







The following is the established format for referencing this article:
Brewer, D.E., T.M. Gehring, B.T. Shirkey, and J.W. Simpson. 2023. King Rail (Rallus elegans) response to audio playback: implications for population estimation, monitoring methodology, and trapping approach. Journal of Field Ornithology 94(2):7.ABSTRACT
Conspecific audio can be broadcast to improve detection probability (detectability) of secretive marsh bird species for population monitoring purposes and as a lure to more effectively trap individuals. Our primary objective was to describe King Rail (Rallus elegans) detectability as a function of distance and so determine if the distance sampling assumption of perfect detectability immediately adjacent to survey points was violated during call-broadcast surveys. We also described what factors affected King Rail detectability during audio broadcast surveys, and how and when this species was detected, and compared effectiveness of various King Rail call recordings used during trapping attempts. To accomplish these objectives, we experimentalized trapping efforts, radio-tagged 12 King Rails, and conducted repeated call-broadcast surveys for each radio-tagged individual in Michigan and Ohio, USA during the 2020 and 2021 breeding seasons. We found that King Rail detectability during 81 surveys was 0.39, which violates the aforementioned distance sampling assumption and necessitates a correction factor. Detectability within survey segments varied among survey periods, though overall detectability remained relatively constant, even after call-broadcast surveys in our study area typically end (post-survey period: 15 June to 10 July), which indicates that King Rail monitoring could effectively continue later than currently occurs. We found that grunt calls resulted in more captures than kek calls and that a multi-call track was most effective at trapping King Rails. Our findings could improve King Rail population estimates and increase success during trapping efforts, and may inform similar efforts for other secretive marsh birds.
RESUMEN
La reproducción de grabaciones con-especificas pueden mejorar la probabilidad de detección (detectabilidad) de aves sigilosas de pantano con propósitos de monitoreo poblacional y como carnada para capturar individuos más eficientemente. Nuestro objetivo principal fue describir la detectabilidad de Rallus elegans como una función de la distancia para determinar si el supuesto de muestreo por distancia de detección perfecta de individuos inmediatamente adyacentes a los puntos de muestreo se violaba durante los monitoreos que utilizan reproducción de llamados. También describimos los factores que afectan la detectabilidad de Rallus elegans durante los monitoreos utilizando reproducción de audio, y como y cuando esta especie es detectada y comparamos la efectividad de varias grabaciones de llamados de Rallus elegans durante intentos de captura. Para cumplir con estos objetivos, experimentamos con el esfuerzo de muestreo, instalamos radios en 12 individuos de Rallus elegans y realizamos monitoreos repetidos utilizando reproducciones de llamados para cada uno de los individuos con radio en Michigan y Ohio, USA durante las temporadas de reproducción del 2020 y 2021. Encontramos que la detectabilidad de Rallus elegans durante 81 monitoreos fue de 0.39, lo cual viola el supuesto del muestreo por distancia mencionado anteriormente y necesita un factor de corrección. La detectabilidad en los segmentos de monitoreo varió entre periodos de muestreo, aunque la detectabilidad se mantuvo relativamente constante, incluso tiempo después de que los muestreos utilizando reproducción de llamados en nuestra área de estudio típicamente terminan (post-periodo de muestreo: 15 de Junio al 10 de Julio), lo que indica que el monitoreo de Rallus elegans puede continuar efectivamente durante un periodo posterior de lo que actualmente ocurre. Encontramos que el llamado tipo gruñido resultó en un número mayor de capturas que el llamado tipo “kek” y que las grabaciones con varios tipos de llamados fueron los más efectivos para la captura de Rallus elegans. Nuestros resultados pueden mejorar los estimativos poblacionales de Rallus elegans e incrementar el éxito durante esfuerzos de captura y pueden informar esfuerzos similares para otras aves sigilosas de pantano.
INTRODUCTION
Visual surveys are commonly used to monitor bird populations, though not all bird species are easily detected visually. For example, many species in the family Rallidae are considered secretive marsh birds (hereafter marsh birds) because of their cryptic nature and preference for dense emergent wetland vegetation. Vocalizations are another common way that many bird species are detected during population monitoring surveys and are typically used for detecting marsh birds (Conway and Gibbs 2005). Call-broadcast surveys, such as those following the Standardized North American Marsh Bird Monitoring (SNAMBM) protocol, can be used to increase detection rates compared to passive listening (Johnson and Dinsmore 1986, Gibbs and Melvin 1993, Conway and Gibbs 2011). During SNAMBM surveys, marsh bird calls are broadcast during three, approximately 2-week survey periods thought to coincide with peak breeding activity. SNAMBM surveys fail to detect individuals that are presumed to be present during > 60% of replicate surveys for most marsh birds, including the King Rail (Rallus elegans) (Conway and Gibbs 2005).
The King Rail is a marsh bird that has experienced extreme population declines since the early 1900s, especially in the northern part of its range where other marsh birds have also declined (Tozer 2016). In areas where King Rails occur at low densities, such as the Midwestern United States, observers who use SNAMBM methodology may frequently fail to detect individuals of this species when they are present (Shirkey et al. 2017). Lower population densities could result in reduced vocal activity by King Rails and other marsh birds (Conway and Gibbs 2005, Schroeder and McRae 2020), which poses challenges to identifying population trends. Particularly in regions where populations are thought to be small, accurate King Rail population density estimates could help identify conservation needs and evaluate management actions.
Distance sampling could be used to accurately estimate King Rail population density by using SNAMBM survey results. Distance sampling allows an investigator to estimate population density based on the rate at which individuals of a focal species are detected at varying distances from observers (Buckland et al. 1993). This method assumes perfect detection of focal individuals at sampling points and nearly perfect detection close to sampling points (Buckland et al. 1993). However, this assumption may be violated for secretive species if detection probability (hereafter detectability) rapidly diminishes with increasing distance from an observer. More specifically, when King Rails silently occur immediately adjacent to the observer (which we have defined as ≤ 30 m herein), the detectability function may lack a shoulder and so violates the shape criterion of distance sampling, thus causing inaccurate density estimation (Buckland et al. 1993). To address this problem, a correction factor, established after dividing the number of individuals detected by the total number present, could allow researchers to correct for those individuals that are present but unavailable for detection due to lack of vocalizations (Andriolo et al. 2006).
An appropriate correction factor for a focal marsh bird species could be estimated by conducting SNAMBM surveys when individuals of that species are known to be nearby. Using radio-telemetry coupled with call-broadcast surveys (Conway et al. 1993, Bui et al. 2015) is one way to accomplish this objective. SNAMBM survey data could likely be used in a distance sampling framework to estimate King Rail population density if an appropriate correction factor for each survey period were used. This could lead to more accurate King Rail population density estimates and demonstrate an approach that could be applied to other marsh birds.
Other important questions regarding King Rail response to audio playback could be answered using radio-telemetry. For example, SNAMBM surveys conducted when radio-tagged individuals are nearby could help optimize future King Rail survey efforts by identifying survey segments that are most associated with King Rail detections throughout the survey season. Call types and call dialects used in SNAMBM surveys are known to affect the rate at which at least one marsh bird, the Hawaiian Moorhen (Gallinula chloropus sandvicensis), responds (DesRochers et al. 2008). Such variation, especially if it leads to detection differences, is important to identify because different audio playback tracks used for SNAMBM surveys could influence estimates of King Rail population density. Further, identifying what audio playback characteristics and time periods are most associated with King Rail detections could improve detectability for this infrequently detected marsh bird (Conway and Gibbs 2011). Additionally, audio playback can attract King Rails to traps (Shirkey et al. 2017), so determining what call types are associated with captures could inform future trapping efforts.
Our overall goal for this study was to address a suite of questions that could improve King Rail population estimates and trapping efficiency. We also intended for our methodology to serve as an example for investigators who are focusing on other marsh birds and/or regions. Specifically, we sought to determine if the distance sampling assumption of perfect detectability immediately adjacent to observers was satisfied, and if not, we aimed to develop correction factors to improve King Rail population estimates throughout the breeding season. We also investigated variables related to environmental factors, time, within-survey details, and attributes of focal individuals that we hypothesized might influence detectability of King Rails during SNAMBM surveys. Additionally, we evaluated how many and which minutes within surveys radio-tagged King Rails tended to be detected and how this varied across the survey season. We predicted overall detectability would decline as the survey season advanced, and that, within surveys, detections would occur primarily during and after the minute when King Rail calls were broadcast. Finally, we sought to determine if trapping success was related to which call type was broadcast. We predicted that a call type presumed to relate to mate attraction (the kek call) would be more conducive to King Rail trapping success than a call type presumed to be more general in its primary function (the grunt call) (Schroeder and McRae 2019). To advance our understanding of the functions of these call types, we also report their use during SNAMBM surveys throughout the survey season.
METHODS
We conducted two separate playback experiments in coastal marshes of southwestern Lake Erie. Fieldwork occurred from 26 April to 10 July in 2020 and 2021, when King Rails were arriving and breeding in our study area (Meanley 1969). We focused efforts at sites where King Rails had been caught in the past in Ohio (Shirkey et al. 2017) and at sites with similar habitat in Ohio and Michigan. To avoid pseudoreplication as described by Kroodsma (1989), we made four audio tracks for each experimental type of playback, and each particular track consisted of calls by individuals not represented in the other tracks. Unless otherwise noted, we randomly determined the order of track types for each bird or trapping site and maintained that order. We randomly selected track types for each playback trial with the constraint that the particular track used previously for that site or individual was not used during the subsequent playback session involving that track type. Audio in all tracks was recorded between 2000 and 2020, during a period when breeding presumably occurred (1 March to 31 August). The tracks originated from xeno-canto (https://xeno-canto.org/), the Macaulay Library (https://www.macaulaylibrary.org), and recordings made by autonomous recording units.
Marked bird experiment
Our objectives for this experiment were to (1) determine if correction factors meant to improve population estimation based on distance sampling were required, and if so, to calculate correction factors, (2) identify factors that may influence King Rail detectability, and (3) describe how many and which minutes King Rails were detected during SNAMBM surveys. Upon capture, we fitted each King Rail with a VHF radio transmitter (ATS – Isanti, Minnesota; model A1050; weight: 2.4 g) using leg-loop harnesses (Rappole and Tipton 1991) consisting of 0.7 mm diameter stretch magic cord (Stretch Magic Inc., Sonoma, California) and initiated the first survey ≥ 24 h later. Prior to each SNAMBM survey, we used handheld radio-telemetry equipment (ATS – Isanti, MN; R2000 and R4000; 3-element folding Yagi antenna) to track individuals. By using radio signal strength and biangulation as an indicator of proximity, we began each SNAMBM survey when we estimated that we were ≤ 30 m from the focal individual. In 2019, we conducted this process on a radio-tagged King Rail, which served solely as practice in preparation for 2020 and 2021. This practice confirmed that we could approach and accurately estimate the distance to a radio-tagged King Rail.
SNAMBM surveys were conducted between 12 May and 10 July in the morning (30 min before sunrise to 3 h after) or in the evening (3 h before sunset to 30 min after), following the SNAMBM protocol (Conway 2011). Each radio-tagged individual (N = 12) was a subject of this experiment, which involved multiple playback sessions throughout the study period for each individual. We attempted to conduct surveys twice per week per individual and never conducted surveys for an individual 2 days in a row. In accordance with the SNAMBM protocol, each survey session began with a 5-min passive period during which no playback was broadcast. This 5-min period was immediately followed by recordings of all focal species that occurred in our study area for which audio playback is required or recommended, including King Rails (Conway 2011). During both the passive and audio broadcast periods, we recorded detections of all focal species and noted which King Rail detections were the radio-tagged bird and which were non-focal King Rails. We were confident that we accurately identified the calls of radio-tagged birds and not nearby King Rails that were not radio-tagged (including two confirmed mates), due to our radio-telemetry methodology and the occasions during surveys (N = 11) that we detected a radio-tagged individual’s mate or other King Rail in a separate, nearby (< 100 m) location compared to where the radio-tagged bird occurred. We used only detections of radio-tagged King Rails for analysis. We used a single primary observer for each survey (multiple observers conducted surveys), though sometimes ≥ 1 additional person was present who may have affected survey results by, for example, alarming focal species or influencing the observer’s surveying process.
We noted each minute when the focal King Rail was detected and what call type they used. Additionally, we noted which call type the focal individual responded to for the King Rail section of playback. If the radio-tagged bird used a call during the period that a King Rail call type was broadcast, or during the 5 s of silence afterward, we considered the focal bird to be responding to that call type. We also noted if the focal bird appeared to move > 5 m during the survey, a common increment used when binning distance, considering that non-movement before detection is an assumption of distance sampling (Buckland et al. 1993). Data were used for analysis only if the focal individual did not move > 20 m during the playback session as estimated by radio-telemetry at the end of the session.
Our SNAMBM surveys used variants of an audio track that we acquired from the SNAMBM survey coordinator (Conway 2011). For each marsh bird species included in this audio track, a 25- to 30-s playback period, including approximately 5 s of silence separating each call bout, was followed by a 25- to 30-s silent period. We modified this track such that eight derived tracks varied only with respect to the King Rail call type instances used (i.e., all the other call types were exactly the same). Four tracks each included a unique 25- to 30-s segment of local King Rail audio clips that originated from the Midwest—specifically, Illinois, Indiana, Michigan, Missouri, and Ohio. Four tracks each included a unique 25- to 30-s segment of foreign King Rail audio clips that originated from outside the Midwest—specifically, Florida, Maine, Maryland, New Jersey, North Carolina, Texas, and Virginia. We alternated foreign and local track types such that a randomly determined track type (e.g., foreign) was used on survey day 1 and was followed by the other type (e.g., local) on survey day 2, and chose a particular track for each survey session as previously described. Each of the King Rail call type bouts within these tracks were 3–6 s in duration and occurred in the same order (grunt, kek, kek-burr). By maintaining the same order and type of calls in this experiment as have been used in SNAMBM surveys completed using the track provided by the program coordinator, we sought to estimate detectability values that are applicable to past and future surveys that used or will use that track or a similarly configured one. We used different kek, grunt, and kek-burr recordings than those used in the trapping experiment to avoid possible negative associations related to the experience of being trapped. We normalized the sound level of audio recordings to -1 dB in Audacity (version 2.3.3). Then we confirmed that relatively little variability existed between King Rail audio segments, and the other segments, regarding maximum sound level as measured 1 m from the handheld speaker (Anker, model AK-A3102031; attached to a Tomameri Mp3 player with an auxiliary cord) that was used in the field (mean = 85.9 dB ± 6.7 [SD]).
Trapping experiment
Our objective for this experiment was to determine which King Rail call types were most effective for capturing King Rails. Trapping was conducted between the nights of 26 April and 7 July in 2020 and 2021. Our trapping attempts used conspecific calls to attract King Rails to walk-in traps, where audio was continuously broadcast between sunset and sunrise. We used the same approach and equipment as Shirkey et al. (2017), and experimentalized trapping efforts by systematically broadcasting two commonly used call types in separate tracks at 54 trapping sites that were > 200 m apart or were visited in different survey years. One track type consisted of grunts; the other consisted of keks (Schroeder and McRae 2019). Within 2 hours of sunrise, we checked traps, noted whether or not a King Rail was captured, and switched track types as previously described if none were captured. The number of trap nights per site varied based on trapping success and presence or absence of King Rail detections in the vicinity. Trapping attempts at particular sites generally ceased after a bird was captured, though occasionally continued at the same site if there was reason to believe that an additional bird was present.
All audio clips used in kek-only and grunt-only tracks consisted of calls recorded where King Rails generally occur solely during the breeding season (specifically, Illinois, Indiana, Kansas, Maryland, Michigan, Missouri, Ohio, or Wisconsin). We used four distinct kek tracks and four distinct grunt tracks. Because our primary overall aim was to catch King Rails, and the experimental tracks had not yet facilitated a capture, on 13 May 2020 we began to include a third track type that had successfully trapped King Rails during previous years. This multi-call track contained three call types (kek, grunt, and kek-burr) (Schroeder and McRae 2019) of unknown origin, and unlike the other track types, was not replicated with four different particular tracks. It was generally incorporated randomly into the sequence at each site, though in some cases was non-randomly chosen when a King Rail was known to be present because of its proven ability to capture King Rails. Although each track was normalized to -0.1 dB in Audacity (version 2.3.3), there was variability among tracks regarding decibel level (measured at 1 m from the speaker) that was broadcast in the field: mean maximum sound level = 80.7 dB ± 12.61 (SD). The multi-call track’s maximum sound level was the greatest (101 dB; next loudest was a grunt track: 98 dB), though the mean sound level for the multi-call track (48 dB) was less than that of two other tracks (a kek [50 dB] and a grunt [57 dB]).
We were approved to conduct this work by Central Michigan University’s IACUC protocol #19-21. We also received approval from the Bird Banding Laboratory (permit # 24149), Ohio (permit # 21-166), and Michigan (permit TE 216).
Statistical analysis
Marked bird experiment: correction factors
We calculated a detectability value for each bird for each survey period when that individual was surveyed ≥ 3 times, a minimum threshold which we assumed would provide a reasonable estimate for each individual. Thus, we were able to produce detectability estimates for SNAMBM periods 2 (15 May to 31 May), 3 (1 June to 14 June), post (15 June to 10 July), and for all periods combined. Periods for our study area were based on Conway (2011). For each individual with sufficient observations within each period, we calculated detectability by dividing the number of surveys during which detection occurred by the total number of surveys that were completed for that individual during that period. Using these values, we calculated mean detectability for each period and divided 1 by the relevant detectability value to determine a correction factor for each period because detectability was imperfect immediately adjacent to the observer. Finally, we calculated detectability for each individual that was surveyed ≥ 3 times across all survey periods (1, 2, 3, and post), determined mean detectability across individuals, and using that value, determined an overall correction factor.
Marked bird experiment: factors affecting detectability
We used AICc (Hurvich and Tsai 1989, Burnham and Anderson 2002) to compare a set of a priori hypotheses to determine which variables, if any, affected King Rail detectability for all surveys that occurred during periods 2, 3, and post. These hypotheses were in the form of generalized linear mixed models (family = binomial; lme4 package [Bates et al. 2015], R v. 4.1.3 [R Core Team 2020]) with a random intercept for bird identity to account for individual differences. We categorized our models into the following groups: environmental, within survey details, temporal, and focal bird attributes (an overview of all models is presented in Table 1). We used 85% confidence intervals to describe variability and identify models with uninformative covariates (Arnold 2010).
We also investigated King Rail detectability within surveys during each survey period. First, we compared the mean number of minutes that King Rails were detected during survey periods 2, 3, and post. To accomplish this, we first omitted individuals from periods if they were not detected at least once therein to reduce zero-inflation. Then we transformed the data via log(x+1) and used a linear mixed model (package = lme4; [Bates et al. 2015]) with a random intercept for bird identity, which accounted for individual differences, to compare detectability for birds that we detected between periods. We used the emmeans package (Lenth 2022) in R v. 4.1.3 (R Core Team 2020) for post-hoc comparisons of means and α = 0.05.
Additionally, we described the probability of detecting individuals during each survey minute in each period. To accomplish this, we first identified which birds were surveyed ≥ 3 times in a particular period and calculated a detectability value for each bird during each minute in that period, thus weighting individuals equally. We used these values to calculate mean detectability for each minute during each period. Finally, we used these means to calculate mean detectability during the silent period, the pre-King Rail period (after silent, before King Rail playback), and the King Rail playback and after period. Similarly, we determined mean detectability within the King Rail minute across survey periods, though the survey segments analyzed were grunt, kek, kek-burr, and silent period (30 s). We also summarized which call types we documented King Rails using during each survey period.
Trapping experiment
We descriptively summarized overall trapping effort and success in relation to each track type that was broadcast. We also report trapping success for each track type during initial trapping nights at sites to remove effects of the previous playback sessions. To account for bias toward using the multi-call track, we summarized trapping efforts when King Rails were not caught during the first night. This excluded several cases when the multi-call track was used because a King Rail was known to be present. Finally, we summarized which audio tracks occurred the night before each successful trapping night to indicate which track types were unsuccessful when there was an increased likelihood that a King Rail was present.
RESULTS
Marked bird experiment
We conducted 83 surveys for 12 radio-tagged birds, of which 81 occurred in periods 2, 3, and post. The number of surveys included for detectability calculation in each period were as follows: 17 (for 5 birds) in period 2, 22 (for 6 birds) in period 3, 26 (for 6 birds) in post, and 81 (for 11 birds) overall. All initial detections during surveys were due to the focal bird vocalizing.
We estimated that, across all surveys conducted, the focal bird moved 5–20 m during 40% (33/83) of surveys and < 5 m during the remainder. Our estimate of mean distance to the birds that we surveyed was 16.4 ± 0.7 m (SE).
Correction factors
Detectability estimates were 0.42 ± 0.2 (SE) for period 2, 0.30 ± 0.12 for period 3, 0.53 ± 0.18 for the post period, and 0.39 ± 0.1 (SE) for overall. These imperfect detectability values necessitated calculation of correction factors for use in distance sampling analyses. The corresponding correction factors were 2.40 for period 2, 3.37 for period 3, 1.90 for the post period, and 2.58 for overall.
Factors affecting detectability
All the model types, except for temporal, had at least one particular model that was ranked better than the null model (Table 2). However, Akaike weights of the null model and those ranked above it were similar (0.09–0.17) (Table 2).
We report parameter estimates for the four top models, each of which included a single variable: cloud cover, observer, trial number, and sex (Table 3). In summary, greater cloud cover, having ≥ 1 other person with the observer, surveying a bird ≤ 5 times, and surveying a male bird all increased detectability.
We found that mean number of minutes when a King Rail was detected varied by period (F[2, 39.2] = 3.3, P = 0.05, df via Satterthwaite’s method). Post-hoc comparisons indicated that, after P value adjustment via Tukey’s method, King Rails were detected during more minutes in period 2 than in period 3. The estimated difference between these periods, based on log-transformed values, was 0.25 (95% CI = 0.05–0.45, t = 2.4, P = 0.05, df = 39.9, via the Kenward-Roger method). The mean number of minutes when a King Rail was detected varied by 1.77 between these periods using untransformed data: period 2 = 2.64 ± 0.77 (SE) minutes; period 3 = 0.87 ± 0.42 min.
Mean detectability was low (≤ 0.31) within each survey minute across all survey periods, regardless of whether or not audio playback occurred (Fig. 1). Although variability was high and estimates of population mean detectability tended to overlap between periods and survey segments, playback tended to increase the likelihood that King Rails were detected. In period 3, the minute of King Rail playback was the only minute that did not include zero in the population mean estimate (Fig. 1). In period 2, playback of King Rail calls (and calls after them) increased mean detectability to 0.20–0.24 (95% CI) compared to population mean estimates that included zero for the other survey segments (Fig. 2). We conducted two surveys when the focal individual was known to be incubating eggs, but neither bird (a male and a female) was detected.
Within the minute that King Rail calls were broadcast, there was little variability regarding when individuals tended to be detected between periods or segments. Mean detectability during these segments ranged from 0.05 to 0.15 in period 2 (highest during the 30 s after segment), 0.04 to 0.18 in period 3 (highest during kek-burr segment), and 0 to 0.07 in the post period (tied for highest during the kek and kek-burr segments). King Rail response varied regarding which call types were used during the different periods. Namely, in period 2, individuals tended to use both kek and grunt calls during surveys, though in subsequent periods, grunt calls were generally the only call type used (Fig. 3).
Trapping experiment
During 2020 and 2021, we successfully trapped ≥ 1 King Rail during 11 of 424 (2.6%) trapping nights between 10 May and 16 June, with a mean capture date of 19 May. During both years, trapping success rate was 2.6% (2020: 5 of 193 trap nights; 2021: 6 of 231 trap nights). The within-site mean number of trapping nights before capture was 4.3 ± 1.4 (SE), and the overall mean was 7.9 ± 0.7 (SE). Five of the 11 successful trapping nights occurred during the first night of trapping at a site. The multi-call track facilitated 8 of 11 captures (6.3% success rate: 8/126), the grunt track type facilitated the remaining three captures (a different track was used for each; 2.0% success rate: 3/149), and kek tracks were unsuccessful (0/149). Success rates for initial trapping nights were 20% for multi-call (4/20), 9.1% for grunt (1/11), and 0% for kek (0/23). Each track type was used during ≥ 1 night before a successful trapping night: multi-call = 3, grunt = 2, and kek = 1. The multi-call track and grunt tracks both facilitated capture of at least one male and one female King Rail. The grunt tracks facilitated King Rail captures only in May, whereas the multi-call track did so in both May and June.
The study design displayed bias toward playing the multi-call track when King Rails were known to be present at a site due to previous success of this track. Four of the eight instances when the multi-call track facilitated capture occurred during the first night audio was played at a site. Upon excluding all five of the instances that a track facilitated capture during the first trapping night, we found that the grunt and multi-call track types achieved more similar success rates (1.4% and 3.3%, respectively).
DISCUSSION
We found imperfect detectability immediately adjacent to observers, so concluded that correction factors could improve population density estimates for King Rails, and perhaps other marsh birds, via distance sampling. Progress has been made toward using SNAMBM survey data to model King Rail occupancy (Glisson et al. 2015, Stevens and Conway 2019, 2020), though little advancement has been made toward estimating population density for this species, likely due to its low abundance, occupancy rate, and detectability in many regions (Steidl et al. 2013). Population density has been estimated for other marsh birds (e.g., Harms and Dinsmore 2012, Wiest et al. 2016, 2019, Vanausdall et al. 2022) despite the high potential for these often-secretive species to remain undetected at or immediately adjacent to the observer, which is a violation of a distance sampling assumption (Buckland et al. 1993). N-mixture models can produce reliable population density estimates via repeated surveys at locations, though are known to overestimate population size where densities are low (Neubauer et al. 2022), as is often true for King Rails. By using a correction factor, a distance sampling approach could feasibly yield an accurate King Rail population density estimate without employing the double-observer method (Thomas et al. 2010), similar to approaches used for taxa such as cetaceans (e.g., Andriolo et al. 2006) that often remain undetected at or immediately adjacent to an observer. For example, a population monitoring effort within our study area that followed the SNAMBM protocol (Conway 2011), which generates an estimate of 1.2 King Rails per square kilometer of suitable habitat, could use our overall correction factor (2.58) to adjust that estimate to 3.1 individuals per square kilometer (1.2 x 2.58) and so account for the inaccurate assumption of perfect detectability during the modeling process. Note, however, that our correction factors should be used with caution given our small sample size. Surveyors who follow the SNAMBM protocol generally conduct surveys at a location once during each of the three, approximately 2-week periods specific to their region, and sometimes use these data in an occupancy modeling framework (Conway 2011). Our overall correction factor should not be used to correct a population density estimate for an area consisting of survey locations for which the survey period with the maximum number of King Rail detections was intentionally chosen for analysis after all three surveys in a season were completed because this could overestimate density. Rather, random selection of which survey to use at each location, or use of the same survey period for all locations, would avoid bias associated with including only surveys conducted when King Rails at a location happened to be most detectable. If the same survey period is used for all locations to estimate population density of an area, then using a correction factor tailored to that survey period could better account for changes in detectability that may occur as a function of breeding stage. Our correction factors are likely less accurate outside our study area given known variability regarding King Rail calling rate based on conspecific density (Schroeder and McRae 2020), which may be exceptionally low in our study area (Bolenbaugh et al. 2012). Given that we radio-tagged only three females, and that we generally had a small sample size, future studies using the same approach could advance our efforts to calculate useful correction factors by radio-tagging more King Rails, in general, and more females, in particular.
We found no evidence that detectability varied based on distance within our immediately-adjacent-to-the-observer (≤ 30 m) category. This indicates that the category could well-represent the initial bin, wherein detectability should be at or near 100% for distance sampling (Buckland et al. 1993), for establishing how King Rail detectability declines with increasing distance from an observer. Our overall detectability estimate (0.39) was similar to those determined by others who, unlike us, used a repeated sampling, occupancy modeling framework that did not involve radio-tagged birds: 0.35 and 0.43 in Missouri/Illinois (Darrah and Krementz 2009), 0.39 in Arkansas (Budd and Krementz 2011), and 0.48 in Louisiana (Pierluissi and King 2008). These detectability estimates may have been higher had King Rails been radio-tagged and approached to within 30 m as we did. For example, Kane (2020) conducted a King Rail occupancy modeling investigation in our study area and estimated detectability during SNAMBM surveys at 0.11. If detectability as derived by occupancy modeling is a consistent fraction of detectability as derived by radio-tagging and surveying nearby King Rails, then correction factors could be calculated and applied to distance sampling wherever a SNAMBM-based occupancy approach is used. However, the broader relationship between these distinct kinds of detection probabilities requires establishment. Bui et al. (2015) employed an approach similar to ours during their study that focused on radio-tagged Ridgway’s Rails (Rallus obsoletus) in California, which, in accord with our study, indicated that focal birds moved relatively little during surveys and produced a detectability estimate (0.41) similar to what we found (0.39). This may provide some support for the generality of our results, though Bui et al. (2015) conducted their surveys within 200 m of radio-tagged individuals rather than within 30 m.
We found little evidence that the variables we investigated had a strong effect on King Rail detectability (Table 2), which could simplify population monitoring and modeling efforts for this species. However, greater cloud cover was correlated with increased detectability (Table 3), which agrees with the finding by Hansen (2019) that King Rails were more detectable when barometric pressure was lower. Similarly, we found at least a slight decline in detectability with increasing trial number (perhaps due to habituation [Harris and Haskell 2013]) and at least a slight increase in detectability when the focal bird was male, as well as during surveys when ≥ 1 person was present. It is possible that nesting status (Legare et al. 1999, Robertson and Olsen 2014) and daily incubating patterns (Clauser and McRae 2017) influenced the differential response between sexes, though sampling more females would have increased confidence in this effect. It is unclear whether having another person present increased true detections (e.g., more observer noise could lead to more calling) or false positives (e.g., a companion could increase observer confidence in an erroneous detection). Our finding that the origin of King Rail calls (foreign or local) did not affect King Rail detectability during SNAMBM surveys is similar to the finding by Conway et al. (2020) regarding a closely related species, the Clapper Rail (Rallus crepitans), and should be considered when creating audio tracks for surveys.
Our results suggest that audio playback increased King Rail detectability (Figs. 1 and 2), which has also been found for other marsh birds (Tozer et al. 2017). Although detectability was highest within surveys during and after King Rail playback in period 2 (Fig. 2), we did not find evidence that detectability at the survey scale declined after the end of the SNAMBM survey season (Conway 2011). This indicates that King Rail monitoring efforts in our study area could effectively extend beyond the current end date (14 June) until at least 10 July, though observers may have fewer opportunities to detect this species within surveys after period 2. Moreover, chicks in our study area may tend to begin hatching in early July (personal observation, D.B.) and so affect detectability thereafter. Shifting the SNAMBM survey period back 2 weeks in our study area could better align efforts with when King Rails are present and detectable, and reduce encounters of migratory Soras (Porzana carolina) and Virginia Rails (Rallus limicola) (Hansen 2019, Hengst 2021). Other investigators have also found that extending SNAMBM surveys later into the season could be beneficial (Harms and Dinsmore 2014, Rehm and Baldassarre 2007). King Rails responded with kek calls less after survey period 2 (Fig. 3), which may correspond to a transition from attempts at mate attraction (Schroeder and McRae 2019). Conversely, grunt calls were used at similar rates in all survey periods, which is in accord with calling activity described by Schroeder and McRae (2020).
Our preliminary findings indicate that King Rail trapping success may be better achieved by broadcasting grunt calls rather than kek calls. Schroeder and McRae (2019) associated grunt calls with territorial intrusions, which our playback may have effectively simulated when using grunt calls but not when using kek calls. Even after controlling for study design bias, it appears that using three call types (kek, grunt, and kek-burr) was more effective at facilitating trapping success than was using either kek or grunt calls alone. However, it is also possible that the improved performance of the multi-call track was due at least in part to inclusion of the kek-burr call type, which we did not broadcast alone. Our ability to draw inferences regarding the effectiveness of different call types for trapping King Rails was limited due to our approach. Future investigators could improve upon our approach by strictly adhering to a random presentation of audio tracks during trapping attempts and by better standardizing the sound level of tracks, though sound level variability may be less important (Conway et al. 2004). Shirkey et al. (2017) used the same traps and approach as we did and found a similar trapping success rate (0.02) as ours (0.03), though they used predominantly the multi-call track described herein rather than systematically varying the presentation of track types. Given that our average number of trap nights at sites until King Rail capture was four, and trapping success often occurred during the first trapping night (45%; 5/11), future investigators may benefit from shifting traps to new locations frequently to increase the likelihood of encountering a King Rail.
In summary, we found that imperfect King Rail detectability in our study area necessitates use of a correction factor to improve population estimates made via distance sampling. Moving beyond occupancy trends toward estimating accurate regional and local population densities would help identify where the best opportunities exist to facilitate recovery of King Rail populations. Further, we found that King Rail detectability tended to remain relatively constant in a variety of contexts and that surveys for this species could likely continue at least until 10 July in our study area. Using multi-call tracks that include the grunt call may optimize efforts to trap King Rails during future research efforts, though further study is warranted. Overall, our findings could improve King Rail population estimates, increase trapping efficiency, and advance similar efforts for other marsh birds.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
Funding acquisition – T.G., B.S., J.S., D.B. Project design – D.B., B.S., T.G., J.S. Fieldwork – D.B., B.S. Analysis – D.B. Writing – D.B. Editing – D.B., B.S., T.G., J.S.
ACKNOWLEDGMENTS
We appreciate the funding support provided by the Upper Mississippi/Great Lakes Joint Venture, The Wetland Foundation, and the Society of Wetland Scientists. Central Michigan University also provided support via the College of Science and Engineering, Department of Biology, and Institute for Great Lakes Research (contribution number 186). This research was also supported by the Earth and Ecosystem Science PhD program at CMU and by Winous Point Marsh Conservancy. We are grateful for the fieldwork done by the following people: T. McClinton, L. Wallace, R. Bealer, T. King, G. Ravary, B. Abt, and J. Schmit.
DATA AVAILABILITY
Data and code are available at https://doi.org/10.5281/zenodo.7464763
LITERATURE CITED
Andriolo, A., C. C. A. Martins, M. H. Engel, J. L. Pizzorno, S. Más-Rosa, A. C. Freitas, and M. E. Morete. 2006. The first aerial survey to estimate abundance of humpback whales (Megaptera novaeangliae) in the breeding ground off Brazil (Breeding Stock A). Journal of Cetacean Research and Management 8(3):307-311. https://doi.org/10.47536/jcrm.v8i3.728
Arnold, T. W. 2010. Uninformative parameters and model selection using Akaike’s Information Criterion. Journal of Wildlife Management 74:1175-1178. https://doi.org/10.1111/j.1937-2817.2010.tb01236.x
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67:1-48. https://doi.org/10.18637/jss.v067.i01
Bolenbaugh, J. R., T. Cooper, R. S. Brady, K. L. Willard, and D. G. Krementz. 2012. Population status and habitat associations of the King Rail in the Midwestern United States. Waterbirds 35:535-545. https://doi.org/10.1675/063.035.0404
Buckland, S. T., D. R. Anderson, K. P. Burnham, and J. L. Laake. 1993. Distance sampling: estimating abundance of biological populations. Chapman and Hall, London, UK.
Budd, M. J., and D. G. Krementz. 2011. Status and distribution of breeding secretive marshbirds in the Delta of Arkansas. Southeastern Naturalist 10:687-702. https://doi.org/10.1656/058.010.0408
Bui, T. D., J. Y. Takekawa, C. T. Overton, E. R. Schultz, J. M. Hull, and M. L. Casazza. 2015. Movements of radio-marked California Ridgway’s Rails during monitoring surveys: implications for population monitoring. Journal of Fish and Wildlife Management 6:227-237. https://doi.org/10.3996/092014-JFWM-069
Burnham, K. P., and D. R. Anderson. 2002. Model selection and multimodel inference: a practical information-theoretic approach. Springer-Verlag, New York, USA.
Clauser, A. J., and S. B. McRae. 2017. Plasticity in incubation behavior and shading by King Rails (Rallus elegans) in response to temperature. Journal of Avian Biology 48:479-488. https://doi.org/10.1111/jav.01056
Conway, C. J. 2011. Standardized North American marsh bird monitoring protocol. Waterbirds 34:319-346. https://doi.org/10.1675/063.034.0307
Conway, C. J., W. R. Eddleman, S. H. Anderson, and L. R. Hanebury. 1993. Seasonal changes in Yuma Clapper Rail vocalization rate and habitat use. Journal of Wildlife Management 57:282-290. https://doi.org/10.2307/3809425
Conway, C. J., and J. P. Gibbs. 2005. Effectiveness of call-broadcast surveys for monitoring marsh birds. Auk 122:26-35. https://doi.org/10.1093/auk/122.1.26
Conway, C. J., and J. P. Gibbs. 2011. Summary of intrinsic and extrinsic factors affecting detection probability of marsh birds. Wetlands 31:403-411. https://doi.org/10.1007/s13157-011-0155-x
Conway, C. J., C. P. Nadeau, and M. Conway. 2020. Broadcasting regional call dialects has little influence on the effectiveness of call-broadcast surveys for marsh birds. Wetlands 40(6):2055-2059. https://doi.org/10.1007/s13157-020-01367-5
Conway, C. J., C. Sulzman, and B. E. Raulston. 2004. Factors affecting detection probability of California Black Rails. Journal of Wildlife Management 68:360-370. https://doi.org/10.2193/0022-541X(2004)068[0360:FADPOC]2.0.CO;2
Darrah, A. J., and D. G. Krementz. 2009. Distribution and habitat use of King Rails in the Illinois and Upper Mississippi River valleys. Journal of Wildlife Management 73:1380-1386. https://doi.org/10.2193/2008-561
DesRochers, D. W., H. K. W. Gee, and J. M. Reed. 2008. Response of Hawaiian Moorhens to broadcast of conspecific calls and a comparison with other survey methods. Journal of Field Ornithology 79:448-457. https://doi.org/10.1111/j.1557-9263.2008.00190.x
Gibbs, J. P., and S. M. Melvin, 1993. Call-response surveys for monitoring breeding waterbirds. Journal of Wildlife Management 57:27-34. https://doi.org/10.2307/3808996
Glisson, W. J., C. J. Conway, C. P. Nadeau, K. L. Borgmann, and T. A. Laxson. 2015. Range-wide wetland associations of the King Rail: a multi-scale approach. Wetlands 35:577-587. https://doi.org/10.1007/s13157-015-0648-0
Hansen, J. M. 2019. Survey methods and habitat associations of secretive marsh birds in coastal wetlands of the western Lake Erie basin. Ohio State University, Columbus, Ohio, USA.
Harms, T. M., and S. J. Dinsmore. 2012. Density and abundance of secretive marsh birds in Iowa. Waterbirds 35:208-216. https://doi.org/10.1675/063.035.0203
Harms, T. M., and S. J. Dinsmore. 2014. Influence of season and time of day on marsh bird detections. Wilson Journal of Ornithology 126:30-38. https://doi.org/10.1676/13-150.1
Harris, J. B. C., and D. G. Haskell. 2013. Simulated birdwatchers’ playback affects the behavior of two tropical birds. PLoS ONE 8:e77902. https://doi.org/10.1371/journal.pone.0077902
Hengst, N. M. 2021. Movements and habitat relationships of Virginia Rails and Soras within impounded coastal wetlands of Northwest Ohio. Ohio State University, Columbus, Ohio, USA.
Hurvich, C. M., and C.-L. Tsai. 1989. Regression and time series model selection in small samples. Biometrika 76:297-307. https://doi.org/10.1093/biomet/76.2.297
Johnson, R. R., and J. J. Dinsmore. 1986. Habitat use by breeding Virginia Rails and Soras. Journal of Wildlife Management 50:387-392. https://doi.org/10.2307/3801092
Kane, M. 2020. Occupancy and relative habitat suitability of King Rails in the Midwest. Central Michigan University, Mt. Pleasant, Michigan, USA.
Kroodsma, D. E. 1989. Suggested experimental designs for song playbacks. Animal Behaviour 37:600-609. https://doi.org/10.1016/0003-3472(89)90039-0
Legare, M. L., W. R. Eddleman, P. A. Buckley, and C. Kelly. 1999. The effectiveness of tape playback in estimating Black Rail density. Journal of Wildlife Management 63:116-125. https://doi.org/10.2307/3802492
Lenth, R. V. 2022. emmeans: estimated marginal means, aka least-squares means. R package. Version 1.8.1-1.
Meanley, B. 1969. Natural history of the King Rail. North American Fauna 67. https://doi.org/10.5962/bhl.title.86991
National Weather Service. 2013. Estimating wind speed and sea state.
Neubauer, G., A. Wolska, P. Rowinski, and T. Wesolowski. 2022. N-mixture models estimate abundance reliably: a field test on Marsh Tit using time-for-space substitution. Ornithological Applications 124:duab054. https://doi.org/10.1093/ornithapp/duab054
Pierluissi, S., and S. King. 2008. Relative nest density, nest success, and site occupancy of King Rails in southwestern Louisiana rice fields. Waterbirds 31:530-541. https://doi.org/10.1675/1524-4695-31.4.530
R Core Team. 2020. R: a language and environment for statistical computing.
Rappole, J. H., and A. R. Tipton. 1991. New harness design for attachment of radio transmitters to small passerines. Journal of Field Ornithology 62:335-337.
Rehm, E. M., and G. A. Baldassarre. 2007. Temporal variation in detection of marsh birds during broadcast of conspecific calls. Journal of Field Ornithology 78:56-63. https://doi.org/10.1111/j.1557-9263.2006.00085.x
Robertson, E. P., and B. J. Olsen. 2014. Density, sex, and nest stage affect rail broadcast survey results. Journal of Wildlife Management 78:1293-1301. https://doi.org/10.1002/jwmg.769
Schroeder, K. M., and S. B. McRae. 2019. Vocal repertoire of the King Rail (Rallus elegans). Waterbirds 42:154-167. https://doi.org/10.1675/063.042.0202
Schroeder, K. M., and S. B. McRae. 2020. Automated auditory detection of a rare, secretive marsh bird with infrequent and acoustically indistinct vocalizations. Ibis 162:1033-1046. https://doi.org/10.1111/ibi.12805
Shirkey, B. T., J. W. Simpson, and M. A. Picciuto. 2017. King Rail (Rallus elegans) trapping efficiency and detection techniques in southwestern Lake Erie coastal marshes, USA. Waterbirds 40:69-73. https://doi.org/10.1675/063.040.0110
Steidl, R. J., C. J. Conway, and A. R. Litt. 2013. Power to detect trends in abundance of secretive marsh birds: effects of species traits and sampling effort. Journal of Wildlife Management 77:445-453. https://doi.org/10.1002/jwmg.505
Stevens, B. S., and C. J. Conway. 2019. Predicting species distributions: unifying model selection and scale optimization for multi-scale occupancy models. Ecosphere 10:e02748. https://doi.org/10.1002/ecs2.2748
Stevens, B. S., and C. J. Conway. 2020. Predictive multi-scale occupancy models at range-wide extents: effects of habitat and human disturbance on distributions of wetland birds. Diversity and Distributions 26:34-48. https://doi.org/10.1111/ddi.12995
Thomas, L., S. T. Buckland, E. A. Rexstad, J. L. Laake, S. Strindberg, S. L. Hedley, J. Rb. Bishop, T. A. Marques, and K. P. Burnham. 2010. Distance software: design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology 47:5-14. https://doi.org/10.1111/j.1365-2664.2009.01737.x
Tozer, D. C. 2016. Marsh bird occupancy dynamics, trends, and conservation in the southern Great Lakes basin: 1996 to 2013. Journal of Great Lakes Research 42:136-145. https://doi.org/10.1016/j.jglr.2015.10.015
Tozer, D. C., C. M. Falconer, A. M. Bracey, E. E. Gnass Giese, G. J. Niemi, R. W. Howe, T. M. Gehring, and C. J. Norment. 2017. Influence of call broadcast timing within point counts and survey duration on detection probability of marsh breeding birds. Avian Conservation and Ecology 12(2):8. https://doi.org/10.5751/ACE-01063-120208
Vanausdall, R. A., T. M. Harms, and S. J. Dinsmore. 2022. Marsh bird response to restored shallow lakes: implications for future management. Wildlife Society Bulletin 46:e1296. https://doi.org/10.1002/wsb.1296
Wiest, W. A., M. D. Correll, B. G. Marcot, B. J. Olsen, C. S. Elphick, T. P. Hodgman, G. R. Guntenspergen, and W. G. Shriver. 2019. Estimates of tidal-marsh bird densities using Bayesian networks. Journal of Wildlife Management 83:109-120. https://doi.org/10.1002/jwmg.21567
Wiest, W. A., M. D. Correll, B. J. Olsen, C. S. Elphick, T. P. Hodgman, D. R. Curson, and W. G. Shriver. 2016. Population estimates for tidal marsh birds of high conservation concern in the northeastern USA from a design-based survey. Condor 118:274-288. https://doi.org/10.1650/CONDOR-15-30.1
Fig. 1
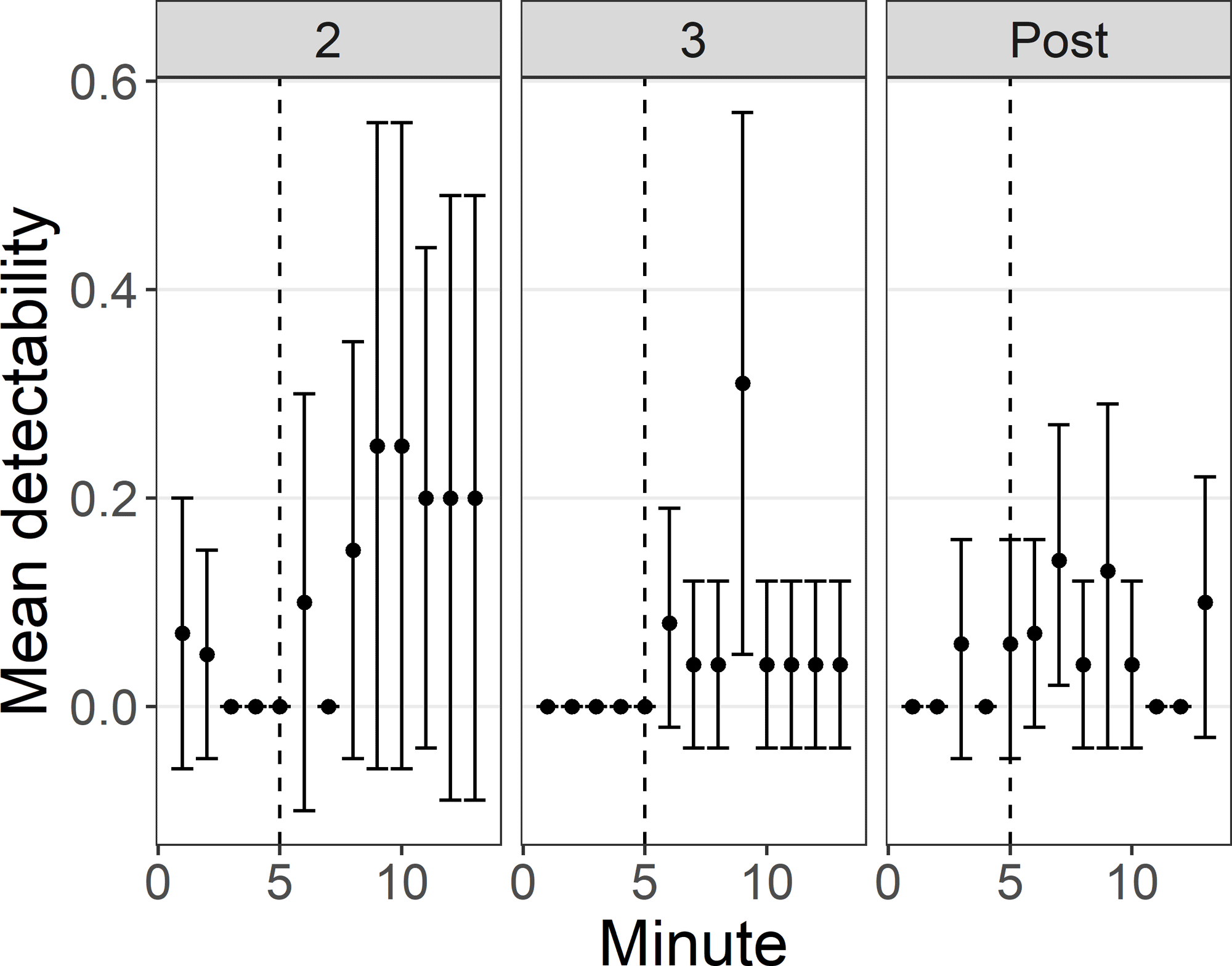
Fig. 1. Mean King Rail detectability in each survey minute during periods 2, 3, and post. The area to the right of the vertical dashed line in each panel shows the minutes when playback occurred. Minute 6 = Least Bittern, minute 7 = Sora, minute 8 = Virginia Rail, minute 9 = King Rail, minute 10 = American Bittern, minute 11 = Common Gallinule, minute 12 = American Coot, minute 13 = Pied-billed Grebe. Bars are 95% confidence intervals.
Fig. 2
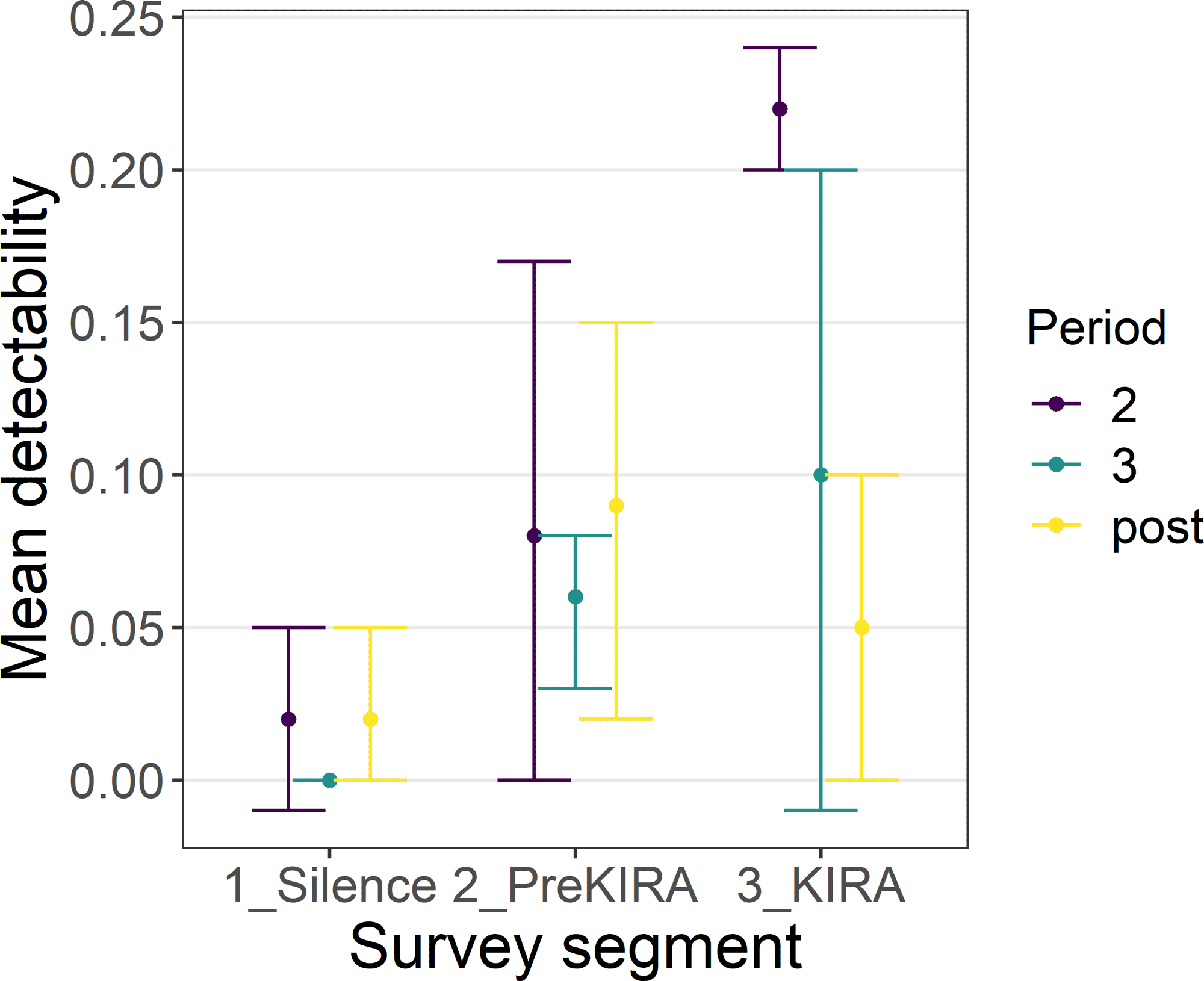
Fig. 2. Mean King Rail detectability during each period (2, 3, and post) across survey segments. 1_Silence = first 5 minutes of survey track, before any playback was broadcast. 2_PreKIRA = 3 minutes following silent period, when Least Bittern, Sora, and Virginia Rail calls were broadcast. 3_KIRA = final 5 minutes, when King Rail, American Bittern, Common Gallinule, American Coot, and Pied-billed Grebe calls were broadcast. Bars are 95% confidence intervals.
Fig. 3
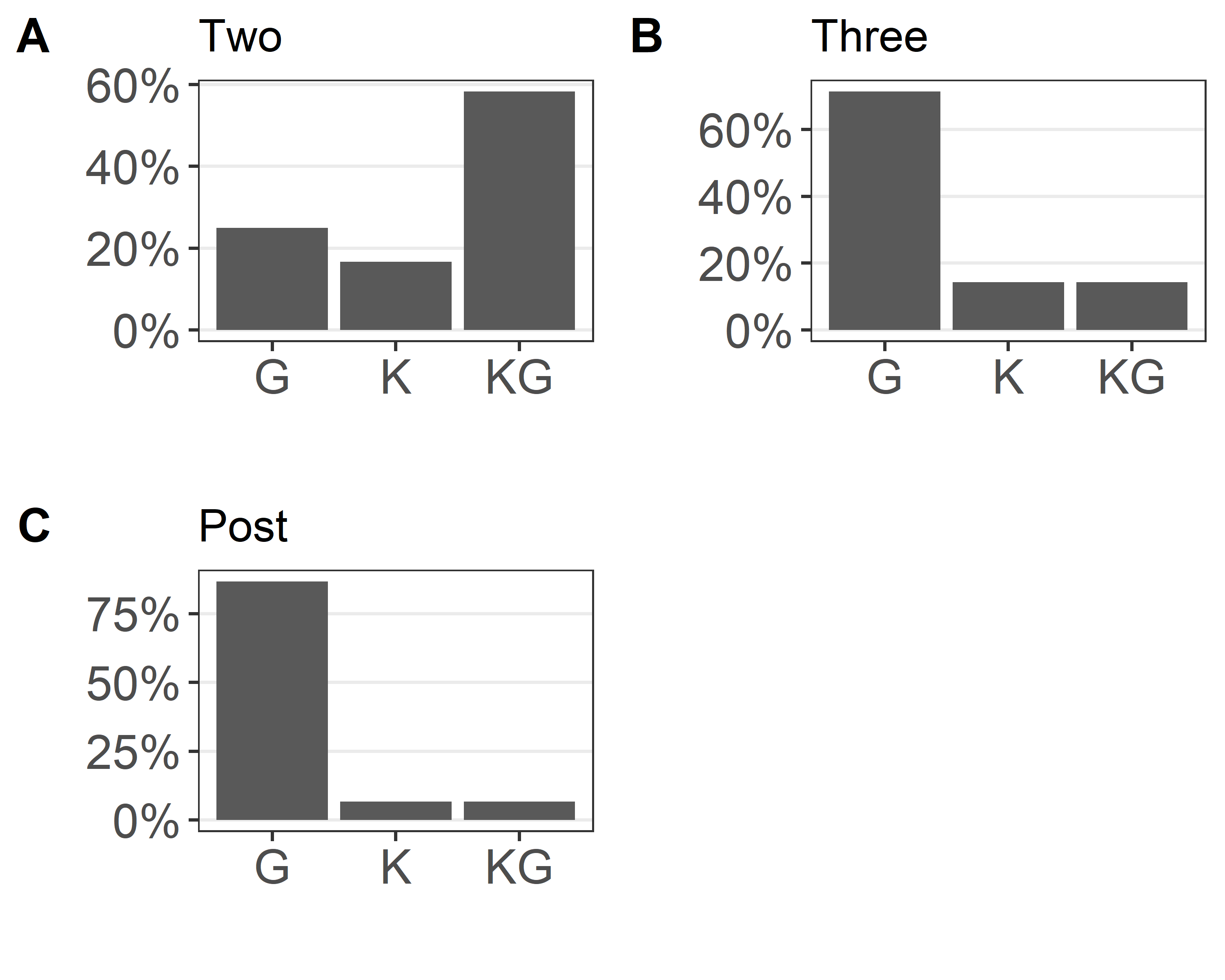
Fig. 3. Percentage of surveys when only a grunt (G), only a kek (K), or a kek and a grunt (KG) were used by the focal King Rail during each survey period. These data reflect surveys when a King Rail was detected via a kek and/or grunt call in periods 2 (A), 3 (B), and post (C). N = 12 in period 2, N = 7 in period 3, N = 15 in the post period. Note that the scales differ.
Table 1
Table 1. Candidate model set used to evaluate effects of variables on King Rail detectability. Emod = environmental model, Wmod = within survey details model, Tmod = temporal model, Bmod = focal bird attributes model. Cloud cover was measured as follows: 0 represents < 10%, 1 represents ≥ 10% and < 50%, 2 represents ≥ 50% and < 80%, 3 represents ≥ 80%. Noise was measured subjectively as follows: 0 = none, 1 = faint, 2 = moderate, 3 = loud. Period refers to Standardized North American Marsh Bird Monitoring period. The Beaufort scale was used as described by the National Weather Service (2013).
| Model name | Details | Model name | Details | ||||||
| Emod1 | Temperature (°C) + cloud cover (0 to 1 or 2 to 3) | Tmod4 | Year (2020 or 2021) | ||||||
| Emod2 | Noise (0 to 1 or 2 to 3) + wind (Beaufort: 0 to 1, 2 to 3, or 4 to 5) | Tmod5 | Morning/evening | ||||||
| Emod3 | Temperature (°C) | Wmod1 | Number of observers (1 or > 1) | ||||||
| Emod4 | Wind (Beaufort: 0 to 1, 2 to 3, or 4 to 5) | Wmod2 | Track type (foreign/local recording) | ||||||
| Emod5 | Noise (0 to 1 or 2 to 3) | Wmod3 | Trial number (for individual; 1 to 5 or after 5) | ||||||
| Emod6 | Cloud cover (0 to 1 or 2 to 3) | Wmod4 | Trial number (for individual; 1 to 5 or after 5)*Track type (foreign/local recording) | ||||||
| Tmod1 | Period (2 or after 2) + morning/evening | Bmod1 | Sex (determined genetically) | ||||||
| Tmod2 | Period (2 or after 2) | Bmod2 | Distance to bird (m; estimated) | ||||||
| Tmod3 | Minutes from sunrise/sunset | Null | No predictors | ||||||
Table 2
Table 2. Ranking of models by AICc. Note that Emod1 did not converge, so it is not listed. The null model and all those that were ranked better than it are bolded. Each model includes a random intercept. Emod = environmental model, Wmod = within survey details model, Tmod = temporal model, Bmod = focal bird attributes model.
| Model details | Model name | K | AICc | Δ AICc | Akaike weight | Log likelihood | |||
| Cloud cover | Emod6 | 3 | 95.76 | 0 | 0.17 | -44.72 | |||
| Observer | Wmod1 | 3 | 96.11 | 0.35 | 0.14 | -44.90 | |||
| Trial number | Wmod3 | 3 | 96.51 | 0.76 | 0.11 | -45.10 | |||
| Sex | Bmod1 | 3 | 96.60 | 0.84 | 0.11 | -45.14 | |||
| No predictors | Null | 2 | 97.00 | 1.25 | 0.09 | -46.43 | |||
| Temperature | Emod3 | 3 | 97.72 | 1.96 | 0.06 | -45.70 | |||
| Distance | Bmod2 | 3 | 98.12 | 2.37 | 0.05 | -45.90 | |||
| Start | Tmod3 | 3 | 98.37 | 2.62 | 0.05 | -46.03 | |||
| Noise | Emod5 | 3 | 98.79 | 3.03 | 0.04 | -46.24 | |||
| Year | Tmod4 | 3 | 98.91 | 3.16 | 0.03 | -46.30 | |||
| Period | Tmod2 | 3 | 99.03 | 3.27 | 0.03 | -46.36 | |||
| Morning or evening | Tmod5 | 3 | 99.16 | 3.40 | 0.03 | -46.42 | |||
| Track type | Wmod2 | 3 | 99.16 | 3.40 | 0.03 | -46.42 | |||
| Trial number*track type | Wmod4 | 5 | 100.24 | 4.48 | 0.02 | -44.72 | |||
| Period + start | Tmod1 | 4 | 100.50 | 4.74 | 0.02 | -45.98 | |||
| Wind | Emod4 | 4 | 100.56 | 4.80 | 0.02 | -46.02 | |||
| Noise + wind | Emod2 | 5 | 102.38 | 6.62 | 0.01 | -45.79 | |||
Table 3
Table 3. Parameter estimates of the top models. Variability is reported in parentheses as 85% confidence intervals pertaining to King Rail detectability.
| Model details | Model name | Variable | Parameter estimate |
| Cloud cover | Emod6 | Sky_2to3 Intercept |
1.31 (0.23 to 2.39) 1.31 |
| Observer | Wmod1 | Observer_Single | -1.40 (-2.64 to -0.16) |
| Intercept | 0.36 | ||
| Trial number | Wmod3 | TrialNumber_6to15 Intercept |
-1.21 (-2.34 to -0.08) -0.37 |
| Sex | Bmod1 | Sex_Male Intercept |
1.91 (0.18 to 3.64) -1.98 |