







The following is the established format for referencing this article:
Pandolfino, E. R., and L. A. Douglas. 2023. Using song dialects to reveal migratory patterns of Ruby-crowned Kinglet populations. Journal of Field Ornithology 94(3):10.ABSTRACT
Conservation of a migratory species requires knowledge not only of its breeding range, but also of its migratory path and non-breeding range. Except for timing, other aspects of the migration of the Ruby-crowned Kinglet (Corthylio calendula) remain largely unstudied, with no published data on migration routes. Breeding populations of this species in the Sierra Nevada and Cascades mountain ranges, as well as those in eastern Canada and the northeastern U.S., have experienced significant declines, whereas Rocky Mountain breeders have increased. Understanding the winter range and migratory pathways used by different breeding populations may be key to explaining these contrasting population trends. Song dialects of the Ruby-crowned Kinglet differ regionally among various breeding populations, and these dialect regions were previously mapped. Because this kinglet sings during spring migration and winter, we obtained archived, non-breeding-season recordings of song and assigned each to one of those regional song dialects. This allowed us to assess the likely winter ranges and migration pathways of different breeding populations. This approach offers some advantages over typical methods of tracking movements. Birds do not need to be captured; one can easily obtain data over large ranges and from many individuals; and it can be applied to species, such as this kinglet, that are too small to permit use of most tracking devices. We were able to assess likely winter range and spring migration routes for populations that breed in the eastern U.S. and Canada, the interior of Alaska, and for the subspecies C c. grinnelli that breeds along the Gulf of Alaska and western British Columbia. We found that kinglets breeding in the eastern portions of the range wintered in the southeastern and south-central U.S., and that their spring migrations occurred across a broad swath of the eastern U.S. Interior Alaska breeders wintered mostly in California, and the subspecies C. c. grinnelli wintered from the southernmost parts of their breeding range, south as far as northwestern California. We obtained too few winter recordings from birds using the dialects of kinglets breeding in the interior west (Rocky Mountains and the Sierra Nevada and Cascades ranges) to determine their winter range, and spring recordings were also sparse from those regions. It is likely that those interior-west breeders winter mainly in Mexico, an area with very few archived recordings. We also analyzed unpublished banding data for the Ruby-crowned Kinglet that, although providing little information about breeding-wintering range connectivity, were consistent with the migratory pathways we determined from song dialects.
RESUMEN
La conservación de una especie migratoria requiere del conocimiento no solo de su rango de reproducción, pero también de su ruta migratoria y su rango no reproductivo. Con excepción del tiempo, no han sido estudiados otros aspectos de la migración de Corthylio calendula, y no existen datos publicados en las rutas de migración. Las poblaciones reproductivas de esta especie en las cordilleras Sierra Nevada y Cascades, así como aquellas en el este de Canadá y noreste de Estados Unidos, han experimentado disminuciones significativas, mientras que las que se reproducen en las Montañas Rocosas han incrementado. Comprender el rango de invierno y las rutas migratorias utilizadas por diferentes poblaciones puede ser la clave para explicar estas tendencias poblacionales contrastantes. Los dialectos del canto en Corthylio calendula difieren regionalmente entre varias poblaciones reproductivas, y estas regiones de dialectos han sido mapeadas previamente. Esto nos permitió evaluar los rangos de invierno probables y las rutas de migración de las diferentes poblaciones. Esta aproximación provee algunas ventajas sobre otros métodos típicos de rastreo de movimiento. Las aves no deben ser capturadas; es posible obtener datos a lo largo de una gran área geográfica y de muchos individuos; y puede ser aplicado a especies, como Corthylio caléndula, que son muy pequeños para el uso de la mayoría de los aparatos de rastreo. Logramos determinar los rangos de invierno probables y la ruta de migración de primavera para las poblaciones que se reproducen en el este de Estados Unidos y Canadá, el interior de Alaska, y para la subespecie C.c. grinnelli que se reproduce a lo largo del golfo de Alaska y el oeste de British Columbia. Encontramos que Corthylio calendula que se reproduce en las regiones del este del rango geográfico, pasan el invierno en el sureste y centro sur de Estados Unidos, y que la migración de primavera ocurrió a lo largo de un amplio territorio del este de Estados Unidos. Las poblaciones que se reproducen en el interior de Alaska pasaron el invierno principalmente en California, y la subespecie C.c. grinnelli paso el invierno desde la región mas al sur de su rango de reproducción, hasta el noroeste de California. Encontramos muy pocos registros de invierno de aves utilizando los dialectos de las poblaciones que se reproducen en el oeste interior (Montañas Rocosas, y cordilleras Sierra Nevada y Cascades) para determinar su rango de invierno, y las grabaciones de primavera fueron también escasas a lo largo de estas regiones. Es probable que estas poblaciones reproductivas del oeste interior pasan el invierno principalmente en México, un área donde obtuvimos muy pocas grabaciones. También analizamos datos de anillamiento no publicados para Corthylio calendula que, a pesar de proveer poca información sobre la conectividad de los rangos reproductivos y de inverno, fueron consistentes con las rutas migratorias determinadas por los dialectos del canto.
INTRODUCTION
Migratory species present challenges to understanding their ecology and any conservation challenges they may face (Faaborg et al. 2010). Whereas most bird research is focused on the breeding season (Marra et al. 2015), migratory birds generally spend most of their year in migration or in the winter range (Newton 2008). Development of effective conservation plans for such species requires knowledge of habitat conditions in both the breeding and non-breeding ranges.
Newer tracking technologies, such as light-level geolocators or satellite tracking, have dramatically improved our ability to learn about bird migration (Stutchbury et al. 2009, Flack et al. 2022). However, these techniques have limitations. They require capturing individuals, usually twice, to attach and recover the devices, and they are limited to tagging animals above a minimum mass (Brlik et al. 2020). Bird banding may also offer glimpses into the migration path or winter range of birds, although encounter rates away from the original banding location, particularly in small passerines, are often low.
For bird species that use different regional song dialects, and also sing during migration or winter, such dialects can allow one to use recordings to assess the non-breeding ranges of birds using these distinct song dialects. This approach offers advantages over other techniques because birds do not need to be captured and large numbers of individuals can be studied from many locations. This approach was used to reveal the migratory strategies of the Puget Sound subspecies of the White-crowned Sparrow (Zonotrichia leucophrys pugetensis; DeWolfe and Baptista 1995) and the Golden-crowned Sparrow (Z. atricapilla; Pandolfino and Douglas 2021), and to determine the source of irruptive individuals of the Black-chinned Sparrow (Spizella atrogularis; Pandolfino et al. 2022). The large and rapidly growing archives of bird recordings offers promise that this approach may have broad applications to many species.
The Ruby-crowned Kinglet (Corthylio calendula) breeds from easternmost Canada and the northeastern U.S. across the boreal regions of Canada and the upper Great Lakes and into western Canada and much of Alaska (Swanson et al. 2021). The breeding range extends through most of the montane western U.S., including the Rockies, Cascades, Sierra Nevada, and higher elevations in the Great Basin region. The winter range includes the Pacific coast from southernmost British Columbia through Baja California, lower elevation inland areas of the western U.S., the U.S. Eastern Seaboard from southern New England south, most of the southern half of the eastern U.S., and south through Mexico (Swanson et al. 2021) into Guatemala (Howell and Webb 1995). According to Swanson et al. (2021), no migratory-route information is available. And the Ruby-crowned Kinglet, at six to seven grams (Swanson et al. 2021), is so small that only the largest individuals meet the criteria for tagging with the smallest currently available geolocators.
With the apparent extinction of the Guadalupe Island endemic C. c. obscurus (Quintana-Barrios et al. 2006), there are now two recognized subspecies, nominate C. c. calendula and C. c. grinnelli (Swanson et al. 2021). C. c. grinnelli breeds in coastal forests from the Gulf of Alaska through British Columbia. There is disagreement about the non-breeding movements of C. c. grinnelli. Browning (1979) asserted that this taxon is resident within its breeding range, and Campbell et al. (1997) speculated that C. c. grinnelli makes short latitudinal movements with northern breeders, possibly wintering in southern British Columbia. Others have characterized it as partially migratory with regular winter movements south along the west coast (Phillips 1991), at least into Washington (Wahl et al. 2005) and Oregon (Marshall et al. 2003), and possibly as far south as coastal central California (Dawson 1923). Winter specimens of this taxon have been collected in coastal California as far south as San Diego County (Unitt 2004).
The winter ranges of various populations of the nominate subspecies remain unknown. Some have suggested that these birds may make altitudinal movements from the western mountain breeding areas to nearby lower elevations (Browning 1979, Ryser 1985, Campbell et al. 1997, Wahl et al. 2005, Floyd et al. 2007). Hobson et al. (2014) analyzed stable isotopes from Ruby-crowned Kinglets wintering in Mexico, but the results showed a very wide range of values potentially representing most of the species’ breeding range.
Different breeding populations of the Ruby-crowned Kinglet have shown starkly different breeding-population trends since the mid-1960s (Sauer et al. 2020). Birds breeding in the easternmost parts of the range demonstrated significant negative population trends, as have breeders of the Sierra Nevada and Cascades ranges in California, Oregon, and Washington. In the Sierra Nevada of California this species was abundant and widespread in the early 20th century (Dawson 1923, Grinnell and Miller 1944), but is now an uncommon and sparsely distributed breeder (Beedy and Pandolfino 2013), even in areas where the habitat is well-protected and largely unchanged from earlier conditions (Moritz 2007). Indeed, a recent Breeding Bird Atlas in the central Sierra Nevada, where this kinglet was previously common, did not record a single observation of the species (Rose and Rose 2019). In contrast, most of the Rocky Mountain breeding populations from Canada to Arizona and New Mexico showed population gains (Sauer et al. 2020). To understand factors that may be driving these contrasting trends, we need to know the migration pathways and winter ranges of these different regional populations.
The Ruby-crowned Kinglet uses distinctly different versions of their songs (dialects) in different regions within their breeding range, and Pandolfino and Douglas (2022) recently mapped these dialects throughout this species’ breeding range. Because this kinglet also sings during spring migration and winter (Marshall et al. 2003, Wahl et al. 2005, Swanson et al. 2021), we chose to use non-breeding season recordings to assess winter ranges and migration pathways of various populations by assigning one of the regional song dialects to each individual recorded. Because spring migration tends to be generally direct in most birds species studied (Newton 2008, Nilsson et al. 2013, La Sorte et al. 2016), we hypothesized that spring migration of the Ruby-crowned Kinglet would be direct.
METHODS
We used Raven Pro software (KLYCCB 2022) to visually examine all song recordings available from xeno-canto (https://www.xeno-canto.org) and the Mark Robbins/Macaulay Library (https://www.macaulaylibrary.org) of Ruby-crowned Kinglets outside the breeding season (August through mid-May). We compared the songs of each individual recorded to the 43 dialect song types (hereafter “song types”) previously identified by Pandolfino and Douglas (2022). Following Pandolfino and Douglas (2022), the song types were based on the type of repeated phrases used in the third part (Unit C) of the song (Fig. 1). As these authors demonstrated, these phrases are visually distinct and can be unambiguously assigned to a type based on visual inspection of spectrograms (Appendix 1), and each individual appears to have only a single song type in their repertoire.
When multiple recordings at the same location on the same day were available, we assumed that all those recordings were of the same individual. The assignment of a given song type to a recording was done in a blind fashion. That is, the recordings were coded such that the authors did not know the location of the recording when assigning a song type, to avoid any possibility of unintended bias. Because many of the songs from the non-breeding period were subsong (a poorly formed vocalization given by birds still perfecting their song), presumably from young birds, we only assigned song types when the song clearly matched one of the types identified by Pandolfino and Douglas (2022). Of the 1110 recordings available from xeno-canto and the Macaulay Library (through 30 April 2022), 165 were of too poor quality to be used. Another 186 recordings were eliminated as possible duplicate recordings from the same individual (recorded on the same day and location). We eliminated 286 recordings of subsong that could not be unequivocally assigned to an adult song type. We used the same numbering system for these types as Pandolfino and Douglas (2022).
We were able to assign a song type to 473 individuals (Appendix 2). We grouped these recordings by breeding region based on song types found exclusively or predominantly within that region by Pandolfino and Douglas (2022), and we used the geographic regions identified by those authors: east, north central, northern Rockies, central Rockies, southern Rockies, west, interior Alaska, and the range of the subspecies C. c. grinnelli (Fig. 2).
We also obtained data on all encounters of banded Ruby-crowned Kinglets from the Bird Banding Laboratory archives (Celis-Murillo et al. 2021). Our sample used only data from encounters at least 500 km from the banding location (Appendix 3). We chose 500 km to eliminate or reduce examples of within-season movement as opposed to migratory movement.
We grouped the recordings and banding data into the following time frames: fall migration (August through October); winter (November through March); spring migration (April); late-spring migration (first half of May); and breeding (mid-May through July; Fig. 3). Table 1 shows proportions of recordings or banding encounters by non-breeding period. We used the time frames noted above based on eBird movement data as displayed in Swanson et al. (2021). Whereas this schedule may not apply to all populations precisely, it does represent movement timing for the bulk of the population. For example, those data show that, whereas a few individuals may begin moving north during the last week of March, spring migration appears to be largely restricted to April and early May. These dates are also consistent with earlier spring migration of males, which likely account for all song recordings we used, as demonstrated by Swanson et al. (1999).
RESULTS
General observations
We found no identifiable song types from August or September, two from October, one from November, none from December or January, and four from February. Nearly all the songs recorded in those months appeared to be subsong and could not be assigned to any song type. We found many individuals giving recognizable song types by March (98), April (315), and early May (54).
East region
Our sample included eight different song types from 241 individuals that were associated exclusively with the east breeding region (types 1, 5, 13, 21, 30, 31, 34, and 40). Song type 21 was the most common, accounting for 60% of the eastern song types in our sample. This was also the most common type found by Pandolfino and Douglas (2022) in the breeding season. The seasonal distribution of these song types (A, Fig. 4) showed the winter individuals in the southeastern U.S. (from Mississippi eastward) and along the Eastern Seaboard. During spring migration (April), most birds were found along the east coast of the U.S., with most on or near the breeding range by early May.
North-central region
We found four north-central song types (9, 26, 28, and 38) from 124 individuals. As with the east region, the most common song type in the breeding-season samples from Pandolfino and Douglas (2022) was the most common (77%) in our sample (type 9). Some winter-season individuals using those song types overlapped with the winter range of the east-region birds in the southeast and along the eastern U.S. coast (B, Fig. 4), but most were found farther west from Mississippi into Texas, with one bird recorded in Mexico. One winter individual in western Nebraska was recorded in late March (23 March) and was likely an early-spring migrant. Birds giving these song types showed spring-migration distribution across a wide swath of the eastern U.S. from Iowa to the east coast. However, birds using these song types in spring were found, on average, significantly farther west than the east-region birds (average spring migration longitude = -85.6° ± 6.6° for north-central song types, vs. -76.1° ± 4.3° for east types). As with the east-region breeders, most late-spring migrants were on or near the breeding range.
Interior Alaska
We found 40 individuals giving song types used by interior Alaska breeders (types 15, 20, 22, and 27). Of those, 33 were of song type 27, also the most common type found by Pandolfino and Douglas (2022). Most of the winter recordings were from California, with a few from Arizona and Washington, and all spring migrants were close to the west coasts of California, Oregon, Washington, and British Columbia (A, Fig. 5).
Range of C. c. grinnelli
Kinglets in the breeding range of this subspecies used two song types (18 and 38) almost exclusively (Pandolfino and Douglas 2022). We found 10 winter-season birds giving these song types from the southern tip of Vancouver Island south through Washington, Oregon, and into California (B, Fig. 5). Spring migrants were recorded along the Pacific coast from Washington, through British Columbia, and into Alaska.
Interior west
This area includes four of the regions identified by Pandolfino and Douglas (2022) encompassing the Rocky Mountains from northern Canada to Arizona, and the Cascades and Sierra Nevada ranges from Washington to California. We found too few recordings of interior-west song types (42 recordings in our sample for all four regions) to derive useful information about winter ranges or migration. For example, our winter recordings of song types from the northern, central, and southern Rockies, and the west regions, numbered just 2, 3, 6, and 1, respectively. In addition, some of the song types from those regions were widespread through more than one breeding region (Pandolfino and Douglas 2022). The few spring-migration examples suggest a fairly direct northward migration, but characterization of migratory pathways is not possible without more information about the winter range of these populations.
Banding data
Banding data provided information about the movements of 46 different individuals (Fig. 6). Unfortunately, we found only a single bird for which there was information about both the winter and breeding locations. That individual was banded 16 July in western Quebec and was encountered the following winter (22 January) in the panhandle of Florida. In addition, three other individuals banded in the first week of May, most likely on or near the breeding range, were encountered in winter. Two of those birds were banded on 6 May and 9 May in southern Ontario and encountered in winter in North Carolina and eastern Texas, respectively, within the winter range inferred from our song-dialect work. The other was banded on 6 May in the Sierra Nevada of California and encountered 20 January in northern Mexico. Kinglets wintering in the southeastern U.S. all remained in the two regions expected from the dialect data, and all the birds banded in the west were encountered in the west. Even the individual showing the longest migration (banded in early spring on the Louisiana coast) bred in central Alberta, within the expected north-central region (longest line in Fig. 6). Thus, all these encounters with banded birds were consistent with the migration paths suggested by our song dialect analyses.
Vagrants
Among the 1066 recordings used in this study and by Pandolfino and Douglas (2022), we assumed that eight were vagrants (Table 2): that is, birds that had wandered well beyond the expected breeding or migration areas based on the song types used. Two of these were birds recorded in the west, singing song types from eastern breeding regions, and six were recorded in the east, singing western-type song dialects.
DISCUSSION
Broader application of the method
To our knowledge, this approach to studying bird movements has only been applied in three prior studies (DeWolfe and Baptista 1995, Pandolfino and Douglas 2021, Pandolfino et al. 2022). The large and growing archives of bird song should allow this method to be applied to many other species. This offers the advantages of birds not needing to be captured, and many individuals across a broad range being able to be sampled very economically. Application of this method requires that the species show distinct regional variation in song and that individuals sing during the non-breeding season. It is also very helpful if the species has only one, or very few, song types in its repertoire. Because nearly all such species studied appear to maintain this repertoire throughout their lives (Marler and Slabbekoorn 2004, Catchpole and Slater 2008), one can be reasonably confident that a song type encountered outside the breeding range will match the one used in that range. One must also be aware that females of many species sing. However, in most cases, as with the Ruby-crowned Kinglet (Pieplow 2017), their song is easily distinguished from that of the male. For example, female song of the Ruby-crowned Kinglet lacks Unit C entirely (Pieplow 2017). However, Pieplow (2017) notes singing mainly from February through August, and our analyses of the available recordings (Pandolfino and Douglas, personal observation) showed only a few examples of poorly developed song during fall, with progressively larger proportions of fully crystallized adult song from March through May. By late May through July, all birds recorded were singing fully developed songs.
Migration routes and winter ranges
Our approach does not reveal migration routes of individuals but provides an overview of winter ranges and migration routes for subpopulations. For example, individual birds may make significant intra-winter movements within the winter range and we did not assume that the location of any individual bird in our sample was stable throughout the winter period. Ruby-crowned Kinglets using the song dialects of the two easternmost regions (east and north central) appeared to take a direct spring-migration route from their winter range to the breeding range. Although we had fewer data points for breeders from interior Alaska and members of the C. c. grinnelli subspecies, those spring migrations also appeared to be direct with spring migrants found mainly along the west coast.
Those results are consistent with the general pattern of most migratory birds for which spring migration is faster and more direct than fall migration (Newton 2008, Nilsson et al. 2013, La Sorte et al. 2016). This difference is believed to be driven by the importance of arriving on the breeding grounds early enough to find and occupy the best territories (Myers 1981, Gauthreaux 1982, Francis and Cooke 1986, Chandler and Mulvihill 1990, Newton 2008). Female Ruby-crowned Kinglets may give a short version of song lacking the last part (Unit C; Pieplow 2017), so we assumed that all the singing individuals in our sample were males. Male Ruby-crowned Kinglets migrate earlier than females (a common observation among most passerines studied; Newton 2008), allowing them to establish breeding territories before females arrive (Swanson et al. 1999). Males also appear to winter farther north than females (Fairfield and Shirokoff 1978, Swanson et al. 1999); thus, the winter ranges inferred from our song data may be skewed northward.
Based on the winter locations of birds singing the dialects of C. c. grinnelli, some members of this taxon migrate well south of the breeding range, which is contra Browning (1979) and Campbell et al. (1997). We found these individuals at the southern tip of Vancouver Island, British Columbia, and south into Washington, Oregon, and northwestern California. Of course, females wintering farther south would not have been found by our method, and the winter range of this subspecies may well extend through the Marine West Coast Forest ecoregion in California.
The migration routes and wintering ranges of birds breeding in the western mountain ranges (Rockies, Sierra Nevada–Cascades) remain largely unknown due to the small number of recordings we found from birds using those dialects and the widespread nature of some of those dialects. It seems likely that many or most of those western montane breeders may winter in Mexico. However, we have information on only two individuals wintering in Mexico: one in Mexico City (south-central Mexico) giving a song type exclusively associated with breeders in the north-central region, and another banded in the central Sierra Nevada in May and encountered in January in northwestern Mexico.
Our data included only one migrating bird west of the Mississippi River using an east-region song; therefore, it seems unlikely that significant numbers of east-region breeders winter in Mexico. Stable isotope analysis of samples from Ruby-crowned Kinglets wintering in Mexico (Hobson et al. 2014) were consistent with this assumption, suggesting most birds may have bred in the northern Rockies from Canada into Wyoming. However, the full range of values reported in that study was broad enough to encompass nearly the entire breeding range.
Migratory connectivity, the extent to which birds breeding in one area also share a wintering area, is not possible to assess from our data. Whereas there may be groups of individual Ruby-crowned Kinglets that have a high degree of migratory connectivity, the relatively widespread nature of the song dialects we observed suggests only a very diffuse sort of connectivity. For example, song type 21, for which we have the most examples, is found in the breeding range from Newfoundland, south into upstate New York, and in winter along the U.S. Eastern Seaboard from Connecticut to Florida and west into eastern Louisiana.
Our samples included only three song types for which we had robust numbers (> 10) of observations from both the breeding range and the wintering range. Therefore, our data were insufficient to assess whether the Ruby-crowned Kinglet uses a chain-migration strategy (in which the most northern breeders also winter the farthest north), or leapfrog migrations (in which the most northern breeders winter the farthest south). And the lack of data from Mexico, where many Ruby-crowned Kinglets winter, makes any estimate of typical wintering latitudes suspect.
Vagrancy
Vagrancy, when a bird is observed well away from its expected breeding, wintering, or migration range, is observed among nearly all migratory species (Newton 2008). Such birds are most often found in spring or fall and are usually young birds making their first migration from the breeding area or from their wintering range (De Sante 1983a, De Sante 1983b, Newton 2008). Many of these birds are believed to have an anomalous sense of orientation in which they choose to migrate on a trajectory different from that of most members of that species. This misorientation can vary among individuals from a few degrees to 180 degrees, resulting in migration in the opposite direction from normal (De Sante 1983b, Newton 2008).
Ruby-crowned Kinglets have been observed as vagrants as far east as Scotland (BOURC 2022) and as far west as Attu Island at the western end of the Aleutian Chain (Dunn and Alderfer 2011). Our use of mapped song dialects allowed us to detect so-called cryptic vagrant Ruby-crowned Kinglets that would never have been noted as being out of range. Seven of the vagrants were recorded within regions and on dates when the species is common. Thus, only the detection of unexpected song dialects revealed that they were outside the expected range. The exception was a bird recorded 18 May in Baja California, Mexico, a date when Ruby-crowned Kinglets are not expected in that area. In each case, these birds were using a dialect otherwise unrecorded in that area. The fact that more vagrants were detected in the east than the west may reflect the higher concentration of birders in the east, though our sample size is too small to draw any conclusions.
Priorities for future research
The paucity of recordings from Mexico leaves an important conservation question unanswered. The starkly contradictory trends among birds breeding in the Rocky Mountains versus those breeding in the Sierra Nevada–Cascades range may be related to the conditions in the wintering ranges of those birds. A concerted effort to record songs of the Ruby-crowned Kinglet in Mexico in March could yield important information, especially about any difference in wintering ranges among populations. Additional recordings may help reveal details about migratory strategy (e.g., chain versus leapfrog) or connectivity, as well.
Other methods could complement our results and provide more information about movements of individual birds. Stable isotope analyses of feathers can produce information about the molting locations of birds (Hobson 1999), and Ruby-crowned Kinglets are believed to molt on the breeding grounds (Swanson et al. 2021). However, the wide range of isotope values from Ruby-crowned Kinglets reported by Hobson et al. (2014) revealed only very general information about breeding ranges. Tagging Ruby-crowned Kinglets with MOTUS (https://www.motus.org) transmitters, which are light enough to use with kinglets, could reveal much more about movements of individuals, at least in areas with a high density of receivers.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
This sort of research would not be possible without the archives of sound recordings maintained by xeno-canto (https://www.xeno-canto.org), the Mark Robbins Macaulay Library (https://www.macaulaylibrary.org) and the many recordists who contribute their recordings to those archives. Appendix 2 includes a complete list of those recordists. Comments and suggestions from two reviewers allowed us to improve the focus and clarity of the manuscript. We are also grateful for the banding data made available by the U.S. Geological Survey’s Bird Banding Laboratory (https://www.usgs.gov/labs/bird-banding-laboratory) and the efforts of bird banders throughout North America. This material uses data from the eBird Status and Trends Project (https://ebird.org/science/status-and-trends) at the Cornell Lab of Ornithology, eBird.org. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the Cornell Lab of Ornithology. The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
DATA AVAILABILITY
All data will be made fully available. In fact, all the raw data are in the Appendices and all recordings are available to the public.
LITERATURE CITED
Beedy, E. C., and E. R. Pandolfino. 2013. Birds of the Sierra Nevada: their natural history, status, and distribution. University of California Press, Berkeley, California, USA. https://doi.org/10.1525/9780520954472
British Ornithologist’s Union Records Committee (BOURC). 2022. British Ornithologists’ Union Records Committee (BOURC): 53rd report (January 2022). Ibis 164:363-368. https://doi.org/10.1111/ibi.13014
Brlik, V. 2020. Weak effects of geolocators on small birds: a meta-analysis controled for phylogeny and publication bias. Journal of Animal Ecology 89:207-220. https://doi.org/10.1111/1365-2656.12962
Browning, M. R. 1979. A review of geographic variation in continental populations of the Ruby-crowned Kinglet (Regulus calendula). Nemouria 21:1-9.
Campbell, R. W., N. K. Dawe, I. McTaggart-Cowan, J. M. Cooper, G. W. Kaiser, M. C. E. McNall, and G. E. Smith. J. 1997. The birds of British Columbia, volume 3: passerines - flycatchers through vireos. University of British Columbia Press, Vancouver, British Columbia, Canada.
Catchpole, C. K., and P. J. B. Slater. 2008. Bird song: biological themes and variations. Cambridge University Press, Cambridge, UK. https://doi.org/10.1017/CBO9780511754791
Celis-Murillo A., M. Malorodova, and E. Nakash. 2021. North American bird banding dataset 1960–2021 retrieved 2021-07-25. U.S. Geological Survey, Eastern Ecological Science Center at the Patuxent Research Refuge, Laurel, Maryland, USA.
Chandler, C. R., and R. S. Mulvihill. 1990. Interpreting differential timing of capture of sex classes during spring migration. Journal of Field Ornithology 61:85-89.
Dawson, W. L. 1923. The birds of California. Volume two. South Moulton Company, San Diego, California, USA.
De Sante, D. F. 1983a. Annual variability in the abundance of migrant landbirds on southeast Farallon Island, California. Auk 100:826-852. https://doi.org/10.1093/auk/100.4.826
De Sante, D. F. 1983b. Vagrants: when orientation or navigation goes wrong. Point Reyes Bird Observatory Newsletter 61:12-16.
DeWolfe, B. B., and L. F. Baptista. 1995. Singing behavior, song types on their wintering grounds and the question of leapfrog migration in Puget Sound White-crowned Sparrows. Condor 97:376-389. https://doi.org/10.2307/1369024
Dunn, J. L., and J. Alderfer. 2011. Field guide to the birds of North America. Sixth edition. National Geographic Society, Washington, D.C., USA.
Faaborg, J., R. T. Holmes, A. D. Anders, K. L. Bildstein, K. M. Dugger, S. A. Gauthreaux, Jr., P. Heglund, K. A. Hobson, A. E. Jahn, D. H. Johnson, et al. 2010. Recent advances in understanding migration systems of New World land birds. Ecological Monographs 80:3-48. https://doi.org/10.1890/09-0395.1
Fairfield, D. M., and P. A. Shirokoff. 1978. Migratory patterns and winter distribution in the Ruby-crowned Kinglet (Regulus calendula). Blue Bill (suppl.) 25:22-25.
Fink, D., T. Auer, A. Johnston, M. Strimas-Mackey, S. Ligocki, O. Robinson, W. Hochachka, L. Jaromczyk, A. Rodewald, C. Wood, et al. 2022. eBird Status and Trends, Data Version 2021. Cornell Lab of Ornithology, Ithaca, New York. https://doi.org/10.2173/ebirdst.2021
Flack, A., E. O. Aikens, A. K. Kölzsch, E. Nourani, K. R. S. Snell, W. Fiedler, N. Linek, H-G. Bauer, K. Thorup, J. Partecke, et al. 2022. New frontiers in bird migration research. Current Biology 32:R1187-R1199. https://doi.org/10.1016/j.cub.2022.08.028
Floyd, T., Elphick, C. S., Chisholm, G., Mack, K., Elston, R. G., Ammon, E. M., and Boone, J. D. 2007. Atlas of the breeding birds of Nevada. University of Nevada, Reno, Nevada, USA.
Francis, C. M., and E. Cooke. 1986. Differential timing of spring migration in wood warblers (Parulinae). Auk 103:548-556.
Gauthreaux, S. A., Jr. 1982. The ecology and evolution of avian migration systems. Avian Biology 6:93-168. https://doi.org/10.1016/B978-0-12-249406-2.50011-3
Grinnell, J., and A. H. Miller. 1944. The distribution of the birds of California. Pacific Coast Avifauna No. 27. Cooper Ornithological Society, Berkeley, California, USA.
Hobson, K. A. 1999. Tracing origins and migration of wildlife using stable isotopes: a review. Oecologia 120:314-326. https://doi.org/10.1007/s004420050865
Hobson, K. A., S. L. Van Wilgenburg, J. Faaborg, J. D. Toms, C. Rengifo, A. L. Sosa, Y. Aubry, and R. B. Aguilar. 2014. Connecting breeding and wintering grounds of Neotropical migrant songbirds using stable hydrogen isotopes: a call for an isotopic atlas of migratory connectivity. Journal of Field Ornithology 85:237-257. https://doi.org/10.1111/jofo.12065
Howell, S. N. G., and S. Webb. 1995. A guide to the birds of Mexico and Northern Central America. Oxford University Press, New York, New York, USA.
Humple, D. L., N. Nadav, G. R. Geupel, and M. P. Lynes. 2001. Female-biased sex ratio in a wintering population of Ruby-crowned Kinglets. Wilson Bulletin 113:419-424. https://doi.org/10.1676/0043-5643(2001)113[0419:FBSRIA]2.0.CO;2
K. Lisa Yang Center for Conservation Bioacoustics (KLYCCB). 2022. Raven Pro: Interactive Sound Analysis Software, Version 1.6.3. Cornell Lab of Ornithology, Ithaca, New York, New York, USA. https://ravensoundsoftware.com/
La Sorte, F. A., D. Fink, W. M. Hochachka, and S. Kelling. 2016. Convergence of broad-scale migration strategies in terrestrial birds. Proceedings of the Royal Society B 283:20152588. https://doi.org/10.1098/rspb.2015.2588
Marler, P., and H. Slabbekoorn. 2004. Nature’s music: the science of birdsong. Elsevier Academic Press, San Diego, California, USA.
Marra, P. P., E. B. Cohen, S. R. Loss, J. E. Rutter, and C. M. Tonra. 2015. A call for full annual cycle research in animal ecology. Biology Letters 11:20150552. https://doi.org/10.1098/rsbl.2015.0552
Marshall, D. B., M. G. Hunter, and A. L. Contreras. 2003. Birds of Oregon: a general reference. Oregon State University Press, Corvallis, Oregon, USA.
Moritz, C. 2007. Final report: a re-survey of the historic Grinnell-Storer vertebrate transect through Yosemite National Park, California. Museum of Vertebrate Zoology, Berkeley, California, USA. http://mvz.berkeley.edu/Grinnell/pdf/2007_Yosemite_report.pdf
Myers, J. F. 1981. A test of three hypotheses for latitudinal segregation of the sexes in wintering birds. Canadian Journal of Zoology 59:1527-1534. https://doi.org/10.1139/z81-207
Newton, I. 2008. The migration ecology of birds. Academic Press, London, UK.
Nilsson, C., R. H. G. Klaassen, and T. Alerstam. 2013. Differences in speed and duration of bird migration between spring and autumn. American Naturalist 181:837-845. https://doi.org/10.1086/670335
Pandolfino, E. R., and L. A. Douglas. 2021. Using song dialects to assess the migration strategy of the Golden-crowned Sparrow. Western Birds 52:311-321. https://doi.org/10.21199/WB52.4.3
Pandolfino, E. R., and L. A. Douglas. 2022. Regional song dialects of the Ruby-crowned Kinglet. Journal of Field Ornithology 93:6. https://doi.org/10.5751/JFO-00120-930206
Pandolfino, E. R., L. A. Douglas, and C. Ray. 2022. Irruptive movements of the Black-chinned Sparrow (Spizella atrogularis) in response to variations in precipitation: implications for climate change resiliency. Western Wildlife 9:24-37.
Phillips, A. R. 1991. The known birds of North and Middle America, part II: Bombycillidae; Sylviidae to Sturnidae; Vireonidae. A. R. Phillips, Denver, Colorado, USA.
Pieplow, N. 2017. Field guide to bird sounds of eastern North America. Houghton Mifflin Harcourt, New York, New York, USA.
Quintana-Barrios, L., G. Ruiz-Campos, P. Unitt, and R. A. Erickson. 2006. Update on the birds of Isla Guadalupe, Baja California. Western Birds 37:23-36.
Rose, S., and Rose, D. 2019. Breeding bird atlas of Nevada County, California. Sierra Foothills Audubon Society, Grass Valley, California, USA.
Ryser, F. A. 1985. Birds of the Great Basin. University of Nevada Press, Reno, Nevada, USA.
Sauer, J. R., W. A. Link, and J. E. Hines. 2020. The North American breeding bird survey, results and analysis 1966–2019. U.S. Geological Survey Eastern Ecological Science Center at the Patuxent Research Refuge, Laurel, Maryland, USA.
Stutchbury, B. J. M., S. A. Tarof, T. Done, E. Gow, P. M. Kramer, J. Tautin, J. W. Fox, and V. Afanasyev. 2009. Tracking long-distance songbird migration by using geolocators. Science 323:896. https://doi.org/10.1126/science.1166664
Swanson, D. L., E. T. Liknes, and K. L. Dean. 1999. Differences in migratory timing and energetic condition among sex/age classes in migrant Ruby-crowned Kinglets. Wilson Bulletin 111:61-69.
Swanson, D. L., J. L. Ingold, and G. E. Wallace. 2021. Ruby-crowned Kinglet (Corthylio calendula), version 1.1. In A. F. Poole and F. B. Gill, editors. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.ruckin.01.1
Unitt, P. 2004. San Diego County bird atlas. Proceedings of the San Diego Society of Natural History 39:1-645.
Wahl, T. R., B. Tweit, and S. G. Mlodinow. 2005. Birds of Washington: status and distribution. Oregon State University Press, Corvallis, Oregon, USA.
Fig. 1
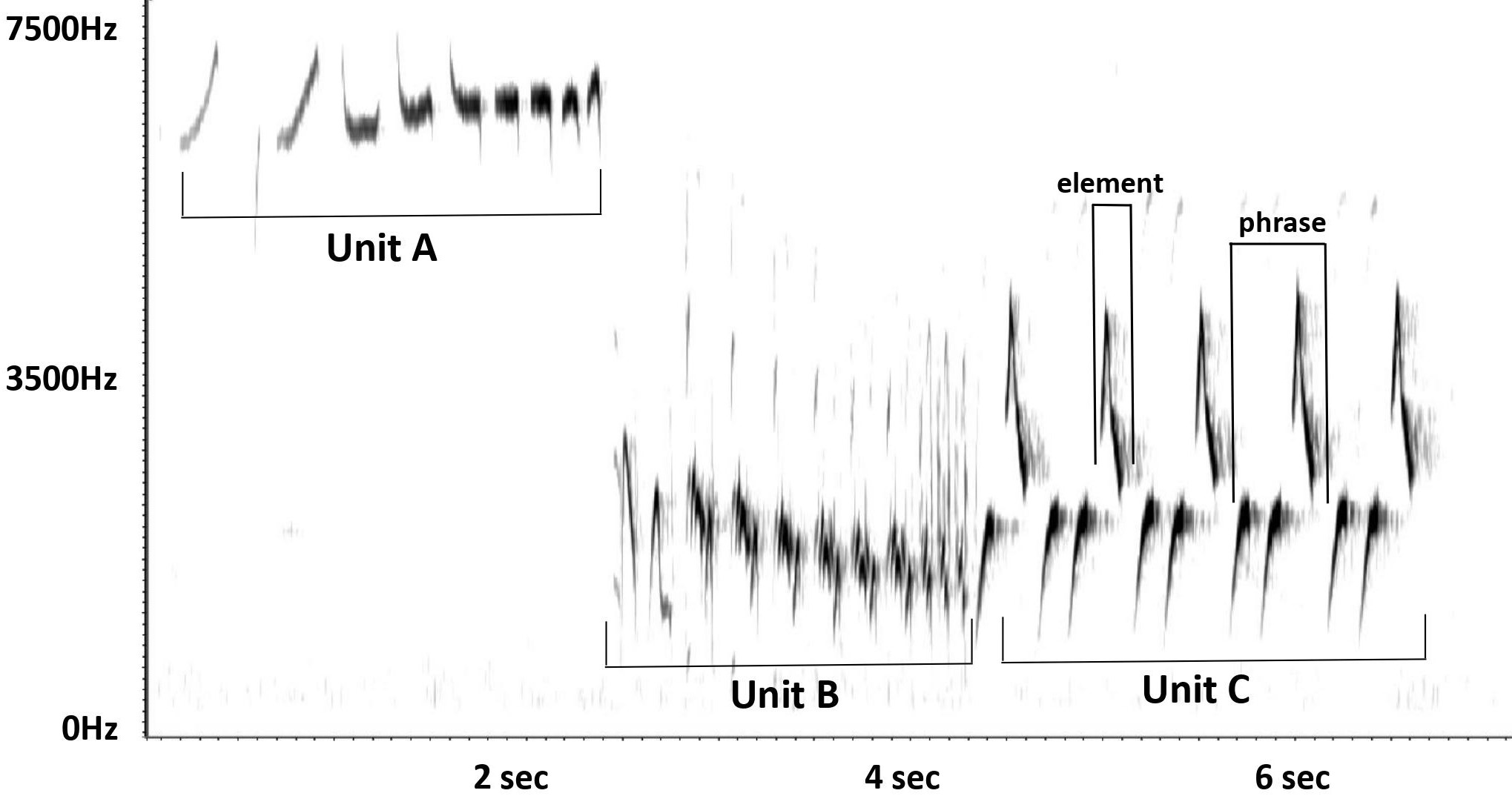
Fig. 1. Spectrograph of a typical Ruby-crowned Kinglet (Corthylio calendula) song.
Fig. 2
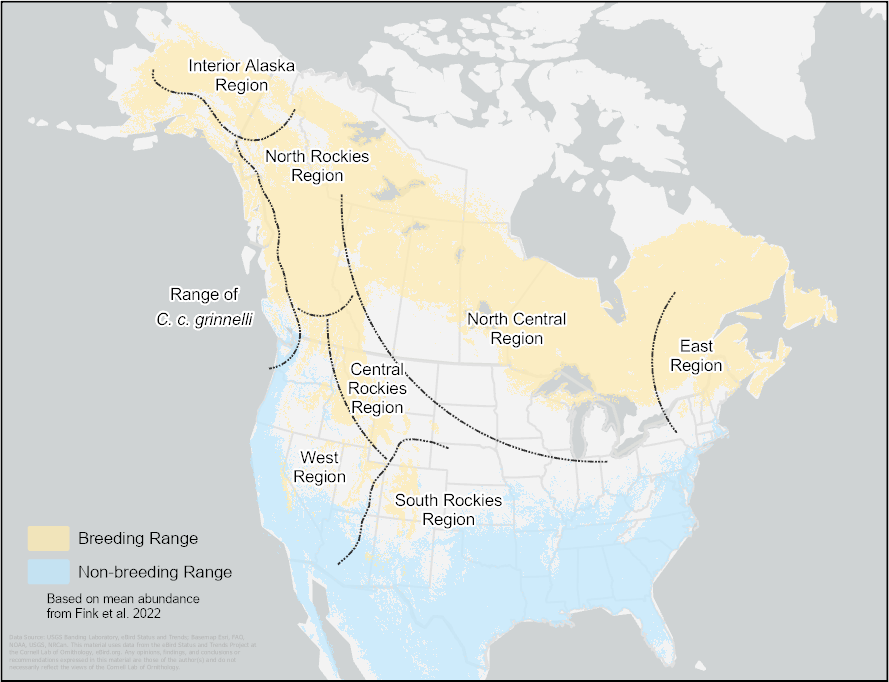
Fig. 2. The song dialect breeding regions used by Pandolfino and Douglas (2022). Ranges (breeding shaded yellow, winter shaded blue) based on Fink et al. (2022).
Fig. 3

Fig. 3. The time periods used to assign the migratory seasons for the Ruby-crowned Kinglet (Corthylio calendula).
Fig. 4
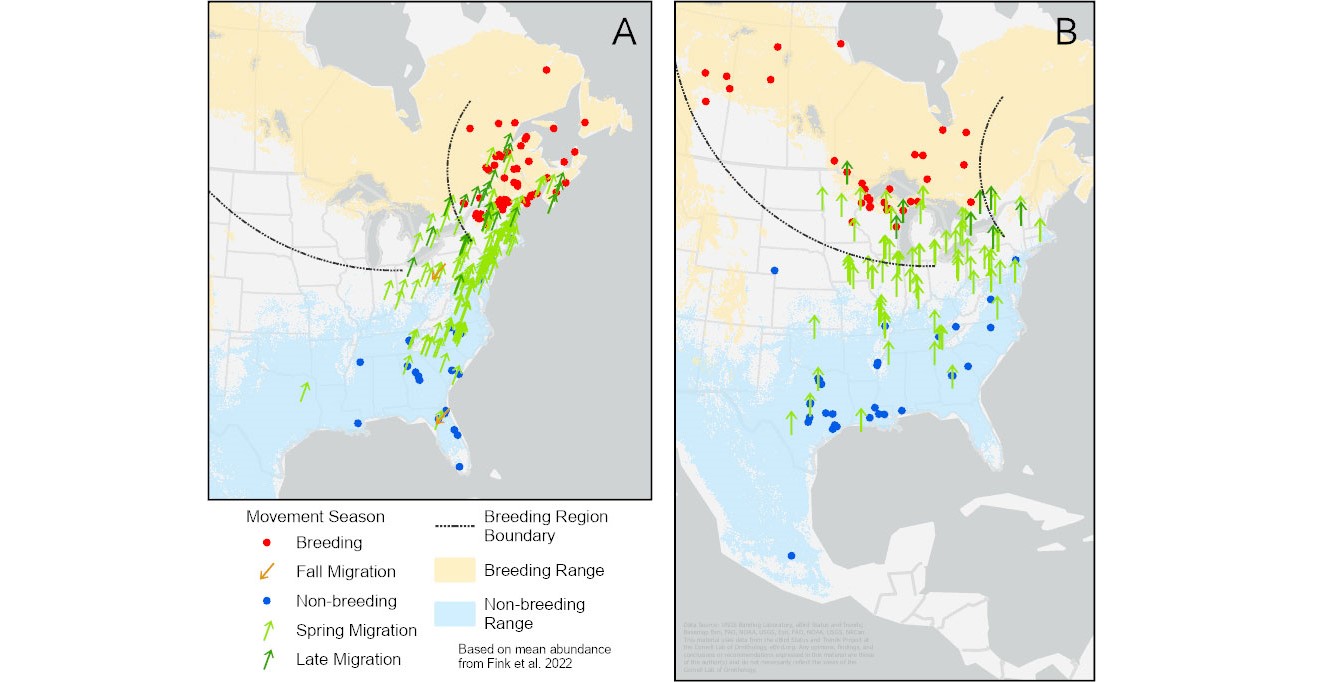
Fig. 4. The locations of individuals using song dialects of the east region (A) and the north-central region (B). Blue dots represent recordings made from November through March; light green arrows, recordings from April; dark green arrows, recordings from the first half of May; brown arrows, recordings from August through October; red dots, recordings from mid-May through July (Pandolfino and Douglas 2022, but only including song types found in this study). Ranges (breeding shaded yellow, winter shaded blue) based on Fink et al. (2022).
Fig. 5
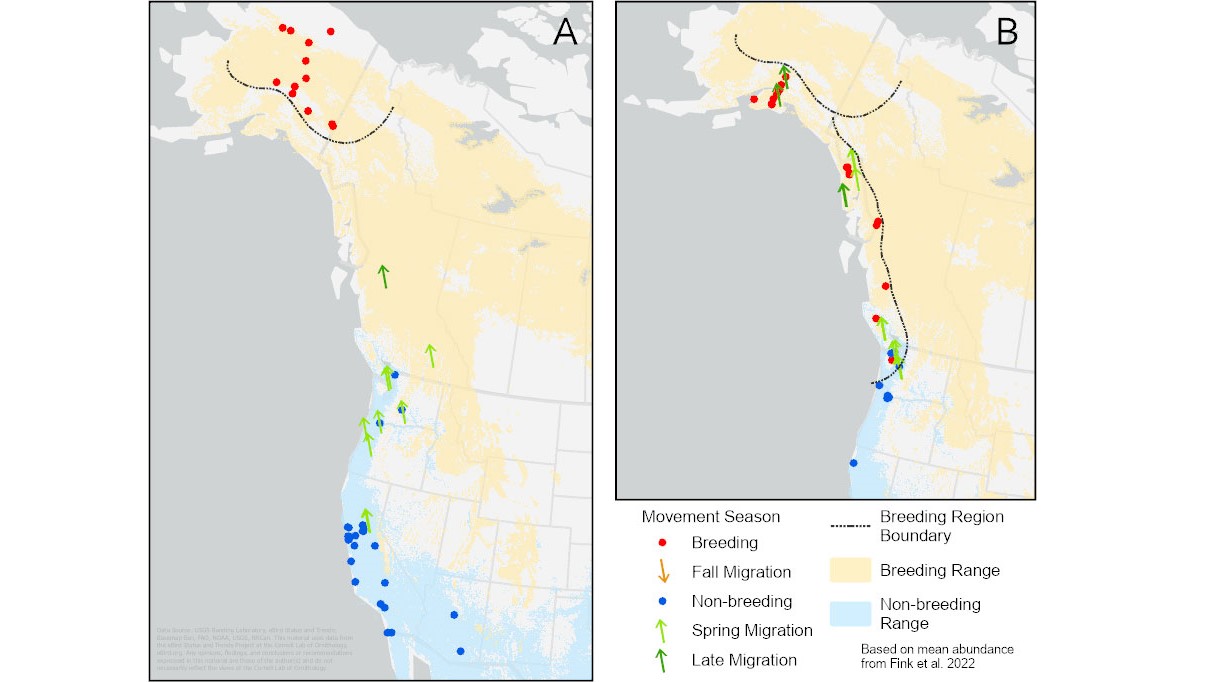
Fig. 5. The locations of individuals using song dialects of the interior Alaska region (A) and those the subspecies Corthylio calendula grinnelli (B). Blue dots represent recordings made from November through March; light green arrows, recordings from April; dark green arrows, recordings from the first half of May; brown arrows, recordings from August through October; red dots, recordings from mid-May through July (from Pandolfino and Douglas 2022, but only including song types found in this study). Ranges (Breeding shaded yellow, Winter shaded blue) based on Fink et al. (2022).
Fig. 6
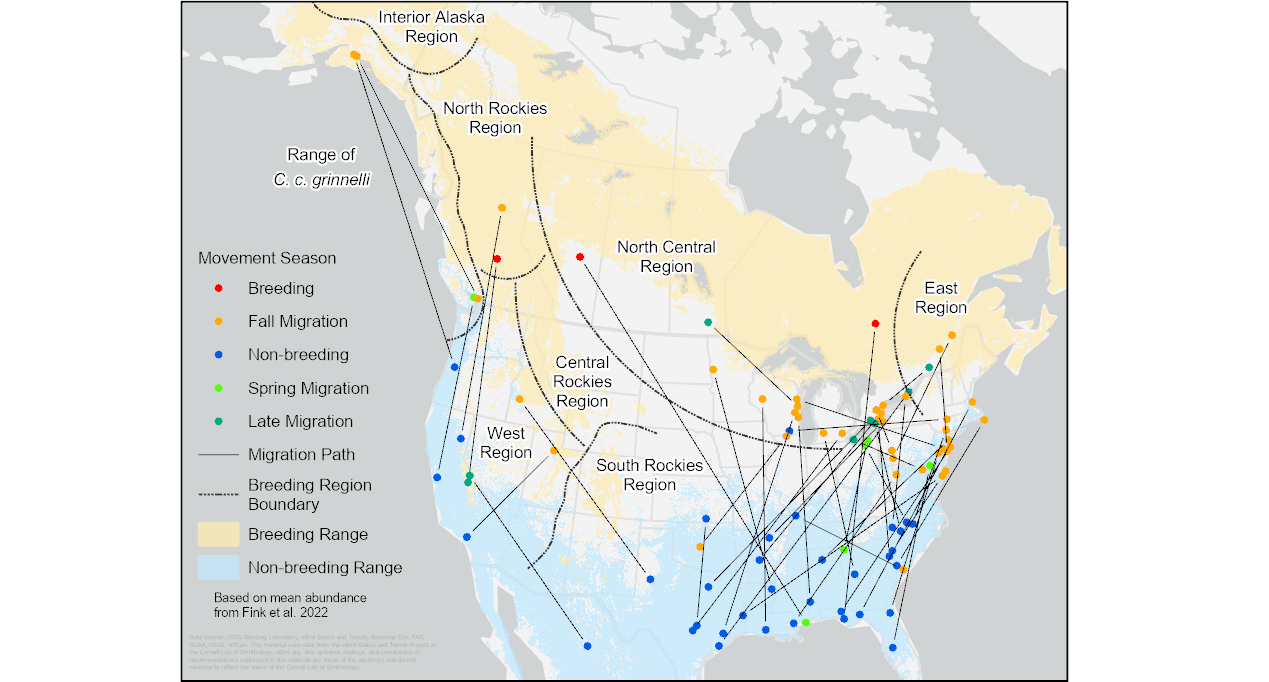
Fig. 6. Banding and encounter locations of banded Ruby-crowned Kinglets (Corthylio calendula). Colors represent the same time periods used in the previous figures. Lines link the locations for each individual. Ranges (breeding shaded yellow, winter shaded blue) based on Fink et al. (2022).
Table 1
Table 1. Number and proportion by season of recorded individuals and banding/encounter events.
| Data source | Fall | Winter | Spring migration | Late spring migration | Breeding |
| Recordings | 2 (<1%)† | 102 (13%) | 315 (40%) | 54 (7%) | 313 (40%)‡ |
| Banding | 41 (44%) | 35 (37%) | 6 (6%) | 8 (9%) | 4 (4%) |
| †As noted in text, nearly all fall recordings were of subsong. ‡Recordings from Pandolfino and Douglas 2022. | |||||
Table 2
Table 2. Apparent vagrant Ruby-crowned Kinglets (Corthylio calendula) based on song types encountered well beyond the expected breeding/migration range. Catalog numbers preceded by “xc” are from xeno-canto, others from the Macaulay Library.
| Cat No. | Recordist | Date | Type | Expected breeding range | Recording location | ||||
| 112050 | Charles Duncan | 31-May | 19 | Central Rockies | Maine | ||||
| 151835141 | Kriss Replogle | 18-Apr | 22 | Interior Alaska | New Jersey | ||||
| 225100031 | Daniel McDermott | 19-Apr | 25 | †North Rockies + | Massachusetts | ||||
| 227321191 | Jeff Ellerbusch | 25-Apr | 25 | †North Rockies + | New Jersey | ||||
| 428783001 | Natasza Fontaine | 26-Mar | 25 | †North Rockies + | South Carolina | ||||
| 160909771 | Justyn Stahl | 18-May | 31 | East | Baja Mexico | ||||
| 333163191 | Teresa Dolman | 1-May | 31 | East | Southern Alberta | ||||
| xc1217 | Don Jones | 11-Apr | 33 | South Rockies | New Jersey | ||||
| †Song Type 25 was widely distributed in the western breeding range with most breeding season examples from the North Rockies Region, but others found from southern Alaska to California. | |||||||||