







The following is the established format for referencing this article:
Prytula, E. D., M. W. Reudink, S. E. LaZerte, J. Sonnleitner, and A. E. McKellar. 2023. Shifts in breeding distribution, migration timing, and migration routes of two North American swift species. Journal of Field Ornithology 94(3):14.ABSTRACT
Climate change has resulted in changes to ecosystems and weather because of earlier onset of spring weather, later onset of fall weather, and more extreme weather patterns. Migratory birds may experience challenges adjusting to these new conditions. We utilized community science data from eBird that spanned 2009–2018 to test for changes in distribution and migration of two North American swift species. We asked if Vaux’s Swifts (Chaetura vauxi) and Chimney Swifts (Chaetura pelagica) changed their breeding distribution, migration routes, timing of migration, or speed of spring and fall migration over time. Our results show that Vaux’s Swifts shifted their breeding centroid south-east and Chimney Swifts shifted their breeding centroid west. There was also a shift in Vaux’s Swifts migration route to the east, almost proportionate in magnitude to its eastern shift in breeding range. Vaux’s Swifts displayed an advance in their start of spring migration, and Chimney Swifts exhibited a delay in their start of fall migration. These responses may be due to earlier onset of spring and a possible delay of colder temperatures associated with the onset of fall conditions. Our results indicated that both species are breeding further away from the coastline and more toward central North America, and suggest that swifts may display some phenotypic plasticity in response to changing environmental conditions. What remains unclear is if this phenotypic plasticity will be enough to prevent further population loss in the two species of swift, in the face of ongoing climate change.
RESUMEN
El cambio climático ha resultado en cambios en los ecosistemas y el clima debido a un inicio más temprano de la primavera, un inicio más tardío del otoño y eventos climáticos más extremos. Las aves migratorias pueden experimentar retos para ajustarse a estas nuevas condiciones. Utilizamos datos de ciencia comunitaria de eBird entre 2009-2018 para evaluar los cambios en la distribución y migración de dos especies de vencejos Norte Americanos. Nos preguntamos si Chaetura vauxi y Chaetura pelagica han cambiado su distribución reproductiva, rutas de migración y tiempo de la migración, o la velocidad de la migración de primavera y otoño a través del tiempo. Nuestros resultados muestran que Chaetura vauxi ha movido el centroide de reproducción hacia el sureste y Chaetura pelagica ha movido su centroide de reproducción hacia el oeste. La ruta de migración de Chaetura vauxi también se ha desplazado hacia el este, casi proporcionalmente a la magnitud del cambio hacia el este de su rango de reproducción. Chaetura vauxi mostró un adelantamiento en el inicio de la migración de primavera, y Chaetura pelagica mostró un retraso en el inicio de su migración de otoño. Estas respuestas pueden deberse a un inicio más temprano de la primavera y posiblemente a un retraso en las temperaturas frías asociadas con el inicio de las condiciones de otoño. Nuestros resultados sugieren que ambas especies se están reproduciendo más lejos de la línea de costa y más hacia Norte América central, y sugieren que los vencejos pueden mostrar alguna plasticidad fenotípica en respuesta a las condiciones ambientales cambiantes. Todavía no tenemos claro si esta plasticidad fenotípica será suficiente para prevenir perdidas poblacionales futuras en estas dos especies de vencejos, con miras al cambio climático actual.
INTRODUCTION
For migratory birds, migration timing and routes have evolved through a series of trade-offs, balancing a suite of selective pressures that favor early arrival to the breeding grounds to maximize reproductive success (Cristol 1995), while synchronizing food availability to peak nestling resource demands, avoiding predation during migration, and avoiding severe weather conditions during migration and on the breeding grounds (Häfker et al. 2017, Reid et al. 2018). Arriving too early on the breeding grounds can result in encountering inhospitable conditions that may lower survival rates (Newton 2007), and arriving too late can result in a poorer territory, missed peak resource abundance, and time constraints on rearing young (Gienapp and Bregnballe 2012). The timing of migration is dictated by both internal endogenous cues and external environmental cues (Berthold 1996). What remains unclear, however, is how migratory birds will respond to a rapidly changing climate. Changes in global temperatures are predicted to increase the frequency and severity of droughts, floods, and sea level rise, and will alter plant and insect phenology (Schwartz et al. 2006, Malhi et al. 2020). With a change in temperature, we can expect a change in weather patterns, an advancement of spring phenology, and changes to fall phenology (Schwartz et al. 2006). As a result, the timing of resource availability during each phase of a bird’s annual cycle may be altered (Parmesan 2007), as will the weather conditions experienced during migration. The rapid change to the tightly co-evolved relationship between migration timing and resource availability during migration may result in temporal mismatches, resulting in reduced survival and reproduction.
Different species of birds will respond to environmental cues with different changes to migratory routes, timing, and speed based on their different life history traits (Knudsen et al. 2011). Some species of birds appear to be altering their timing of migration in response to changes in temperature, wind, and precipitation (Studds and Mara 2011, Hurlbert and Liang 2012, Prytula et al. 2021). In response to differences in environmental cues as well as fueling requirements, birds may change migratory routes (Supp et al. 2015), the speed at which they migrate, daily flight duration, or the amount of time spent at stopover sites (Nilsson et al. 2013). Climate change may also result in changes to breeding distributions, as northern latitudes and higher elevations could become more favorable for breeding (Hitch and Leberg 2007). Because of endogenous rhythms constraining changes in migration timing, some species may respond to environmental cues by choosing different migratory routes but maintaining migratory timing (Alerstam et al. 2006, Vardanis et al. 2011). La Sorte et al. (2013) found migrants in the western flyway of North America chose migratory routes with higher primary production and greener conditions in the spring and less productive and potentially more direct routes in the fall, while migrants in the eastern flyway of North America chose areas with maximum greenness for both spring and fall.
The ability to modify migratory timing can have important consequences, as Møller et al. (2008) showed that European bird species that did not advance their migration timing were more likely to be in decline. Some studies have found that long-distance migrants may have greater difficulty in adjusting their migration timing compared to short-distance migrants, which could potentially leave them vulnerable to climate change (Knudsen et al. 2011, Fraser et al. 2013, Lehikoinen et al. 2019). However, results are inconsistent because some long-distance migrants have advanced their spring arrival more than short distance migrants (Both et al. 2005, Jonzén et al. 2006, Knudsen et al. 2011). One reason long-distance migrants may have difficulty altering their migratory timing in response to climate change may be because they are less able to detect changes in the earlier onset of spring from their wintering grounds (Visser et al. 2009), and instead rely on endogenous rhythms to time migration (Both and Visser 2001). Long-distance migrants may also be constrained in their ability to change their speed of migration and/or departure date from the wintering grounds, resulting in a mismatch between arrival on the breeding grounds and optimal breeding conditions (Fraser et al. 2013). Finally, seasonality is likely to play a role in migratory birds’ responses to climate change. Birds are generally under greater time constraints to arrive early in spring to establish high quality territories and find the best mates (Cristol 1995), and thus may be more likely to advance their spring migration than fall migration (Prytula et al. 2021). During fall migration, they may be more responsive to weather variables experienced en route to the wintering grounds (La Sorte et al. 2013, Prytula et al. 2021).
In the past, studies of migratory timing and migration routes often utilized bird banding, but data was dependent on recaptures, resulting in limited sample sizes and biases due to variation in recovery effort across the range (Thorup and Conn 2009). More recently, tracking devices such as satellite trackers, GPS loggers, and light-level geolocators have been used to track individual birds, but such studies can be limited by cost, sample size, weight of devices, and coverage of different populations (Bridge et al. 2011). Alternatively, large-scale community science databases provide an unparalleled resource for detecting changes in migration and distribution over broad spatial and temporal scales (Heim et al. 2020). These databases allow for the study of species across their entire range over multiple years, and in locations not always accessible to scientists (Dickinson et al. 2010). In this study, we make use of eBird to analyze changes to the migration and distribution of two long-distance migratory swift species in North America, the Vaux’s Swift (Chaetura vauxi) and the Chimney Swift (Chaetura pelagica).
Vaux’s Swifts and Chimney Swifts are both long-distance migratory aerial insectivores that migrate in flocks of hundreds to thousands, using abandoned chimneys and hollow old-growth trees as migratory roosts (Steeves et al. 2020, Schwitters et al. 2021). Chimney Swifts overwinter in South America, migrate through eastern Central America, and breed from Texas to as far north as southeastern Canada (Steeves et al. 2020). Vaux’s Swifts are a slightly smaller counterpart to the Chimney Swift; they overwinter in Mexico and Central America, migrate through the west coast of North America, and breed from California to the Yukon (Schwitters et al. 2021). Both swift species have declining populations that are likely caused by multiple drivers: they are long distance migrants, have declining food abundance due to a decline in aerial insects, and have specific roosting needs that could be subject to habitat loss (Spiller and Dettmers 2019).
We analyzed temporal trends in breeding season latitude and longitude, as well as spring and fall migration (referred to as “migratory seasons”) routes, timing, and speed over a 10-year period. We predicted that both species’ breeding distributions would shift northward over time, as conditions at higher latitudes are becoming more temperate in the northern hemisphere (Hitch and Leberg 2007). In addition, based on previous work by Prytula et al. (2021), we predicted that both species would begin spring migration earlier over time.
METHODS
All statistical analyses were performed in R (v 4.0.2, R Core Team 2022). Presence data for Vaux’s Swifts and Chimney Swifts from 2009 to 2018 were gathered from the community science database eBird (eBird 2021). The eBird database includes information on whether observations were incidental or collected while travelling or stationary. Observation effort is also recorded, including the length of observation, distance travelled while observing, and how many observers were included. Finally, if all birds observed were identified and recorded, the checklist is considered “complete,” which allows for the “zero-fill” of data for locations where species were not present. We used the auk package (v 0.4.0; Strimas-Mackey et al. 2018) to filter the data to include checklists that met the eBird criteria of best practices. Specifically, we included lists that were “Travelling” or “Stationary” and excluded “Incidental” and “Historical” data. We also only included lists where observations lasted for a duration between 0–5 hr, observers travelled a distance between 0–5 km, and records were considered “complete” (Strimas-Mackey et al. 2020). We subsequently added counts of 0 to the checklists that did not include Vaux’s or Chimney Swift observations to “zero-fill” the data. Sample sizes of checklists for each species for each year can be found in Appendix 1.
We used the methodology developed by Supp et al. (2015) and subsequently used by Sonnleitner et al. (2022), to compile presence data for Vaux’s and Chimney Swifts from 2009 to 2018 and generate equal-area icosahedron hex grids (23,323 km²) based on an icosahedron map overlaying North America using the dggridR package (v 2.0.3; Barnes 2018). We summarized daily presence for each hex cell as the total number of binned checklists with a swift present in the hex cell divided by total number of checklists for each date (Fig. 1; sample sizes of checklists found in Appendix 1). The weighted daily mean location was then calculated using the central longitude and latitude of each cell to measure spatial variation in presence data over time. To track migration on an annual basis, we used generalized additive models (GAM; package mgcv; Wood 2011) to model these mean locations for each species and produce smoothed paths of migration (Fig. 1). From these smoothed paths we extracted population-level occurrence centroids (hereafter referred to as GAM-predicted occurrence centroids) with latitude and longitude for each species for each day of the year. Data for Chimney Swifts overwintering in South America was too sparse to accurately generate GMAs; thus, we omitted ordinal dates of < 75 and > 300 for this species prior to modeling.
Breeding/migration distributions
For the breeding season, the maximum latitude and median longitude were calculated for the population GAM-predicted occurrence centroids between 23 June and 12 August (ordinal dates 175–225) for each species each year. For spring and fall migrations, the median longitude was calculated during each migratory season (between the start and end of each migration as calculated in "Migration timing” below). Median longitudinal values were once again used to minimize the effects of potential outliers, or edge cases when the population was near breeding or non-breeding grounds.
Migration timing
We defined the start and end of migration (seasons of the annual cycle) based upon whether the GAM-predicted occurrence centroid had reached a stable latitude. At its minimum latitude, the species was considered to be on its wintering grounds, and at its maximum latitude, the species was considered to be on its breeding grounds. Based on methodology from Supp et al. (2015), we used a threshold approach to determine the date a species left this stable latitude (start of migration) and when they arrived (end of migration). We established a latitudinal threshold for each species, year, and season (i.e., Vaux’s Swift, 2009, fall migration) based on the 99% confidence band of predicted daily locations. Southern latitudinal thresholds were calculated as the minimum latitude of the upper confidence limit, whereas northern latitudinal thresholds were calculated as the maximum latitude of the lower confidence limit. In each calculation we limited dates to those likely to include migration. Specifically, we considered the start of spring migration to be within the period Jan–March 20 (ordinal date 1–80), the end of fall migration to be within the period 11 Oct–10 Dec (285–345), the end of spring migration to be within the period 20 March–23 June (80–175), and the start of fall migration to be within the period 12 Aug–11 Oct (225–285).
The dates on which the GAM-predicted occurrence centroids crossed these latitudinal thresholds gave us coarse estimates of the start and end dates of migration for each species, year, and season. Using these dates as a starting point we then used segmented regression to calculate more precise break points in latitude and ordinal dates that more accurately reflected the start and end of migration for spring and fall (segmented package v1.0-0; Muggeo 2008; cf Supp et al. 2015). Because the data for Chimney Swifts overwintering in South America was too sparse to accurately generate GAMs, we were unable to generate values for southern latitudes, including the start of spring migration and end of fall migration for this species.
Migration speed
Maximum population-level daily speed of migration was calculated as the median kilometers per day over the five fastest days (km/day) of GAM-predicted occurrence centroid movements for each season (Supp et al. 2015). We chose the median speed to minimize the effect of potential outliers. We searched for the fastest spring migration dates between 1 January–23 June (ordinal dates 1–175) and the fastest fall migration dates from 12 August–5 December (225–340). To ensure the speeds at the start and end of migration were not excluded, date ranges extended into breeding and overwintering seasons. Because there was little movement during the non-breeding period, we did not expect that this would alter our calculations of maximum migration speed.
Examining changes over time
We constructed a series of linear models to examine changes over time in maximum breeding latitude and median breeding longitude, median migratory longitude for both migratory seasons, the start and end of migration for both seasons, and maximum daily migration speed for both season. We ran models separately for each species and included year as a main effect in all models. An alpha value of 0.05 was used to determine significance. The DHARMa package (v 0.4.3; Hartig 2021) was used to assess model assumptions and fit. Results are presented as model estimates when slope interpretation was required, or as type III ANOVAs (car package v 3.0-8; Fox and Weisberg 2019).
RESULTS
Breeding/migration distributions
Breeding latitude
We found that the maximum breeding latitude for Vaux’s Swifts shifted 1.20 degrees (133.2 km) south per year from 2009 to 2018 (r² = 0.89, P < 0.001, df = 9), while Chimney Swifts did not change their maximum breeding latitude over time (r² = 0.0005, P = 0.35, df = 9; Table 1, Fig. 2).
Breeding longitude
When examining changes to the median longitude of the GAM-predicted occurrence centroid during breeding, we found that Vaux’s Swifts shifted the median longitude east 1.44 degrees (159.84 km) per year from 2009 to 2018 (r² P < 0.001, df = 9), while Chimney Swifts shifted their median longitude 0.24 degrees (26.64 km) west per year (r² = 0.80, P < 0.001, df = 9; Table 1, Fig. 2).
Spring migration longitude
Vaux’s Swifts shifted their median spring migration longitude 1.12 degrees (124.32 km) east per year from 2009 to 2018 (r² = 0.84, P< 0.001, df = 9), while Chimney Swifts did not show a significant change in median longitude over time (r² = 0.015, P = 0.32, df = 9; Table 1, Fig. 2).
Fall migration longitude
Similar to spring migration, Vaux’s Swifts shifted their fall median longitude 1.46 degrees (162.06) east per year from 2009 to 2018 (r² = 0.35, P = 0.04, df = 9), while Chimney Swifts did not change over time (r² = -0.087, P = 0.61, df = 9; Table 1, Fig. 2).
Migration timing
Spring
Because we did not have sufficient data from eBird for Chimney Swifts to calculate the start of spring migration, we only examined potential changes to Vaux’s Swift start of spring migration. The start of spring migration advanced 1.35 days per year from 2009 to 2018 for Vaux’s Swifts (r² = -0.77, P = 0.06, df = 9; Table 1). We had sufficient data for both species to calculate end of spring migration. We found a marginal increase to the end of spring migration for Vaux’s Swifts (r² = 0.30, P < 0.001, df = 9), but no change over time for Chimney Swifts (r² = -0.12, P = 0.89, df = 9; Table 1).
Fall
We found no change over time for the start of fall migration for Vaux’s Swifts (r² = 0.074, P = 0.23, df = 9), but we found that Chimney Swifts delayed their start of fall migration 4.2 days per year from 2009 to 2018 (r² = 0.38, P = 0.035, df = 9; Table 1). Because we did not have sufficient data for Chimney Swifts to calculate the end of fall migration, we ran a linear model for only Vaux’s Swifts to examine change over time. The resulting model indicated that the end of fall migration was delayed 1.64 days per year from 2009 to 2018, though this pattern was not significant (r² = 0.30, P = 0.06, df = 9; Table 1).
Migration speed
Spring
Vaux’s Swifts (mean ± SD: 102.7 ± 32 km/day) appeared to migrate more quickly during spring migration than Chimney Swifts (49.6 ± 24.5 km/day; F = 17.43, P < 0.001); however, we found no change in migration speed over time for either Vaux’s Swifts (r² = 0.07, P = 0.24, df = 9) or Chimney Swifts (r² = 0.12, P = 0.18, df = 9; Table 1).
Fall
We found no difference in fall migration speed between species (F = 0.16, P = 0.70). In addition we found no change over time in the speed of fall migration for Vaux’s Swifts (r² = 0.03, P = 0.29, df = 9) or Chimney Swifts (r² = 0.07, P = 0.23, df = 9).
DISCUSSION
In this study, we used a 10-year community science dataset spanning from 2009 to 2018 to analyze population level changes in breeding grounds distribution, as well as migration routes, timing, and speed for Vaux’s Swifts and Chimney Swifts. Although both species shifted their breeding GAM-predicted occurrence centroid, it was not in the expected northern direction; instead, Vaux’s Swifts shifted to the southeast and Chimney Swifts shifted to the west. Although we did not predict a change in migration routes, Vaux’s swifts shifted their migratory routes east in both spring and fall. In contrast, Chimney Swifts did not display a change in migratory routes, at least in the northern section of their routes, given we were not able to model patterns in the south. We also found support for our prediction that Vaux’s Swifts departed on spring migration earlier, while Chimney Swifts left their breeding grounds later over time. We found no change in speed of migration for both species of swifts.
There are many documented changes in the breeding ranges of avian species in response to climate change (Thomas and Lennon 1999, Hitch and Leberg 2007, Potvin et al. 2016), often with a northern range expansion accompanied by a southern range contraction (Hitch and Leberg 2007, Rushing et al. 2020), and sometimes a southern range expansion (Rushing et al. 2020). Hovick et al. (2016) documented a southern shift in breeding range for 24% of 277 species, possibly because of uneven warming within North America, adaptation to new climates (Parmesan 2006), habitat modification (Archaux 2004), or inter species interactions (Lenoir et al. 2010). In contrast, the southern breeding GAM-predicted occurrence centroid shift for Vaux’s Swifts in our analysis is consistent with historical findings that there are aerial insectivore declines in northern North America, and increased populations in their southern range (Nebel et al. 2010). For Vaux’s Swifts, the southern shift by 1.2 degrees per year could result in reduced energy expenditure by individuals because of a shorter migration distance (Nebel et al. 2010). However, because distributions on the breeding grounds and wintering grounds can shift independently of one another and under different environmental conditions (Curley et al. 2020), and because we only examined breeding GAM-predicted occurrence centroid shifts of the entire population, we cannot say whether there have been shifts to overall migration distance experienced by individuals.
The breeding GAM-predicted occurrence centroid and migration route shift to the east for Vaux’s Swifts and the breeding GAM-predicted occurrence centroid shift to the west for Chimney Swifts mirror a recent finding by Sonnleitner et al. (2022), who showed that the migration routes of Eastern (Sialia sialis) and Western Bluebirds (Sialia mexicana) have shifted away from the coastline toward central North America. They suggested this may be a result of human encroachment on the coasts and associated habitat loss. Increases in human population density could possibly decrease swift prey availability because of increases in agriculture, and therefore pesticides (Nocera et al. 2012, Møller et al. 2021), as well as vehicle traffic (Martin et al. 2018). However, increasing coastal population density per se is probably unlikely to have had a substantial effect during the relatively short time span of our study. A loss of old growth trees and chimney roosts could also be associated with changes to migration routes and breeding distributions for Vaux’s Swifts (Bull 2003), though this is unlikely to be the case for Chimney Swifts because nestable chimneys do not seem to a limiting factor for Chimney Swift populations (Fitzgerald et al. 2014). However, with suitable chimneys being capped at approximately 5% per year, nest site availability may soon become a limiting factor (Kearney-McGee 2012, COSEWIC 2018). The eastern shift in the Vaux’s Swift migration route appears to be correlated with an eastern shift in their breeding range. With climate change generating asynchronous changes in insect abundance and phenology across latitudes and longitudes (Parmesan 2006, Aukema et al. 2008) parts of the breeding grounds could potentially become more or less productive, resulting in changes in migration route to arrive at more productive breeding grounds.
Vaux’s Swifts left their wintering grounds earlier over time and exhibited a trend to arrive back at their wintering grounds later over time. A recent analysis of Vaux’s Swift migration patterns using a separate community science database (Vaux’s Happening, https://www.vauxhappening.org/) found that first arrivals and peak roost occupancy advanced from 2008 to 2017 during spring migration (Prytula et al. 2021), which aligns with our finding that Vaux’s Swifts are leaving wintering grounds earlier over time. Chimney Swifts left their breeding grounds later over time, possibly as a result of environmental changes on the breeding grounds. Leaving the wintering grounds earlier may have been due to a change in environmental conditions on the wintering grounds, which has been shown to be a factor in other long distance migratory species (Haest et al. 2020). Arriving at wintering grounds later could be a response to increasingly harsh environmental conditions during fall migration (Jenni and Kery 2003, Prytula et al. 2021).
Previous work on Vaux’s Swift migration using community science data at migratory roost sites indicated that migratory timing was influenced by both temperature and wind (Prytula et al. 2021). Specifically, higher temperatures were associated with later fall migration and higher winds were associated with earlier spring and fall migration. As such, changes to local weather conditions resulting from climate change or large-scale climatic oscillations could result in changes to both migratory routes and the timing of migration at a large-scale. Future work that models the effects of both range-wide weather conditions and land-use changes over time will thus be critical for understanding the underlying drivers of the patterns we observed in this study.
Although there are advantages to working with population-level community science data that may not be possible based on tracking of individual birds via banding or tracking devices, such datasets present their own difficulties in terms of data reliability and generating data sets that are not spatially or temporally biased (Dickinson et al. 2010). Specifically, eBird data can include spatial bias, variation in observer effort, and a bias in species reporting (Johnston et al. 2021). Swifts are not always easy to detect and changes in observer spatial patterns or skill levels over the 10-year data period could result in relatively fewer swift detections in particular areas. Although we used several best practices techniques to mitigate these problems (e.g., spatial binning by hex grids, filtering checklists by observer effort, weighting observations by effort; Strimas-Mackey et al. 2020), it is possible that observer effects may contribute to some of the patterns observed, particularly if there were a large increase in the number of inexperienced observers in coastal areas, thereby neutralizing the observations from those regions with false negatives. However, we find it unlikely that such changes in observer behavior could be so pronounced as to completely explain the results here.
In conclusion, we documented a change in migratory timing, migration route, and breeding GAM-predicted occurrence centroid for Vaux’s Swifts, and a change in breeding GAM-predicted occurrence centroid, and start of fall migration for Chimney Swifts over a 10-year period. These changes suggest the existence of phenotypic plasticity in at least some migration-related traits, although mortality of individuals that exhibit certain traits (i.e., evolutionary change) cannot be ruled out. Though swifts may be influenced by long-term climatic cycles (e.g., ENSO), given the short period of our study, our findings may demonstrate shifts resulting from recent changes in temperature and precipitation. For example, within Canada, northern regions experienced the most drastic changes (2.3ºC from 1948 to 2016), suggesting that plant and animal phenology in northern regions may have experienced concurrent shifts (Zhang et al. 2019). Similarly, changes to precipitation regimes across North America have been drastic, especially in the west, which has experienced an increased frequency of drought over the past half century, with especially dry conditions over the past decade (Zhang et al. 2021).
Future studies could make use of individual tracking technologies such as GPS, geolocators, or banding data in combination with community science to confirm whether individuals are changing their behavior, as well as to create more accurate migratory route information (Heim et al. 2020). Future work could also examine changes in the wintering grounds of Vaux’s Swifts and Chimney Swifts over time because some species have shifted their wintering grounds by moving toward areas that were formerly too cold (Chamorro et al. 2019). Because swifts are long distance migrants, they may winter closer to their breeding grounds as the climate changes (Visser et al. 2009).
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
Funding was provided by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant to M.W.R. (4603-2018), and an Irving K. Barber BC Indigenous Student Award, New Relationship Trust Scholarship, Indspire, and the Ken Lepin Award to E.P.
DATA AVAILABILITY
Code and instructions to reproduce the analysis can be found at https://doi.org/10.5281/zenodo.7998801.
LITERATURE CITED
Alerstam, T., M. Hake, and N. Kjellén. 2006. Temporal and spatial patterns of repeated migratory journeys by ospreys. Animal Behaviour 71:555-566. https://doi.org/10.1016/j.anbehav.2005.05.016
Archaux, F. 2004. Breeding upwards when climate is becoming warmer: no bird response in the French Alps. Ibis 146:138-144. https://doi.org/10.1111/j.1474-919X.2004.00246.x
Aukema, B. H., A. L. Carroll, Y. Zheng, J. Zhu, K. F. Raffa, R. D. Moore, K. Stahl, and S. W. Taylor. 2008. Movements of outbreak populations of mountain pine beetle: influences of spatiotemporal patterns and climate. Ecography 31:348-358. https://doi.org/10.1111/j.0906-7590.2007.05453.x
Barnes, R. 2018. dggridR: Discrete global grids. R package version 2.0.3. https://CRAN.R-project.org/package=dggridR
Berthold, P. 1996. Control of bird migration. Chapman & Hall, London, UK.
Both, C., R. G. Bijlsma, and M. E. Visser. 2005. Climatic effects on spring migration and breeding in a long-distance migrant, the Pied Flycatcher Ficedula hypoleuca. Journal of Avian Biology 36:368-373. https://doi.org/10.1111/j.0908-8857.2005.03484.x
Both, C., and M. E. Visser. 2001. Adjustment to climate change is constrained by arrival date in a long-distance migrant bird. Nature 411:296-298. https://doi.org/10.1038/35077063
Bridge, E. S., K. Thorup, M. S. Bowlin, P. B. Chilson, R. H. Diehl, R. W. Fléron, P. Hartl, R. Kays, J. F. Kelly, W. D. Robinson and M. Wikelski. 2011. Technology on the move: recent and forthcoming innovations for tracking migratory birds. BioScience 61:689-698. https://doi.org/10.1525/bio.2011.61.9.7
Bull, E. L. 2003. Declines in the breeding population of Vaux’s Swifts in Northeastern Oregon. Western Birds 34:230-234.
Chamorro, D., I. Nieto, R. Real, and A. R. Muñoz. 2019. Wintering areas on the move in the face of warmer winters. Ornis Fennica 96:41-54.
Committee on the Status of Endangered Wildlife in Canada (COSEWIC). 2018. COSEWIC assessment and status report on the Chimney Swift (Chaetura pelagica) in Canada. Species at Risk Public Registry, COSEWIC, Ottawa, Ontario, Canada. https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/cosewic-assessments-status-reports/chimney-swift-2018.html
Cristol, D. A. 1995. Early arrival, initiation of nesting, and social status: an experimental study of breeding female Red-winged Blackbirds. Behavioral Ecology 6:87-93. https://doi.org/10.1093/beheco/6.1.87
Curley, S. R., L. L. Manne, and R. R. Veit. 2020. Differential winter and breeding range shifts: implications for avian migration distances. Diversity and Distributions 26:415-425. https://doi.org/10.1111/ddi.13036
Dickinson, J. L., B. Zuckerberg, and D. N. Bonter. 2010. Citizen science as an ecological research tool: challenges and benefits. Annual Review of Ecology, Evolution, and Systematics 41:149-172. https://doi.org/10.1146/annurev-ecolsys-102209-144636
eBird. 2021. eBird: An online database of bird distribution and abundance [web application]. eBird, Cornell Lab of Ornithology, Ithaca, New York, USA.
Fitzgerald, T. M., E. van Stam, J. J. Nocera, and D. S. Badzinski. 2014. Loss of nesting sites is not a primary factor limiting northern Chimney Swift populations. Population Ecology 56:507-512. https://doi.org/10.1007/s10144-014-0433-6
Fox, J., and S. Weisberg. 2019. car: Companion to applied regression. R package version 3.0-8. https://CRAN.R-project.org/package=car
Fraser, K. C., C. Silverio, P. Kramer, N. Mickle, R. Aeppli, and B. J. M. Stutchbury. 2013. A trans-hemispheric migratory songbird does not advance spring schedules or increase migration rate in response to record-setting temperatures at breeding sites. PLoS ONE 8(5):e64587. https://doi.org/10.1371/journal.pone.0064587
Gienapp, P., and T. Bregnballe. 2012. Fitness consequences of timing of migration and breeding in cormorants. PLoS ONE 7:e46165. https://doi.org/10.1371/journal.pone.0046165
Haest, B., O. Hüppop, and F. Bairlein. 2020. Weather at the winter and stopover areas determines spring migration onset, progress and advancements in Afro-Palearctic migrant birds. Proceedings of the National Academy of Sciences USA 117:17056-17062. https://doi.org/10.1073/pnas.1920448117
Häfker, N. S., B. Meyer, K. S. Last, D. W. Pond, L. Hüppe, and M. Teschke. 2017. Circadian clock involvement in zooplankton diel vertical migration. Current Biology 27:2194-2201. https://doi.org/10.1016/j.cub.2017.06.025
Hartig, F. 2021. DHARMa: residual diagnostics for hierarchical (multi-level / mixed) regression models. R package version 0.4.3. https://CRAN.R-project.org/package=DHARMa
Heim, W., R. J. Heim, I. Beermann, O. A. Burkovskiy, Y. Gerasimov, P. Ktitorov, K. Ozaki, I. Panov, M. M. Sander, S. Sjöberg, S. M . Smirenski, A. Thomas, A. Tøttrup, I. M. Tyunov, M. Willemoes, N. Hölzel, K. Thorup, and J. Kamp. 2020. Using geolocator tracking data and ringing archives to validate citizen-science based seasonal predictions of bird distribution in a data-poor region, Global Ecology and Conservation 24:e01215. https://doi.org/10.1016/j.gecco.2020.e01215
Hitch, A. T., and P. L. Leberg. 2007. Breeding distributions of North American bird species moving north as a result of climate change. Conservation Biology 21(2):534-539. https://doi.org/10.1111/j.1523-1739.2006.00609.x
Hovick, T. J., B. W. Allred, D. A. McGranahan, M. W. Palmer, R. D. Elmore, and S. D. Fuhlendorf. 2016. Informing conservation by identifying range shift patterns across breeding habitats and migration strategies. Biodiversity Conservation 25:345-356. https://doi.org/10.1007/s10531-016-1053-6
Hurlbert, A. H., and Z. Liang. 2012. Spatiotemporal variation in avian migration phenology: citizen science reveals effects of climate change. PLoS ONE 7(2):e31662. https://doi.org/10.1371/journal.pone.0031662
Jenni, L., and M. Kéry. 2003. Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proceedings of the Royal Society B 270:1467-1471. https://doi.org/10.1098/rspb.2003.2394
Johnston, A., W. M. Hochachka, M. E. Strimas-Mackey, V. Ruiz Gutierrez, O. J. Robinson, E. T. Miller, T. Auer, S. T. Kelling, and D. Fink. 2021. Analytical guidelines to increase the value of community science data: an example using eBird data to estimate species distributions. Diversity and Distributions 27(7):1265-1277. https://doi.org/10.1111/ddi.13271
Jonzén, N., A. Lindén, T. Ergon, E. Knudsen, J. O. Vik, D. Rubolini, D. Piacentini, C. Brinch, F. Spina, L. Karlsson, M. Stervander, A. Andersson, J. Waldenström, A. Lehikoinen, E. Edvardsen, R. Solvang, and N. C. Stenseth. 2006. Rapid advance of spring arrival dates in long-distance migratory birds. Science 312:1959-1961. https://doi.org/10.1126/science.1126119
Kearney-McGee, S. 2012. State Wildlife Grants Program interim performance report (T2-1-R-2): Inventory and assessment of Connecticut’s priority species. State of Connecticut Wildlife Division, Hartford, Connecticut, USA.
Knudsen, E., A. Lindén, C. Both, N. Jonzén, F. Pulido, N. Saino, W. J. Sutherland, L. A. Bach, T. Coppack, T. Ergon, P. Gienapp, J. A. Gill, O. Gordo, A. Hedenström, E. Lehikoinen, P. P. Marra, A. P. Møller, A. L. Nilsson, G. Péron, E. Ranta, D. Rubolini, T. H. Sparks, F. Spina, C. E. Studds, S. A. Saether, P. Tryjanowski, and N. C. Stenseth. 2011. Challenging claims in the study of migratory birds and climate change. Biological Reviews 86(4):928-946. https://doi.org/10.1111/j.1469-185X.2011.00179.x
La Sorte, F. A., D. Fink, W. M. Hochachka, J. P. DeLong, and S. Kelling. 2013. Population-level scaling of avian migration speed with body size and migration distance for powered fliers. Ecology 94:1839-1847. https://doi.org/10.1890/12-1768.1
Lehikoinen, A., A. Lindén, M. Karlsson, A. Andersson, T. L. Crewe, E. H. Dunn, G. Gregory, L. Karlsson, V. Kristiansen, S. Mackenzie, S. Newman, J. E. Røer, C. Sharpe, L. V. Sokolov, Å. Steinholtz, M. Stervander, I. S. Tirri, and R. S. Tjørnløv. 2019. Phenology of the avian spring migratory passage in Europe and North America: asymmetric advancement in time and increase in duration, Ecological Indicators 101:985-991. https://doi.org/10.1016/j.ecolind.2019.01.083
Lenoir, J., J.-C. Gégout, A. Guisan, P. Vittoz, T. Wohlgemuth, N. E. Zimmermann, S. Dullinger, H. Pauli, W. Willner, and J.-C. Svenning. 2010. Going against the flow: potential mechanisms for unexpected downslope range shifts in a warming climate. Ecography 33(2):295-303. https://doi.org/10.1111/j.1600-0587.2010.06279.x
Malhi, Y., J. Franklin, N. Seddon, M. Solan, M. G. Turner, C. B. Field, and N. Knowlton. 2020. Climate change and ecosystems: threats, opportunities and solutions. Philosophical Transactions of the Royal Society B 375:20190104. https://doi.org/10.1098/rstb.2019.0104
Martin, A. E., S. L. Graham, M. Henry, E. Pervin, and L. Fahrig. 2018. Flying insect abundance declines with increasing road traffic. Insect Conservation and Diversity 11:608-613. https://doi.org/10.1111/icad.12300
Møller, A. P., D. Czeszczewik, E. Flensted-Jensen, J. Erritzøe, I. Krams, K. Laursen, W. Liang, and W. Walankiewicz. 2021. Abundance of insects and aerial insectivorous birds in relation to pesticide and fertilizer use. Avian Research 12:43. https://doi.org/10.1186/s40657-021-00278-1
Møller, A. P., D. Rubolini, and E. Lehikoinen. 2008. Populations of migratory bird species that did not show a phenological response to climate change are degrading. Proceedings of the National Academy of Sciences USA 105:16195-16200. https://doi.org/10.1073/pnas.0803825105
Muggeo, V. M. R. 2008. segmented: An R package to fit regression models with broken-line relationships. R package version 1.0-0. https://journal.r-project.org/articles/RN-2008-004/RN-2008-004.pdf
Nebel, S., A. Mills, J. D. McCracken, and P. D. Taylor. 2010. Declines of aerial insectivores in North America follow a geographic gradient. Avian Conservation and Ecology 5(2):1. https://doi.org/10.5751/ACE-00391-050201
Newton, I. 2007. Weather-related mass-mortality events in migrants. Ibis 149:453-467. https://doi.org/10.1111/j.1474-919X.2007.00704.x
Nilsson, C., R. H. G. Klaassen, and T. Alerstam. 2013. Differences in speed and duration of bird migration between spring and autumn. American Naturalist 181:837-845. https://doi.org/10.1086/670335
Nocera, J. J., J. M. Blais, D. V. Beresford, L. K. Finity, C. Grooms, L. E. Kimpe, K. Kyser, N. Michelutti, M. W. Reudink, and J. P. Smol. 2012. Historical pesticide applications coincided with an altered diet of aerially foraging insectivorous chimney swifts. Proceedings of the Royal Society B 279:3114-3120. https://doi.org/10.1098/rspb.2012.0445
Parmesan, C. 2006. Ecological and evolutionary responses to recent climate change. Annual Review of Ecology, Evolution, and Systematics 37:637-669. https://doi.org/10.1146/annurev.ecolsys.37.091305.110100
Parmesan, C. 2007. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology 13:1860-1872. https://doi.org/10.1111/j.1365-2486.2007.01404.x
Potvin, D. A., K. Välimäki, and A. Lehikoinen. 2016. Differences in shifts of wintering and breeding ranges lead to changing migration distances in European birds. Journal of Avian Biology 47(5):619-628. https://doi.org/10.1111/jav.00941
Prytula, E. D., A. E. McKellar, L. Schwitters, and M. W. Reudink. 2021. Rapid advancement of spring migration and en route adjustment of migration timing in response to weather during fall migration in Vaux’s Swifts (Chaetura vauxi). Canadian Journal of Zoology 100:56-63. https://doi.org/10.1139/cjz-2021-0089
R Core Team. 2022. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Reid, J. M., J. M. J. Travis, F. Daunt, S. J. Burthe, S. Wanless, and C. Dytham. 2018. Population and evolutionary dynamics in spatially structured seasonally varying environments. Biological Reviews 93:1578-1603. https://doi.org/10.1111/brv.12409
Rushing, C. S., J. A. Royle, D. J. Ziolkowski Jr, and K. L. Pardieck. 2020. Migratory behavior and winter geography drive differential range shifts of eastern birds in response to recent climate change. Proceedings of the National Academy of Sciences USA 117:12897-12903. https://doi.org/10.1073/pnas.2000299117
Schwartz, M. D., R. Ahas, and A. Aasa. 2006. Onset of spring starting earlier across the Northern Hemisphere. Global Change Biology 12:343-351. https://doi.org/10.1111/j.1365-2486.2005.01097.x
Schwitters, L., D. Schwitters, E. L. Bull, and C. T. Collins. 2021. Vaux's Swift (Chaetura vauxi), version 1.1. In P. G. Rodewald, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.vauswi.01.1
Sonnleitner, J., S. Lazerte, A. E. McKellar, N. J. Flood, and M. W. Reudink. 2022. Rapid shifts in migration routes and breeding latitude in North American bluebirds. Ecosphere 13:e4316. https://doi.org/10.1002/ecs2.4316
Spiller, K. J., and R. Dettmers. 2019. Evidence for multiple drivers of aerial insectivore declines in North America. Condor 121:duz010. https://doi.org/10.1093/condor/duz010
Steeves, T. K., S. B. Kearney-McGee, M. A. Rubega, C. L. Cink, and C. T. Collins. 2020. Chimney Swift (Chaetura pelagica), version 1.0. In A. F. Poole, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.chiswi.01
Strimas-Mackey, M., W. M. Hochachka, V. Ruiz-Gutierrez, O. J. Robinson, E. T. Miller, T. Auer, S. Kelling, D. Fink, and A. Johnston. 2020. Best practices for using eBird data. version 1.0. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.5281/zenodo.3620739
Strimas-Mackey, M., E. Miller, and W. Hochachka. 2018. auk: eBird data extraction and processing with R. https://cornelllabofornithology.github.io/auk/
Studds, C. E., and P. P. Marra. 2011. Rainfall-induced changes in food availability modify the spring departure programme of a migratory bird. Proceedings of the Royal Society of London B 278:3437-3443. https://doi.org/10.1098/rspb.2011.0332
Supp, S. R., F. A. La Sorte, T. A. Cormier, M. C. W. Lim, D. R. Powers, S. M. Wethington, S. Goetz, and C. H. Graham. 2015. Citizen-science data provides new insight into annual and seasonal variation in migration patterns. Ecosphere 6(1):1-19. https://doi.org/10.1890/ES14-00290.1
Thomas, C. D., and J. J. Lennon. 1999. Birds extend their ranges northwards. Nature 399:213. https://doi.org/10.1038/20335
Thorup, K., and P. B. Conn. 2009. Estimating the seasonal distribution of migrant bird species: can standard ringing data be used? Pages 1107-1117 in D. L. Thomson, E. G. Cooch, and M. J. Conroy, editors. Modeling demographic processes in marked populations. Springer, New York, New York, USA. https://doi.org/10.1007/978-0-387-78151-8_52
Vardanis, Y., R. H. G. Klaassen, R. Strandberg, and T. Alerstam. 2011. Individuality in bird migration: routes and timing. Biology Letters 7:502-505. https://doi.org/10.1098/rsbl.2010.1180
Visser, M. E., A. C. Perdeck, J. H. van Balen, and C. Both. 2009. Climate change leads to decreasing bird migration distances. Global Change Biology 15(8):1859-1865. https://doi.org/10.1111/j.1365-2486.2009.01865.x
Wood, S. N. 2011. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. Journal of the Royal Statistical Society B 73(1):3-36. https://doi.org/10.1111/j.1467-9868.2010.00749.x
Zhang, F., J. A. Biederman, M. P. Dannenberg, D. Yan, S. C. Reed, and W. K. Smith. 2021. Five decades of observed daily precipitation reveal longer and more variable drought events across much of the western United States. Geophysical Research Letters 48(7):e2020GL092293. https://doi.org/10.1029/2020GL092293
Zhang, X., G. Flato, M. Kirchmeier-Young, L. Vincent, H. Wan, X. Wang, R. Rong, J. Fyfe, G. Li, and V. V. Kharin. 2019. Changes in temperature and precipitation across Canada. Pages 112-193 in E. Bush and D. S. Lemmen, editors. Canada’s changing climate report. Government of Canada, Ottawa, Ontario, Canada. https://doi.org/10.4095/327811
Fig. 1
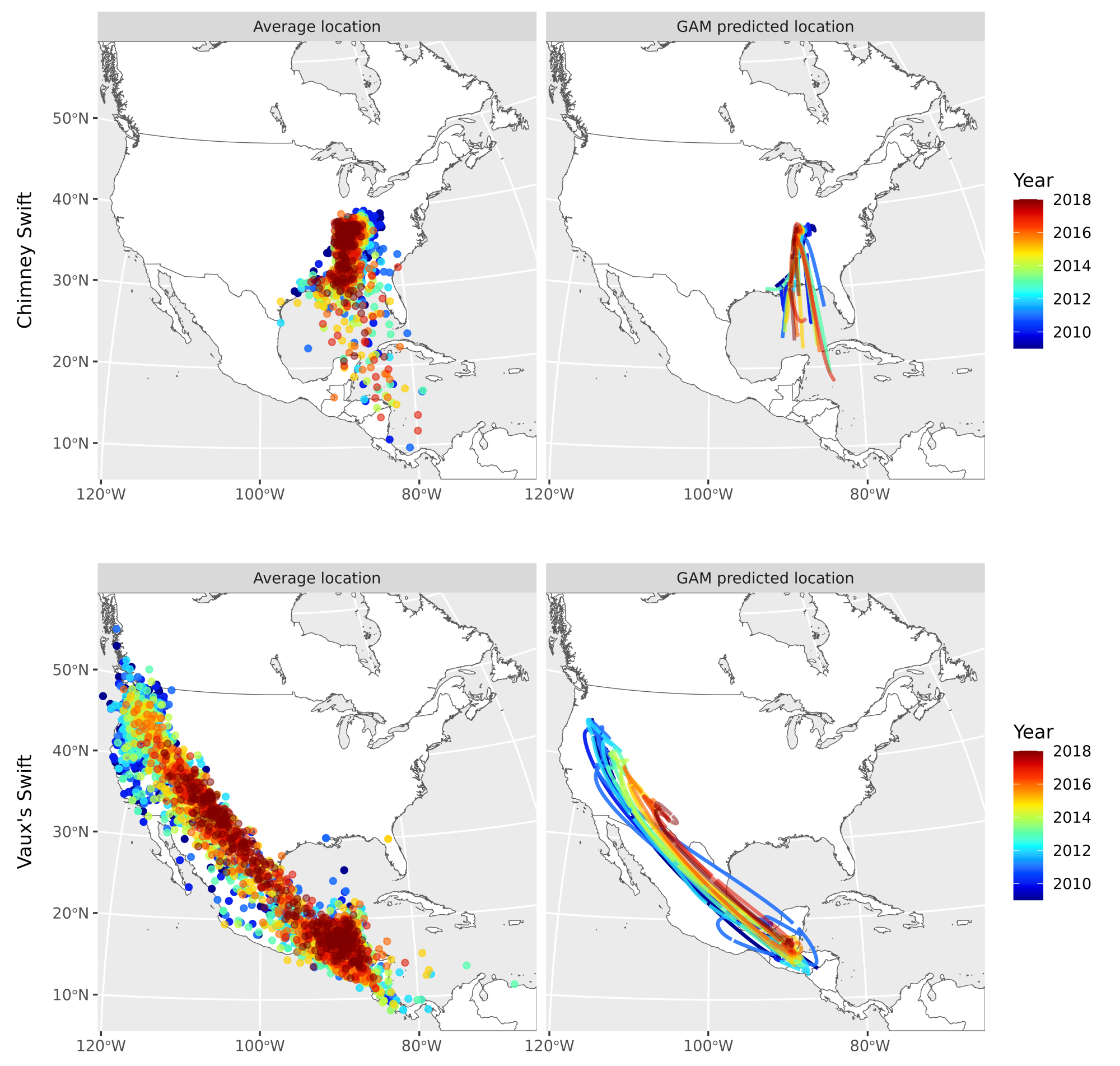
Fig. 1. Mean location of swift populations over 10 years (2009–2018) for Vaux’s Swifts (Chaetura vauxi) and Chimney Swifts (Chaetura pelagica) through spring and fall migration and on the breeding grounds. Left panel: points represent the daily mean weighted population centroid, colored by year. Right panel: lines represent the migration path predicted by population centroids, colored by year. Note that data for Chimney Swifts was too sparse in the overwintering period, so dates have been filtered to ordinal dates < 75 and > 300.
Fig. 2
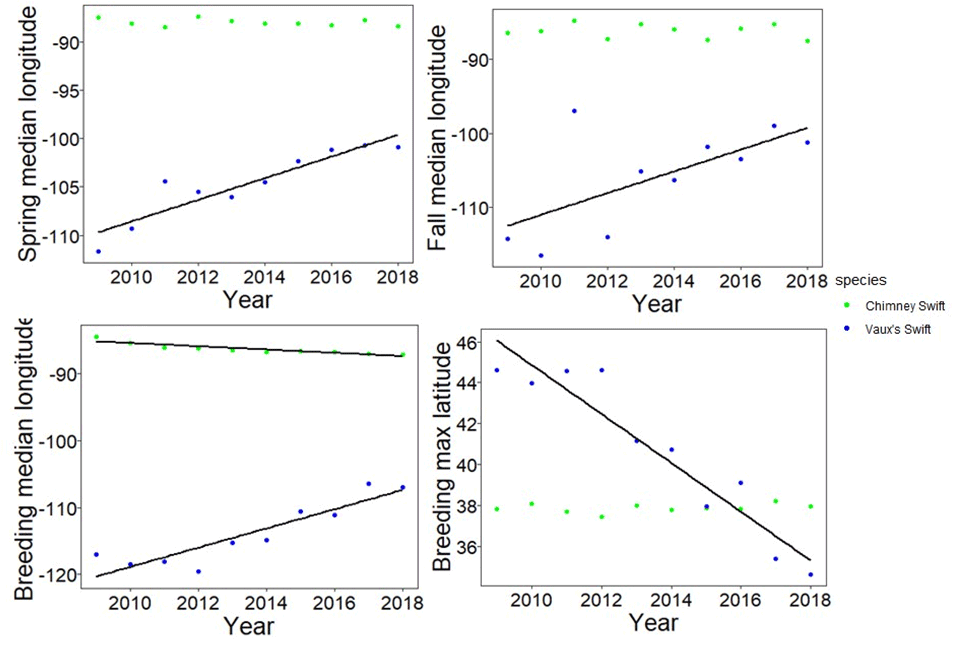
Fig. 2. Changes in the spring and fall median longitude during migration, and the breeding median longitude and maximum latitude for Vaux’s (Chaetura vauxi) and Chimney Swifts (Chaetura pelagica), calculated from daily population centroid longitudes and latitudes predicted using generalized additive models (GAMs). Best fit lines are shown for significant changes over time. Chimney Swift measures are based on the northern part of their migration only.
Table 1
Table 1. Direction and magnitude of significant changes in Vaux’s Swift (Chaetura vauxi) and Chimney Swift (Chaetura pelagica) breeding distribution, migration timing, and migration routes over a 10-year period based on eBird data. Chimney Swift measures are based on the northern part of their migration only.
| Vaux Swift | Chimney Swift | ||||||||
| Max breeding latitude | 1.2° South/year* | No change | |||||||
| Median breeding longitude | 1.4° East/year* | 0.24° West/year* | |||||||
| Spring median longitude | 1.1° East/year* | No change | |||||||
| Fall median longitude | 1.5° East/year* | No change | |||||||
| Start of spring migration | Advanced 1.4 days/year* | NA | |||||||
| End of spring migration | No change | No change | |||||||
| Start of fall migration | No change | Delayed 4.2 days/year* | |||||||
| End of fall migration | Delayed 1.6 days/year† | NA | |||||||
| Max speed spring | No change | No change | |||||||
| Max speed fall | No change | No change | |||||||
| * P < 0.05. † Non-significant trend (P = 0.06) | |||||||||