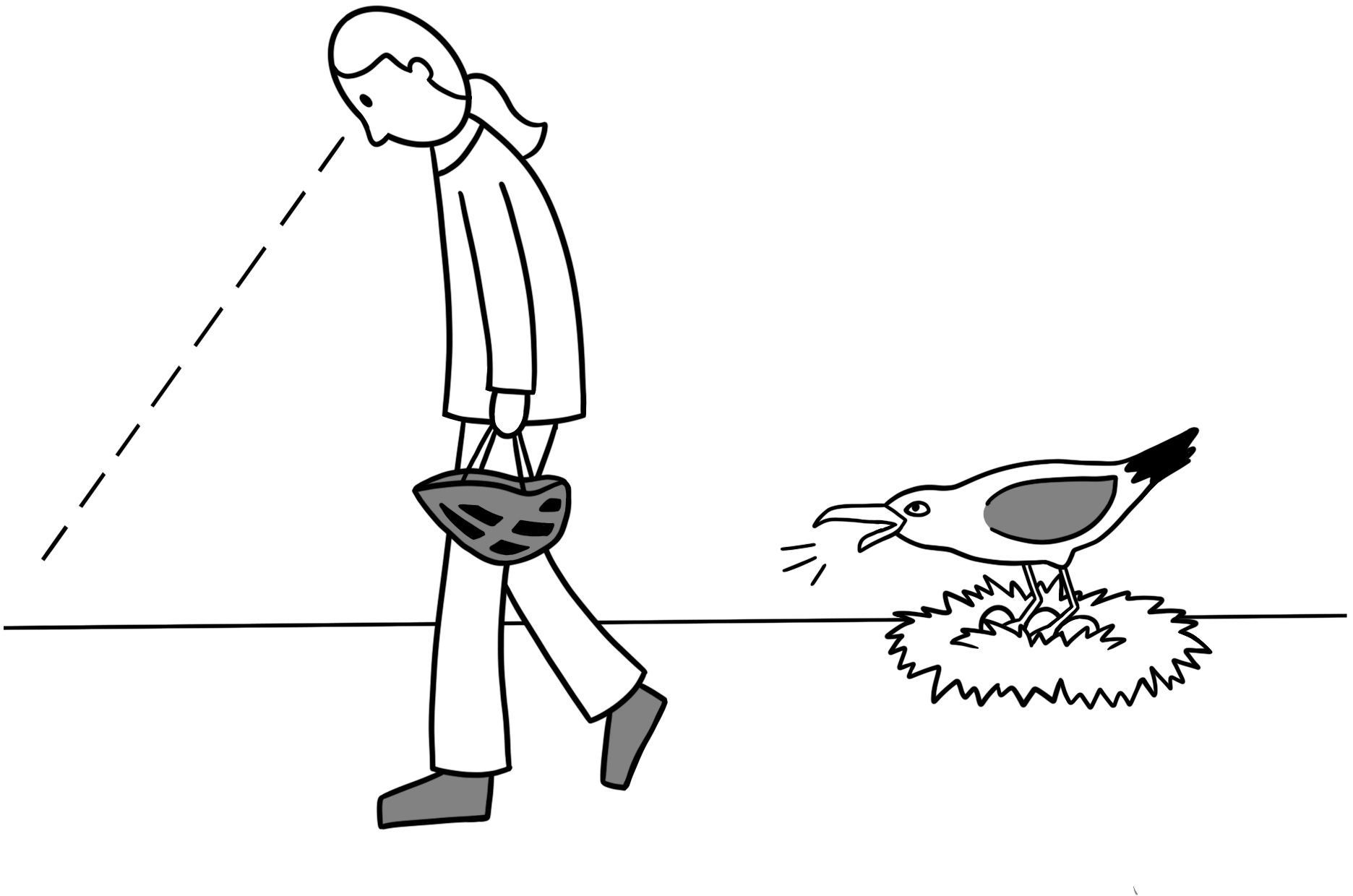
Fig. 2. Depiction of a researcher walking past a nesting gull showing a response of 5 (standing on the nest), including oblique posture. Researcher is performing the “holding helmet” trial in this example. Rendition by Dora Donacik.
Fig. 3
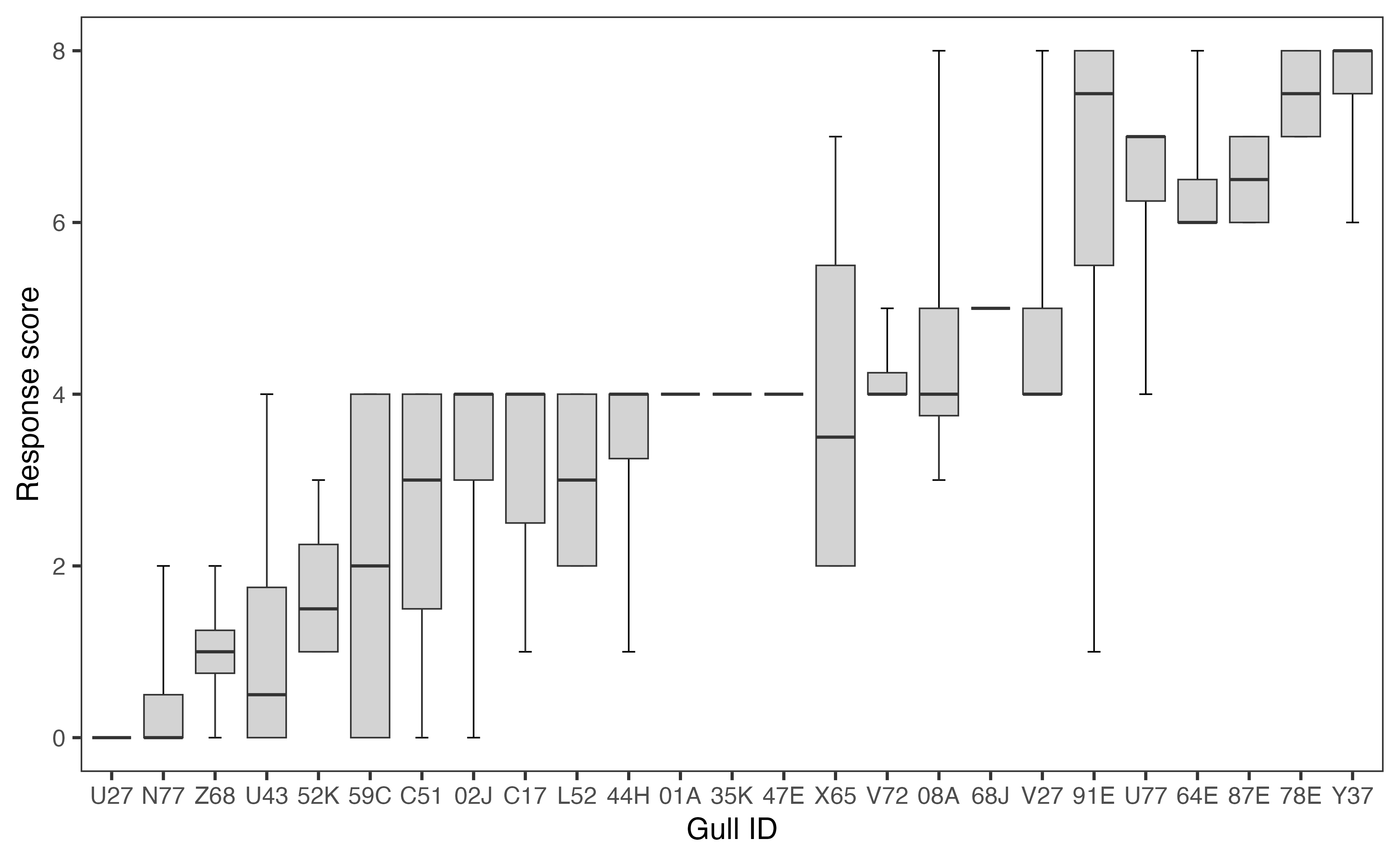
Fig. 3. Distribution of behavioral responses for each gull (n = 25) scored using a categorical scale spanning 0 (lowest) to 8 (highest) (Covino et al., unpublished, as modified from MacLean and Bonter 2013). Each of the 25 boxes represents the response score distribution of an individual gull, with the gull’s band ID labeled on the x-axis. The boxes are arranged from lowest to highest mean response level. The bolded lines represent an individual’s median response, interquartile range is indicated by the grey boxes, and the “whiskers” represent the minimum and maximum score for each bird (including statistical outliers). Individuals that are represented by only a bolded line displayed the same response level in every trial.
Fig. 4
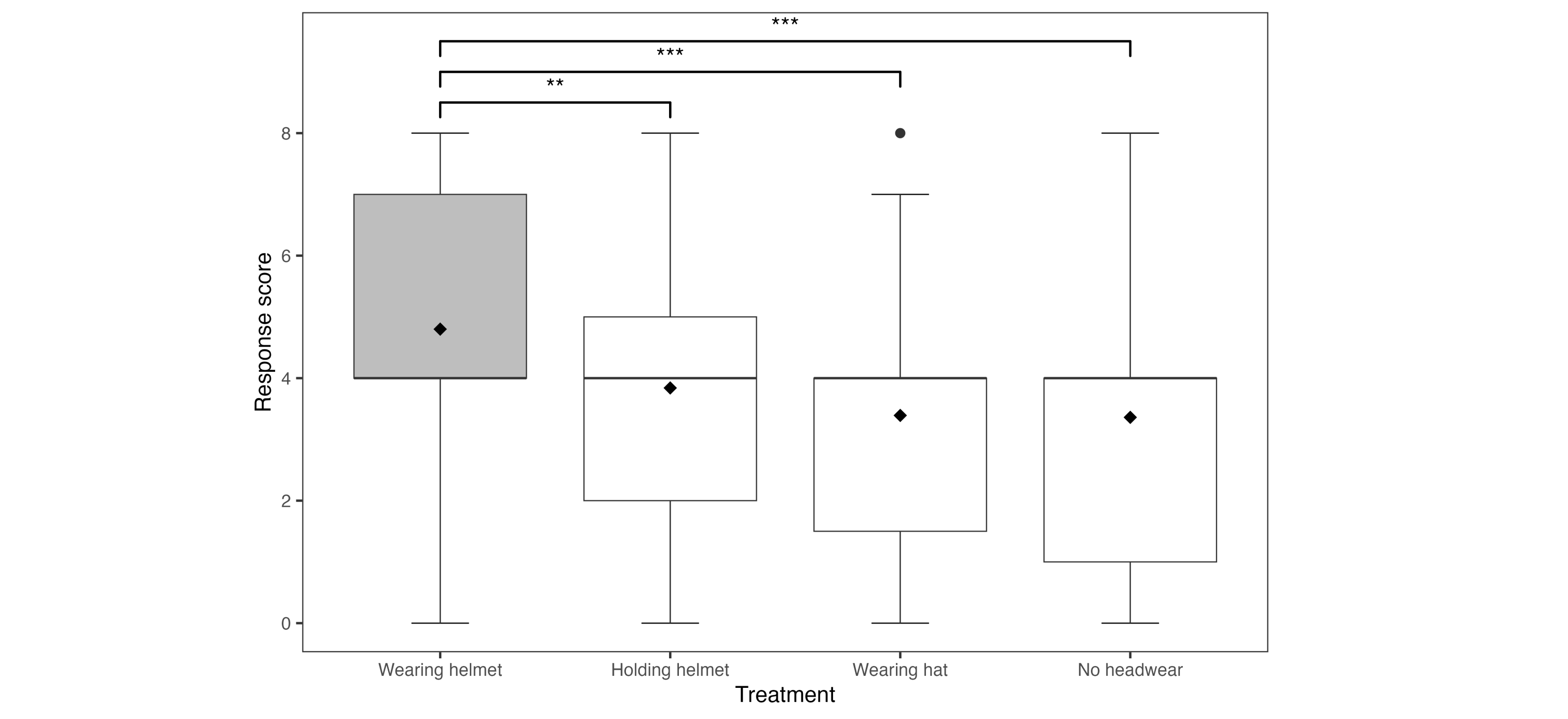
Fig. 4. Distribution of response scores of Herring Gulls (n = 25) across the four trial conditions, scored using a categorical scale where 8 is the highest and most aggressive response (Table 1). Median and mean of each condition are provided by the bolded line and solid dot, respectively. The boxes represent the interquartile range, whiskers represent the first and fourth quartiles, and a statistical outlier value is indicated by the closed dot. Between-group differences are indicated by asterisks at the P < 0.01 (**) and P < 0.001 (***) levels.
Table 1
Table 1. Behavioral response scores used to rate gulls’ overall reaction to the stimuli (Covino et al., unpublished, as modified from MacLean and Bonter 2013). Higher values represent more energetically intensive responses.
| Response value | Response description | ||||||||
| 0 | No response | ||||||||
| 1 | Immediate increase in vigilance in response to stimulus, followed by relaxation | ||||||||
| 2 | Sustained vigilance for entire walk by; neck slightly raised, looking at researcher, but does not interrupt activities | ||||||||
| 3 | Sustained vigilance for entire walk by; neck fully outstretched, looking at researcher, activities often interrupted, use of “kek-kek” calls | ||||||||
| 4 | Extreme vigilance for at least a portion of the walk by, overall vigilance sustained for entire walk by; use of “yeow” calls in combination with vigilance (above-described behaviors) and/or oblique posture while still incubating | ||||||||
| 5 | Extreme vigilance for at least a portion of the walk by, overall vigilance sustained for entire walk by; use of above-described behaviors in combination with interruption of incubation, standing up off eggs (over the nest), and/or oblique posture | ||||||||
| 6 | Extreme vigilance sustained for entire walk by; use of above-described behaviors in combination with movement within 3 m of nest, including localized charging or lunging and/or oblique posture | ||||||||
| 7 | Extreme vigilance sustained for entire walk by; use of above-described behaviors in combination with movement beyond 3 m of the nest, including charging and/or short flights toward the researcher | ||||||||
| 8 | Extreme vigilance sustained for entire walk by; use of above-described behaviors in combination with direct attack behaviors, including swooping flight | ||||||||