







The following is the established format for referencing this article:
Tattoni, D. J., and K. LaBarbera. 2023. A simple method to estimate capture height biases at landbird banding stations: opportunities and limitations. Journal of Field Ornithology 94(4):6.ABSTRACT
Mist-nets are one of the most important tools for the capture of wild birds in ornithological research. The probability of capturing birds may vary by net height, which may drive capture biases. Such biases are rarely estimated, likely because of the relatively high cost and effort associated with constructing and operating elevated mist-net rigs where multiple mist-nets are stacked above one another. Therefore, a low-cost and -effort method to collect capture height data may allow broader investigation and better accounting of potential bias in existing banding protocols. Here, we investigate whether recording net panel of capture (with net panels indicating capture height, e.g., “upper panel”) in ground-level mist-nets provides sufficient information to estimate capture height biases and compare these estimations to those obtained with traditional elevated mist-net rigs. Of the 29 taxa analyzed, we detected elevated capture biases for 11 (37.9%) and ground-level capture biases for seven (24.1%). When compared to estimates derived from elevated mist-net rigs at the same study site, we found high agreement with ground-level biases (75.0%) and low agreement with elevated biases (23.1%). These results suggest panel height of ground-level nets is a reliable method to estimate ground-level biases; however, scale of sampling may influence elevated biases, particularly for species that center their activity at the mid-story. Recording panel height may be quickly integrated into a station’s processing protocols and broader application may improve our understanding of these biases.
RESUMEN
Las redes de niebla son una de las herramientas más importantes para la captura de aves silvestres en investigaciones ornitológicas. La probabilidad de capturar aves puede variar según la altura de las redes, lo que puede conducir a sesgos en las capturas. Estos sesgos son raramente estimados, probablemente por el costo y esfuerzo, relativamente alto, asociado con la construcción y operación de conjuntos de redes de niebla elevadas, donde múltiples redes de niebla son instaladas una sobre la otra. Consecuentemente, un método de bajo costo y esfuerzo, para la colecta de datos de alturas de capturas puede permitir investigaciones más amplias y tener en cuenta de mejor manera el sesgo potencial que existe en los protocolos de anillamiento. Aquí investigamos si registrar el panel de captura (con los paneles de las redes indicando la altura de la captura, e.g. “panel superior”) en redes de niebla a nivel del suelo, proporciona información suficiente para estimar los sesgos en la altura de las capturas y comparar estas estimaciones con las obtenidas con métodos tradicionales de conjuntos de redes de niebla elevados. De los 29 taxones analizados, detectamos sesgos de capturas elevadas en 11 (37.9%) y sesgos de captura a nivel del suelo en siete (24.1%). Cuando comparamos los estimados derivados de conjuntos de redes de niebla elevadas en el mismo sitio de estudio, encontramos una alta coincidencia con los sesgos a nivel del suelo (75%) y baja coincidencia con los sesgos de capturas elevadas (24.1%). Estos resultados sugieren que el muestreo puede influenciar los sesgos de capturas elevadas, particularmente para especies que centran su actividad en estratos medios del bosque. Registrar la altura del panel puede ser integrado rápidamente a los protocolos de procesamiento de una estación y ampliar la aplicación puede mejorar nuestro entendimiento de estos sesgos.
INTRODUCTION
Mist-nets are a widely used and important capture method in the study of wild birds (Dunn and Ralph 2004). Standardized mist-netting (often referred to as “constant effort banding”) is the primary sampling protocol at numerous bird observatories and research stations around the world (DeSante 1992, Dunn and Ralph 2004). Data from mist-netting have been widely applied to demographic (Saracco et al. 2022), conservation (Gray et al. 2022), physiological (de Zwaan et al. 2022), and phenological (Barton and Sandercock 2018) research. Despite its wide use and importance, validation studies to assess potential biases are rarely conducted (but see Remsen and Good 1996, Bonter et al. 2008, Tattoni and LaBarbera 2022).
Potential sources of bias in mist-netting include net avoidance (Roche et al. 2013), net escape (Lövei et al. 2001), and capture height (Remsen and Good 1996). A capture height bias may confound analyses if the vertical activity pattern of a species (or group of interest, e.g., sex) varies by a parameter of interest; e.g., shifts in vertical activity patterns due to habitat succession may result in declining capture rates, obscuring population trends (Remsen and Good 1996). Typically, mist-nets are operated at the “ground level” and sample the vertical space within 2–3 m of the ground. Previous studies that have assessed capture height biases have constructed elevated mist-net rigs that stack two or more mist nets on top of one another (Fitzgerald et al. 1989, Bonter et al. 2008, Tattoni and LaBarbera 2022) or raise a single net to variable heights (Vecchi and Alves 2015) to sample greater vertical space. The high cost and effort associated with constructing these elevated mist-net rigs may contribute to the overall lack of research in this area. Developing a low cost and low effort method for estimating capture height biases would permit easy integration into existing mist-netting protocols and allow more widespread investigation of the impacts of capture height bias on mist-netting data.
Mist-nets have discrete panels (also called shelves or tiers), separated by trammel lines, that form pockets in which birds are captured. In a typical operation, mist-nets are tensioned between two poles that are in a fixed (but adjustable) position. The height of the bottom trammel line should be sufficiently high as to prevent captured birds from hitting the ground. Then, the spacing of the trammel lines above this point will determine the absolute height of the mist-net. Four-paneled mist-nets are widely used at constant effort banding stations (DeSante et al. 2022) but other options are available. A bird may initially fly into only a single panel at each capture event; therefore, the panel provides information on the height of capture. Occasionally a bird may become entangled in netting from adjacent panels after its initial capture (e.g. “double pocketed,” wrapped over a trammel line, etc.); however, with proper extraction techniques the initial panel of capture is usually unambiguous. Net panel capture patterns have been infrequently examined, and mostly in the context of net avoidance, with studies showing greater avoidance of the uppermost versus lowermost net panels (Jenni et al. 1996, Lövei et al. 2001).
The development of a low-effort and low-cost method to detect capture height biases may greatly improve the value of long-term monitoring datasets, particularly those in habitats undergoing succession where changes in vertical activity patterns may confound population trends. Therefore, the goals of our study were to (1) determine whether recording the panel of capture in a four paneled ground-level mist-net would provide sufficient data to estimate capture height biases, and (2) compare these estimates to those obtained with traditional elevated mist-net rigs. We predicted that mist-net panel data would be of sufficient resolution and scale to estimate capture biases and that these biases would agree with those estimated from elevated mist-net rig data.
METHODS
Coyote Creek Field Station (37° 26′ N, 121° 55′ W), Milpitas, California, is located along the lower Coyote Creek where it enters the San Francisco Bay Estuary. The banding station occupies a grassy meadow that functions as a flood control channel and three differently aged stands of riparian woodland (> 70 years, 36 years, and 30 years, respectively; see Table 1 for mean overstory height) where the canopy is dominated by box elder (Acer negundo), Fremont cottonwood (Populus fremontii), and western sycamore (Platanus racemosa) and the understory is dominated by poison hemlock (Conium maculatum), coyote brush (Baccharis pilularis), and elderberry (Sambucus nigra).
We subset our mist-net capture data to include records from ground-level mist-nets within the riparian woodland habitats (excluding data from mist-nets in the overflow channel because capture efficiency of mist-net panels may vary by habitat type; see Lövei et al. 2001). We used capture data from November 2020 through February 2023 from arrays of 13, 14, and 15 ground-level mist-nets, operated on Wednesdays, Saturdays, and Sundays respectively, every week of the year, conditions permitting. During this period, mist-nets were opened approximately 30 minutes prior to sunrise and operated for a total of 5 hours, conditions permitting.
Mist-nets were opened in a standardized manner so that panel heights were comparable across nets: every net pole measured 3.0 m in height with an eyehook set approximately 2.5 cm below the top for guy lines. The net loop of the top trammel line was pushed up until it touched the bottom of the eyehook, and all trammel lines were measured and set 0.54 m apart. PVC poles with an elbow joint to push net loops and markings every 0.54 m were used to allow for rapid and consistent setting and resetting of mist-nets (Fig. 1).
Trained personnel continuously patrolled net lanes to prevent predation events (Churchwell and Barton 2006) and extracted all captured birds with official mist-net checks occurring every 30 minutes. Birds were placed in individual cloth bags and marked with two clothespins noting the mist-net identifier and the panel of capture (“L” for lower, “LM” for lower middle, “UM” for upper middle, and “U” for upper). Occasionally a bird flew into a net when it was being opened or lowered to extract another bird. These birds were not assigned a net panel. Each bird was fitted with an appropriately sized United State Geological Survey band. The mist-net identifier and panel were recorded with each capture event, along with standard demographic and morphological data. We filtered our data to include only the first capture of each individual during the study time period to avoid pseudoreplication.
We modified the modeling approach of Tattoni and LaBarbera (2022) to estimate capture height biases with panel data. The lower and lower-middle panels were grouped together as a binary indicator variable for lower capture (x =1) and the upper-middle and upper panels were grouped together as binary indicator variable for elevated capture (x = 0). Even though panel data provide capture information of higher resolution, we used this binary approach so that our results would be comparable to estimates Tattoni and LaBarbera (2022) obtained using paired ground-level (x = 1) and elevated (x = 0) mist-nets. We then used a binomial model in a Bayesian framework to estimate the probability of lower capture for each taxon in our dataset. Our model was written in the BUGS language (Lunn et al. 2000) and conducted Gibbs sampling of three parallel Markov chain Monte Carlo (MCMC) chains in JAGS v. 4.3.0 (Plummer 2017) through the package “jagsUI” (Kellner 2019) in R v. 3.4.2 (R Core Team 2017). We analyzed all taxa with a sample size n ≥ 13, as is appropriate for simple binomial models in a Bayesian framework (Joseph et al. 1995). We confirmed convergence of estimate parameters by inspecting the trace plots and use of the Gelman-Rubin diagnostic (Gelman et al. 2003). To validate our estimates, we compared results to those of Tattoni and LaBarbera (2022) who used capture data from 1993 to 2020 from paired ground-level and elevated mist-nets at this same study site.
RESULTS
From November 2020 through February 2023, we recorded net panel for 4262 captures. After filtering the first capture record of each band number, we had 3229 captures among 29 taxa with sufficient sample sizes. There was a significant difference in the number of captures across the four panels (X² = 177.73, df = 3, p < 0.005) with the lower-middle and upper-middle panels capturing more than expected (123% and 122%, respectively) and the lower and upper panels capturing less than expected (84% and 70%, respectively). This capture distribution was consistent across the three differently aged woodland stands (Table 1).
The analysis of panel capture data detected significant capture height biases for 18/29 (62.1%) taxa. Biases for capture in the upper-middle and upper panels were found for Nuttall’s Woodpeckers (Dryobates nuttallii), Willow Flycatchers (Empidonax traillii), California Scrub-Jays (Aphelocoma californica), Northern Mockingbirds (Mimus polyglottos), American Robins (Turdus migratorius), House Finches (Haemorhous mexicanus), Lesser Goldfinches (Spinus psaltria), Puget-sound White-crowned Sparrows (Zonotrichia leucophrys pugetensis), Bullock’s Orioles (Icterus bullockii), Audubon’s Yellow-rumped Warblers (Setophaga coronata auduboni), and Myrtle Yellow-rumped Warblers (S. c. coronata; Table 2, Fig. 2). In contrast, Bewick’s Wrens (Thryomanes bewickii), Swainson’s Thrushes (Catharus ustulatus), Hermit Thrushes (Catharus guttatus), Fox Sparrows (Passerella iliaca), Lincoln’s Sparrows (Melospiza lincolnii), Song Sparrows (Melospiza melodia), and Common Yellowthroats (Geothlypis trichas) were biased toward capture in the lower and lower-middle panels (Table 2, Fig. 2).
Overall, we analyzed 28 of the same taxa as Tattoni and LaBarbera (2022) who estimated capture height biases from paired ground-level and elevated mist-net rigs at the same study site (Lesser Goldfinches were only evaluated in this study). There were no instances of opposite biases detected for any taxa. Of the 21 taxa for which a bias was detected in at least one study, 9 (42.9%) had the bias detected in both methods. This agreement varied substantially by bias type: 3/13 (23.1%) for elevated biases and 6/8 (75.0%) for ground-level biases.
DISCUSSION
We found that recording the panel of capture in a standard four-paneled mist-net provided adequate information to estimate taxon-level capture height biases in a bird banding dataset. We detected a significant capture height bias for over half of the taxa analyzed with 11 taxa biased to elevated capture and seven taxa biased to ground-level capture (Fig. 2). These biases mostly followed our expectations based on the life history characteristics of each taxon (e.g., ground foraging sparrow species were biased toward ground-level capture). Our comparison of the analysis of panel data with data from elevated net rigs showed agreement between the two methods in estimating ground-level biases but not elevated biases.
Although there was high agreement of the ground-level biases detected using panel data versus paired net data, there was considerable variation in the species to which were attributed elevated biases. Differences in vertical scale between these sampling techniques may account for this result. Activity patterns for many taxa may extend above the height of a single ground-level net (Remsen and Good 1996); therefore, ground-level nets produce incomplete capture distributions for some species, particularly those favoring the canopy. Elevated mist-net rigs sample twice as much vertical space, which may help produce a more representative sample, particularly if the top of the canopy is near the top of the elevated mist-net. In contrast, for species with ground-level biases, a ground-level net alone appears to capture a representative sample.
The most common discrepancy we found was the detection of elevated biases for taxa that had no bias in the elevated mist-net data (Nuttall’s Woodpeckers, California Scrub-Jays, Northern Mockingbirds, American Robins, House Finches, Puget-sound White-crowned Sparrows, and Bullock’s Orioles). This pattern is expected for species that center their activity pattern in the mid-story (~3 m; Fig. 3C). The smaller vertical space sampled by ground-level nets alone results in data that detect an “elevated bias” at a lower absolute height than for elevated mist-net rigs. Less common were taxa which had an elevated bias in the elevated mist-net data but not in our panel data (Chestnut-backed Chickadees [Poecile rufescens] and Yellow Warblers [Setophaga petechia]). This pattern could be explained by an activity pattern distribution that is centered in the upper-story with a high peak but long tail, such that there is no appreciable difference in captures across the ground-level mist-net panels (Fig. 3D). Additionally, these hypothetical capture distributions may be modified by habitat structure (i.e., higher vegetation height might stretch the tails of capture distributions, making biases difficult to detect with even a stacked mist-net rig, or lower vegetation height might compress capture distributions, making some easier to detected with a ground-level net). Collection of panel data at elevated mist-net rigs nets would allow these theoretical capture patterns to be tested in the future.
An important assumption when using panel data is that of equal bird capture probability among panels. Two avian capture efficiency studies show that net avoidance and escape are highest in the uppermost panel (Jenni et al. 1996, Lövei et al. 2001). If these factors affect our data, we expect ground-level biases to be exaggerated and elevated biases to be masked. We do observe in our dataset that the uppermost panel has the fewest captures, but the difference is moderate (the uppermost panel has 83% as many captures as the next-lowest-capture panel). The effects of differential escape probabilities on our results were likely small because (1) Jenni et al. (1996) showed that this bias was weakest at shaded nets, and all of the mist-nets included in this study were partially or fully shaded; and (2) we operate with near continuous mist-net patrolling wherein birds are extracted closer to their time of capture than with a standard 30-minute net check protocol, which likely results in fewer mist-net escapes. Additionally, large-bodied passerines (> 40 g) have higher escape rates (Jenni et al. 1996) and therefore should be most prone to an escape-driven underestimate of elevated captures, yet three of the species with elevated capture biases in this study were large-bodied passerines (California Scrub-Jays, Northern Mockingbirds, and American Robins).
Recording the net panel of capture can be easily integrated into a banding station’s protocol. Logistically, it requires (1) an additional data field on datasheets or electronic data forms, (2) a method of marking and tracking the panel of capture as birds are transported from mist-nets to the processing location, and (3) training and adjustment of personnel to the new protocol. At our banding station we began collecting panel data in November 2020. We distributed clothespins labeled L, LM, UM, and U to extractors prior to opening the mist-nets each morning. At the time of extraction, extractors attached the appropriate clothespin indicating the panel of capture to each bird holding bag. This was easily rolled into our existing extraction protocols and our extractors quickly adjusted to the addition. We hope the ease of obtaining these data makes it attractive for other banding stations to adopt.
Perhaps the most challenging aspect of integrating panel of capture into an existing banding station is the need to standardize mist-net height and trammel spacing so that each panel corresponds to the same vertical space at each mist-net. Mist-nets are often set up based on “what looks right,” which varies by mist-net brand, individual preferences, and experience/training. Frequently knee height is used as a guide for the bottom trammel line and hand depth as a guide for each pocket and therefore the trammel spacing. The widely used NABC Manual (The North American Banding Council 2001) implies trammel spacing should be standardized; however, it does not offer any recommendations on how to efficiently set and re-set spacing during operation. We reported our technique using marked PVC net poles in the methods section; and certainly, other options may be used. Although spacing and height need not be the same among stations, all the nets within a single station (or study) should be standardized for the greatest data utility. Whether or not they are interested in panel capture data, many stations would benefit from establishing standardization of net height and trammel spacing. Although “what looks right” may be appropriate for short-term or targeted studies, it opens the possibility of confounding effects at long-term sites when personnel and infrastructure (e.g., net poles, mist-net brand) change. As Remsen and Good (1996) demonstrated, even small changes to the height of the top trammel line of a mist-net can lead to significant changes in sampling.
Standardization protects long-term datasets from being influenced by easily overlooked variables such as the average height of the banding team or the length of conduit pole sold at the local hardware store. To illustrate the importance of standard mist-net set up, consider the following scenario: a new station manager at Coyote Creek Field Station ends our use of marked PVC poles to set mist-net height and trammel spacing. Although banders might be encouraged to use sticks from the ground, these are inherently unreliable (e.g., too short, too big, snaps with pressure, wet with morning dew, muddy after rain, etc.). Undoubtedly, our mist-nets would end up at lower absolute heights, especially as the memory of the old mist-net set up is lost as banders retire and new banders arrive. For taxa such as Willow Flycatchers, where 76% of captures occurred in the two uppermost panels (Table 2), this change may drive a detectable decline in captures. This error becomes even more problematic given Willow Flycatchers are listed as endangered in California. Thus, having standard mist-net height and trammel spacing at Coyote Creek Field Station is not just scientific best practice, but also provides conservation value.
Because panel data may be collected at all ground-level nets, the methods presented here greatly expand the accessibility of studying capture height biases. Previous studies using paired nets have operated just 3–6 net locations (Bonter et al. 2008, Tattoni and LaBarbera 2022), likely because of the cost and effort of constructing these rigs. Without easy replication it becomes difficult to test hypotheses, particularly those linked to habitat structure (e.g., does habitat succession confound population trends?). In contrast, ground-level nets are easy to set up temporarily or in permanent settings, and those that are already being operated by a banding station can provide a wealth of data. Although there are limitations to using panel data, there are also new opportunities to study mist-net methodology and avian capture patterns at station, or even multi-station, scales.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
JT and KL developed, piloted, and implemented the protocol for collecting and recording panel capture information. JT conducted the analyses with input from KL. JT wrote the manuscript with substantial contributions from KL.
ACKNOWLEDGMENTS
We are appreciative of the dozens of CCFS/SFBBO volunteers that helped maintain and collect data at our banding station. SFBBO staff including Gabbie Burns, Josh Scullen, and Dan Wenny were instrumental in piloting and establishing the mist-net panel protocol. We thank Arleen Feng, Gerry Ellis, and the other volunteers (whose names have been lost to time) who in the 1990s developed the PVC net raising pole described here. We want to acknowledge the anonymous reviewer of a previous manuscript who prompted us to look at panel capture data. Our study site is on land that was and continues to be important to the Ramaytush Ohlone people and Muwekma Ohlone Tribes.
DATA AVAILABILITY
The data and code used in this paper can be found at https://doi.org/10.5061/dryad.zw3r228f4.
LITERATURE CITED
Barton, G. G., and B. K. Sandercock. 2018. Long-term changes in the seasonal timing of landbird migration on the Pacific Flyway. Condor 120:30-46. https://doi.org/10.1650/CONDOR-17-88.1
Bonter, D. N., E. W. Brooks, and T. M. Donovan. 2008. What are we missing with only ground-level mist-nets? Using elevated nets at a migration stopover site. Journal of Field Ornithology 79:314-320. https://doi.org/10.1111/j.1557-9263.2008.00179.x
Churchwell, R. T., and G. Barton. 2006. Predation of mist net birds and an investigation of a solution. North American Bird Bander 31:115-120.
DeSante, D. F. 1992. Monitoring avian productivity and survivorship (MAPS): a sharp, rather than blunt, tool for monitoring and assessing landbird populations. Pages 511-521 in D. R. McCullough and R. H. Barrett, editors. Wildlife 2001: populations. Springer, Dordrecht, The Netherlands. https://doi.org/10.1007/978-94-011-2868-1_39
DeSante, D. F., K. M. Burton, P. Velez, D. Froehlich, D. Kaschube, and S. Albert. 2022. MAPS manual: 2022 protocol. Instructions for the establishment and operation of constant-effort bird-banding stations as part of the monitoring avian productivity and survivorship program. The Institute for Bird Populations, Petaluma, California, USA.
de Zwaan, D. R., A. Huang, Q. McCallum, K. Owen, M. Lamont, and W. Easton. 2022. Mass gain and stopover dynamics among migrating songbirds are linked to seasonal, environmental, and life-history effects. Ornithology 139:1-16. https://doi.org/10.1093/ornithology/ukac027
Dunn, E. H., and C. J. Ralph. 2004. The use of mist-nets as a tool for bird population monitoring. Studies in Avian Biology 29:1-6.
Fitzgerald, B. M., H. A. Robertson, and A. H. Whitaker. 1989. Vertical distribution of birds mist- netted in a mixed lowland forest in New Zealand. Notornis 36:311-321.
Gelman, A., J. B. Carlin, H. S. Stern, and D. B. Rubin. 2003. Bayesian data analysis. Second edition. Chapman and Hall/CRC, Boca Raton, Florida, USA. https://doi.org/10.1201/9780429258480
Gray, M., K. E. Sieving, and J. A. Cox. 2022. The nuthatch and the hare: slow explorers dominate in a re-established population of the Brown-headed Nuthatch (Sitta pusilla) two decades later. Avian Conservation and Ecology 17(1):7. https://doi.org/10.5751/ACE-02111-170107
Jenni, L., M. Leuenberger, and F. Rampazzi. 1996. Capture efficiency of mist nets with comments on their role in the assessment of passerine habitat use. Journal of Field Ornithology 67:263-274.
Joseph, L., D. B. Wolfson, and R. Du Berger. 1995. Sample size calculations for binomial proportions via highest posterior density intervals. Journal of the Royal Statistical Society. Series D (The Statistician) 44:143-154. https://doi.org/10.2307/2348439
Kellner, K. 2019. jagsUI: A wrapper around “rjags” to streamline “JAGS” analyses.
Lövei, G. L., T. Csörgõ, and G. Miklay. 2001. Capture efficiency of small birds by mist nets. Ornis Hungarica 11:19-25.
Lunn, D. J., A. Thomas, N. Best, and D. Spiegelhalter. 2000. WinBUGS - a Bayesian modelling framework: concepts, structure, and extensibility. Statistics and Computing 10:325-337. https://doi.org/10.1023/A:1008929526011
North American Banding Council. 2001. The North American banders’ study guide. North American Banding Council, Point Reyes Station, California, USA. http://www.nabanding.net/wp-content/uploads/2012/04/STUDYGUIDE1.pdf
Plummer, M. 2017. JAGS: Just Another Gibbs Sampler. https://sourceforge.net/projects/mcmc-jags/
R Core Team. 2017. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Remsen Jr., J. V., and D. A. Good. 1996. Misuse of data from mist-net captures to assess relative abundance in bird populations. Auk 113:381-398. https://doi.org/10.2307/4088905
Roche, E. A., C. R. Brown, M. B. Brown, and K. M. Lear. 2013. Recapture heterogeneity in Cliff Swallows: increased exposure to mist nets leads to net avoidance. PLoS ONE 8:e58092. https://doi.org/10.1371/journal.pone.0058092
Saracco, J. F., P. Pyle, D. R. Kaschube, M. Kohler, C. M. Godwin, and K. R. Foster. 2022. Demographic declines over time and variable responses of breeding bird populations to human footprint in the Athabasca Oil Sands Region, Alberta, Canada. Ornithological Applications 124:duac037. https://doi.org/10.1093/ornithapp/duac037
Stewart, I. T., L. Healey, K. LaBarbera, H. Li, J. C. Scullen, Y. Wang, and D. Wenny. 2022. Long term progress in riparian restoration with concurrent avian declines in the southern San Francisco Bay Area (CA). Ecological Restoration 40:203-217. https://doi.org/10.3368/er.40.3.203
Tattoni, D. J., and K. LaBarbera. 2022. Capture height biases for birds in mist-nets vary by taxon, season, and foraging guild in northern California. Journal of Field Ornithology 93(1):1. https://doi.org/10.5751/JFO-00021-930101
Vecchi, M. B., and M. A. S. Alves. 2015. Bird assemblage mist-netted in an Atlantic Forest area: a comparison between vertically-mobile and ground-level nets. Brazilian Journal of Biology 75:742-751. https://doi.org/10.1590/1519-6984.00914
Fig. 1

Fig. 1. Marked PVC poles are used at Coyote Creek Field Station, California, USA, to rapidly set and re-set a standardized mist-net height and trammel spacing. (A) A volunteer extractor stands with one such net raising pole, which may also be used as a walking stick across uneven and muddy terrain. (B) The elbow joint at the top of the PVC pole is used to raise/lower trammel lines and the black tape marks the spacing of the trammel lines.
Fig. 2
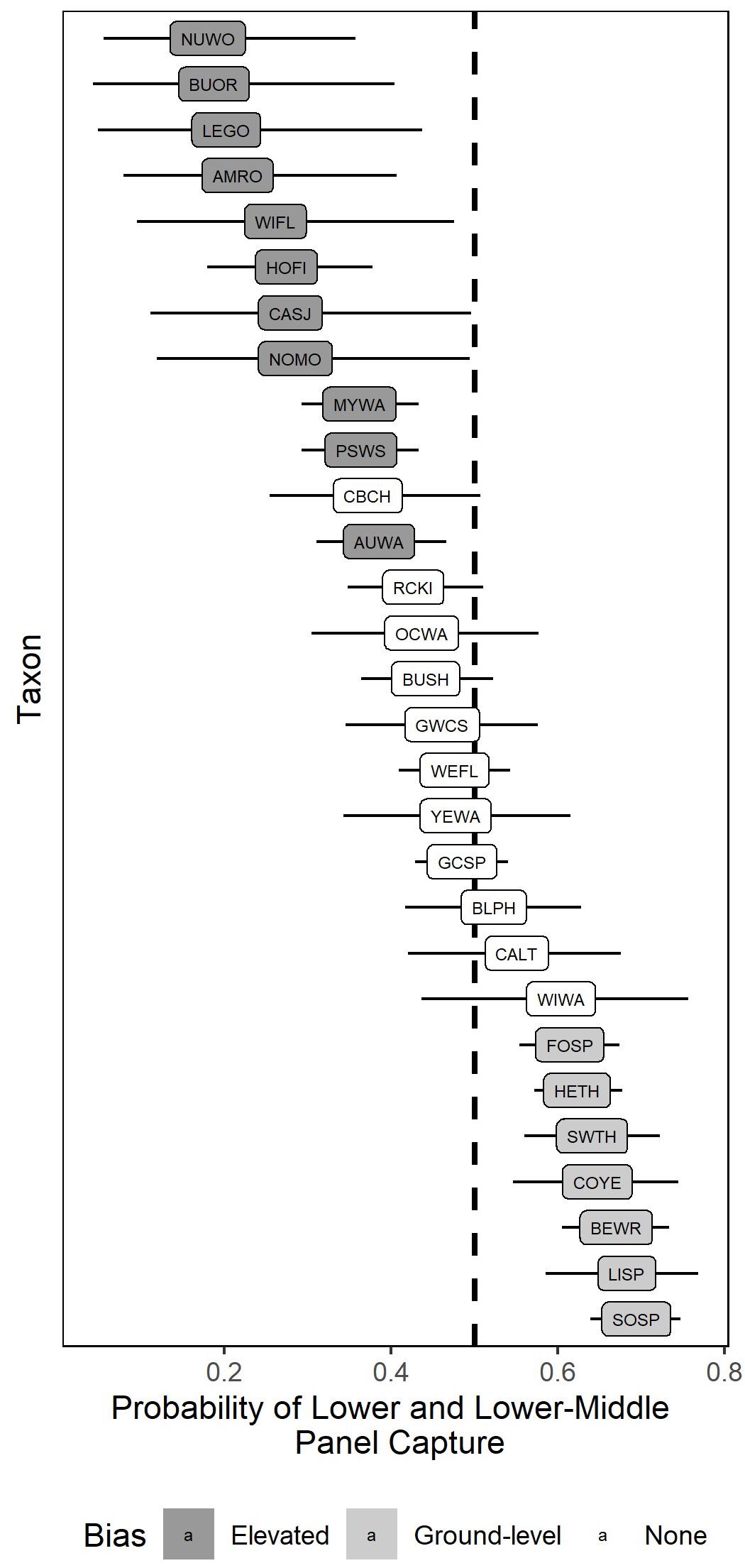
Fig. 2. Estimated probabilities of capture in the two lowest mist-net panels (lower and lower-middle) for 29 taxa at the Coyote Creek Field Station, California, USA. Black bars represent the 95% Credible Intervals (CIs). We considered taxa with 95% CIs below 0.5 to be biased toward the upper panels (marked with dark gray) and those with 95% CIs above 0.5 to be biased toward the lower panels (marked with light gray). Taxa codes used can be found in Table 2.
Fig. 3
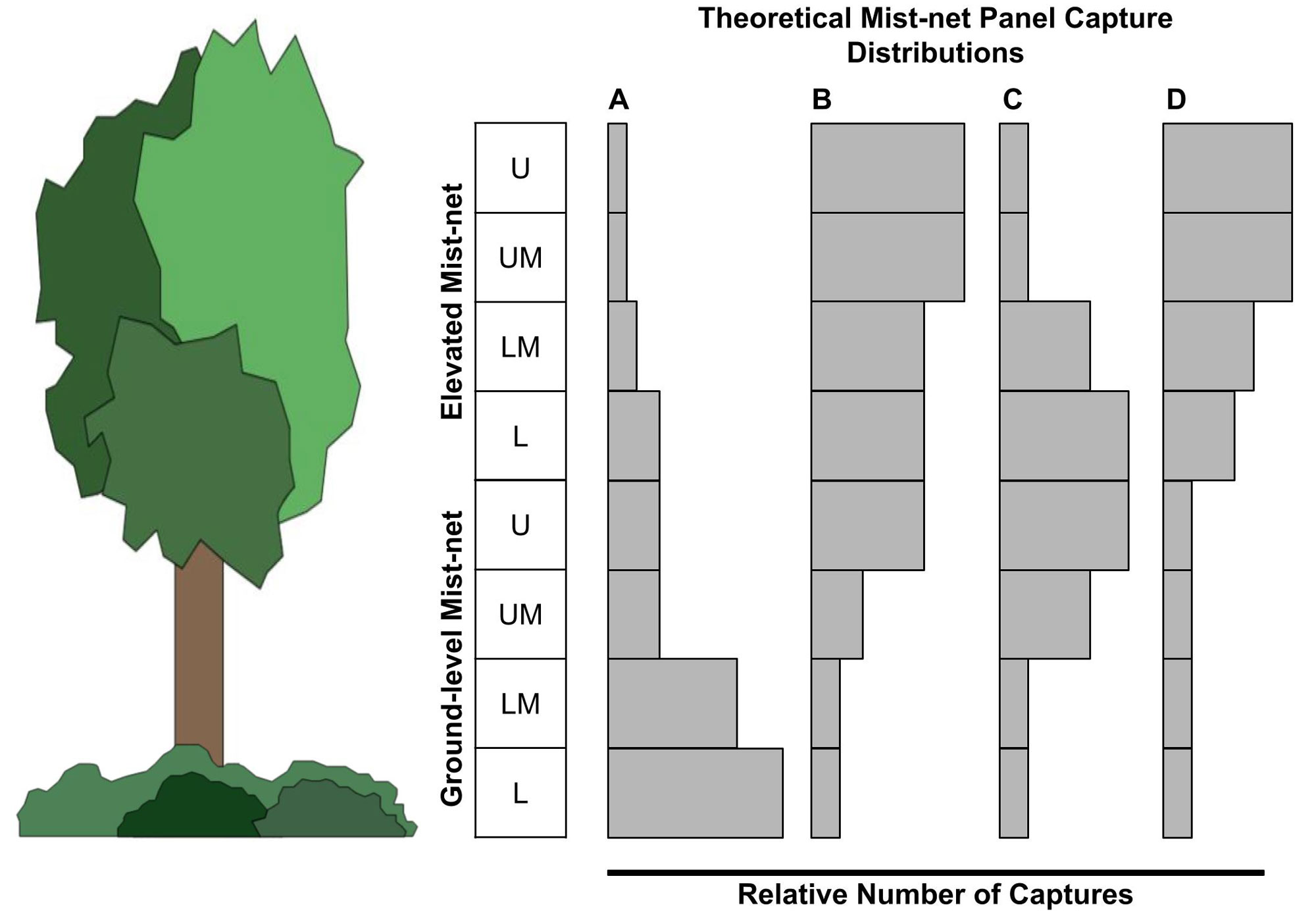
Fig. 3. Theoretical mist-net panel capture distributions that may account for discrepancies between capture height biases estimate with data from ground-level panels versus elevated mist-net rigs. The gray bars represent the relative number of captures from each mist-net panel (L = Lower, LM = Lower Middle, UM = Upper Middle, and U = Upper). These capture distributions would result in (A) ground-level bias detected in both methods, (B) elevated bias detected in both methods, (C) elevated bias using panels and no bias with elevated mist-net rigs, and (D) elevated bias using elevated mist-net rigs and no bias using panels.
Table 1
Table 1. Summary of captures across the mist-net panels by riparian woodland age at Coyote Creek Field Station, California, USA. Overstory vegetation structure is summarized by each habitat from data collected June–July 2020. Across all three woodland ages, the lower-middle and upper-middle panels captured the most birds and the lower and upper panels captured the least. See Stewart et al. (2022) for details on vegetation sampling methods.
| Overstory height (m) | Captures by mist-net panels | ||||||||
| Habitat age | Mean | SD | Lower | Lower-middle | Upper-middle | Upper | Totals | ||
| >70 years | 5.92 | 4.30 | 173 (0.23) | 248 (0.32) | 231 (0.30) | 116 (0.15) | 768 | ||
| 36 years | 7.73 | 4.31 | 199 (0.21) | 304 (0.32) | 290 (0.30) | 162 (0.17) | 955 | ||
| 30 years | 4.95 | 2.17 | 308 (0.20) | 441 (0.29) | 470 (0.31) | 287 (0.19) | 1506 | ||
| Totals | 680 (0.21) | 993 (0.31) | 991 (0.31) | 565 (0.17) | 3229 | ||||
Table 2
Table 2. From November 2020 through February 2023, we had 3229 unique captures of our 29 study taxa at Coyote Creek Field Station, California, USA. Overall captures were distributed across the four mist-net panels with 680 (21.1%) in the lower panel (L), 944 (30.8%) in the lower-middle panel (LM), 991 (30.7%) in the upper-middle panel (UM), and 565 (17.5%) in the upper panel (U). Taxon level panel capture distributions and estimates for the probability of capture in the lower and lower-middle mist-net panels are shown. Estimates below 0.5 were considered an elevated (E) bias while those above 0.5 were considered a ground-level (G) bias. We compared these biases to those obtained by Tattoni and LaBarbera (2022) using elevated mist-net rigs at the same study site.
| Taxon | Code | Sample Sizes | Probability of Capture in L and LM panels | Bias | Bias from Tattoni and LaBarbera 2022 | ||||
| L | LM | UM | U | ||||||
| Nuttall’s Woodpecker Dryobates nuttallii | NUWO | 0 | 3 | 13 | 4 | 0.18 (0.05-0.37) | E | ||
| Willow Flycatcher Empidonax traillii | WIFL | 2 | 2 | 8 | 5 | 0.26 (0.10-0.47) | E | E | |
| Western Flycatcher Empidonax difficilis/occidentalis | WEFL | 34 | 63 | 72 | 35 | 0.46 (0.41-0.55) | E | ||
| Black Phoebe Sayornis nigricans | BLPH | 15 | 30 | 24 | 17 | 0.52 (0.42-0.62) | |||
| California Scrub-Jay Aphelocoma californica | CASJ | 2 | 2 | 8 | 4 | 0.28 (0.10-0.50) | E | ||
| Chestnut-backed Chickadee Poecile rufescens | CBCH | 8 | 11 | 21 | 12 | 0.37 (0.25-0.51) | E | ||
| Bushtit Psaltriparus minimus | BUSH | 21 | 46 | 51 | 34 | 0.44 (0.37-0.52) | |||
| Ruby-crowned Kinglet Corthylio calendula | RCKI | 15 | 42 | 42 | 35 | 0.43 (0.35-0.51) | |||
| Bewick’s Wren Thryomanes bewickii | BEWR | 72 | 77 | 48 | 25 | 0.67 (0.61-0.73) | G | G | |
| Northern Mockingbird Mimus polyglottos | NOMO | 3 | 2 | 9 | 5 | 0.29 (0.12-0.49) | E | ||
| Swainson’s Thrush Catharus ustulatus | SWTH | 35 | 48 | 31 | 15 | 0.64 (0.56-0.72) | G | G | |
| Hermit Thrush Catharus guttatus | HETH | 88 | 111 | 76 | 44 | 0.62 (0.57-0.67) | G | G | |
| American Robin Turdus migratorius | AMRO | 2 | 2 | 13 | 4 | 0.22 (0.08-0.40) | E | ||
| House Finch Haemorhous mexicanus | HOFI | 7 | 12 | 28 | 24 | 0.27 (0.18-0.38) | E | ||
| Lesser Goldfinch Spinus psaltria | LEGO | 1 | 1 | 5 | 6 | 0.20 (0.04-0.43) | E | NA | |
| Fox Sparrow Passerella iliaca | FOSP | 60 | 83 | 68 | 22 | 0.61 (0.55-0.67) | G | G | |
| Gambel’s White-crowned Sparrow Zonotrichia leucophrys gambelii | GWCS | 6 | 23 | 21 | 13 | 0.46 (0.34-0.58) | |||
| Puget-sound White-crowned Sparrow Zonotrichia leucophrys pugetensis | PSWS | 19 | 43 | 61 | 48 | 0.36 (0.30-0.44) | E | ||
| Golden-crowned Sparrow Zonotrichia atricapilla | GCSP | 55 | 93 | 106 | 51 | 0.49 (0.43-0.54) | G | ||
| Song Sparrow Melospiza melodia | SOSP | 98 | 94 | 57 | 27 | 0.70 (0.64-0.75) | G | G | |
| Lincoln’s Sparrow Melospiza lincolnii | LISP | 34 | 36 | 23 | 9 | 0.68 (0.58-0.76) | G | ||
| California Towhee Melozone crissalis | CALT | 10 | 22 | 19 | 7 | 0.55 (0.43-0.67) | |||
| Bullock’s Oriole Icterus bullockii | BUOR | 0 | 2 | 9 | 3 | 0.19 (0.04-0.40) | E | ||
| Orange-crowned Warbler Leiothlypis celata | OCWA | 11 | 9 | 18 | 8 | 0.43 (0.30-0.58) | |||
| Common Yellowthroat Geothlypis trichas | COYE | 27 | 31 | 19 | 12 | 0.65 (0.54-0.74) | G | G | |
| Yellow Warbler Setophaga petechia | YEWA | 7 | 15 | 15 | 9 | 0.48 (0.33-0.62) | E | ||
| Audubon’s Yellow-rumped Warbler Setophaga coronata auduboni | AUWA | 22 | 36 | 53 | 40 | 0.39 (0.32-0.47) | E | E | |
| Myrtle Yellow-rumped Warbler Setophaga coronata coronata | MYWA | 19 | 42 | 65 | 43 | 0.36 (0.29-0.43) | E | E | |
| Wilson’s Warbler Cardellina pusilla | WIWA | 7 | 12 | 8 | 4 | 0.61 (0.44-0.76) | |||
| Totals | 680 | 993 | 991 | 565 | |||||