







The following is the established format for referencing this article:
Sanders, F. J., A. D. Smith, J. M. Thibault, D. L. Carter, M. C. Handmaker, and F. M. Smith. 2023. South Atlantic Bight – a final stop for Ruddy Turnstones migrating to the Arctic. Journal of Field Ornithology 94(2):5.ABSTRACT
Migratory stopover sites are of high conservation concern especially those sites where shorebirds concentrate in large numbers to acquire fat reserves to fuel continued flight to breeding grounds. Many shorebirds use only a few stopover sites during northward migration to Arctic breeding grounds, thus identifying important locations, migration chronology, and flight routes are priority research topics to inform effective management strategies. We examined Ruddy Turnstones (Arenaria interpres) route and timing to Arctic breeding grounds from an island in South Carolina in the heart of the South Atlantic Bight using the Motus Wildlife Tracking System. Ruddy Turnstones leaving South Carolina did not migrate northward up the Atlantic coast but migrated inland through the Great Lakes Basin. Most Ruddy Turnstones did not make a stop in the Great Lakes Basin, thus making South Carolina the last presumed stopover before reaching Arctic habitats. Like other shorebird migration studies, most of the Ruddy Turnstones made use of tailwinds at departure and ground speeds were positively correlated with tailwind support. Future conservation planning for Ruddy Turnstones must consider the varied migratory routes and strategies of this declining shorebird species. This research also demonstrates the usefulness of Motus for tracking the movement of smaller shorebirds and the potential for strategic expansion of the Motus network to understand their full life cycle.
RESUMEN
Los sitios de parada durante la migración son de alta importancia para la conservación, especialmente los sitios en donde las aves playeras se concentran en grandes cantidades para adquirir las reservas de grasa necesarias para el vuelo continuado a los sitios de reproducción. Muchas aves playeras usan solo unos pocos sitios de parada durante la migración en dirección norte hacia las zonas de reproducción en el Ártico, por lo que identificar las localidades importantes, la cronología de la migración y las rutas de vuelo, se convierten en temas prioritarios para la investigación con el fin de informar estrategias de manejo efectivas. Examinamos las rutas y el tiempo hacia las zonas de reproducción en el Ártico de Arenaria interpes, desde una isla en Carolina del Sur en el corazón de la bahía del Atlantico Sur usando sistemas de rastreo de vida silvestre Motus. Los individuos de Arenaria interpes que partieron de Carolina del Sur, no migraron con dirección norte a lo largo de la costa Atlántica, pero migraron por el continente a través de la cuenca de los Grandes Lagos. La mayoría de los individuos de Arenaria interpes no pararon en la cuenca de los Grandes Lagos, haciendo de Carolina del Sur, presumiblemente, su último sitio de parada antes de alcanzar los hábitats del Ártico. Similar a otros estudios sobre la migración de aves playeras, la mayoría de los individuos de Arenaria interpes utilizaron los vientos de cola al momento de partida y la velocidad en tierra estuvo positivamente correlacionada con el soporte por el viento de cola. La planeación futura para la conservación de Arenaria interpes debe considerar la variación en las rutas migratorias y las estrategias de esta especie de ave playera con poblaciones en disminución. Esta investigación también demuestra la utilidad de Motus para rastrear los movimientos de aves playeras pequeñas y el potencial para la expansión estratégica de la red Motus para entender su ciclo de vida completo.
INTRODUCTION
Shorebird numbers in North America are declining, especially the species that nest in the Arctic (Munro 2017, Rosenberg et al. 2019). Three-quarters of North America’s shorebird species experience significant threats during migration, a phase of the annual cycle when mortality rates are much higher than any other time of year (Brown et al. 2001, Klaassen et al. 2014, Piersma et al. 2016, Watts et al. 2019). Spring migratory stopover sites are of high conservation concern because shorebirds concentrate in large numbers to acquire fat reserves to fuel continued flight north to the breeding grounds (Myers et al. 1987, Warnock 2010). Stopover sites may also provide fuel for egg development on nesting grounds (Hobson and Jehl 2010), and food abundance at stopover sites is linked to shorebird stopover mass gain and subsequent annual survival and recruitment of young (Baker et al. 2004, McGowan et al. 2011, Piersma et al. 2016). Many shorebirds use only a few stopover sites during northward migration to Arctic breeding grounds; therefore, identifying important stopover sites, migration chronology, and flight routes are priority research topics to inform effective conservation strategies (Piersma and Baker 2000, Brown et al. 2001).
Delaware Bay, USA, serves as one of the most important stopover and staging areas in the Western Hemisphere for northbound shorebirds (Clark et al. 1993, Niles et al. 2009) representing the expected terminal staging area before a single direct flight to Arctic breeding grounds for several species (Nettleship 2020, Niles and Porter, personal communication). Northbound migratory shorebirds that stage at Delaware Bay rely on the eggs of spawning horseshoe crabs (Limulus polyphemus) to accumulate adequate fuel stores for the final leg of their northbound migration (Robinson et al. 2003, McGowan et al. 2011). This connection between horseshoe crab spawn and shorebird abundance has only recently been recognized in another critically important region for shorebirds, the South Atlantic Bight, which encompasses South Carolina, Georgia, and northeast Florida, USA, and includes three Western Hemisphere shorebird reserve network sites (Department of the Interior 2021, Takahashi et al. 2021, WHSRN 2022). The Ruddy Turnstone (Arenaria intrepes) is abundant in both Delaware Bay and the South Atlantic Bight during northbound migration. In Delaware Bay, Ruddy Turnstones stage in large numbers from late April to early June (peak 20-31 May) before moving north-northwest to Arctic breeding grounds (Clark et al. 1993, Harrington and Flowers 1996). As in Delaware Bay, Ruddy Turnstones are most abundant along the South Atlantic Bight throughout May (Wallover et al. 2015, International Shorebird Survey 2022), suggesting that the South Atlantic Bight may be the final stopover area for some Ruddy Turnstones before migrating to Arctic nesting areas.
Ruddy Turnstone populations have experienced substantial declines since the 1970s, particularly along the U.S. Atlantic Coast (Niles et al. 2009, Andres et al. 2012, Rosenberg et al. 2019). Understanding how and when this species uses key stopover sites and what migratory routes they use upon leaving these sites is necessary to understand their full life cycle and inform conservation decisions. In this first tracking study of Ruddy Turnstones in the South Atlantic Bight, we examined their northbound migration routes and timing using the Motus Wildlife Tracking System, hereafter Motus (Taylor et al. 2017, Birds Canada 2022), an automated telemetry network that enables continental scale tracking studies of species too small for other tracking technologies (e.g., GPS/GSM or satellite tags). Our primary goal was to evaluate the relative use of an Atlantic migratory route through Delaware Bay versus an inland route through the Great Lakes “en route” to Arctic breeding grounds and identify areas of apparent stopovers, particularly along inland routes. Secondarily, because migration routes and departure decisions evolved in part to use beneficial atmospheric conditions, e.g., wind assistance (Åkesson and Hendenström 2000, Conklin and Battley 2011, Duijns et al. 2017, Senner et al. 2018), we explored the association of Ruddy Turnstone routes and ground speeds with prevailing atmospheric conditions.
METHODS
Study area
In late May (21-24 May) of 2018 and 2019, we trapped Ruddy Turnstones at Turtle Island wildlife management area, South Carolina located north of the mouth of the Savannah River in Jasper County, South Carolina (32°3′59″ N, 80°53′33″ W; Fig. 1). Turtle Island is a stopover site in the heart of the South Atlantic Bight with abundant horseshoe crab spawning (Cushman et al. 2019). The 600-ha island is composed primarily of salt marsh with a narrow, forested ridge bordered by a sandy beach, approximately 10 m wide and 1 km long, which is owned by the South Carolina Department of Natural Resources. Thousands of shorebirds fed on horseshoe crab eggs on Turtle Island, with estimates of over 14,000 shorebirds, including 800 Ruddy Turnstones, observed on the 1 km long beach during this study (Sanders 2019, unpublished data, https://ebird.org/checklist/S103555872). The arrival dates of Ruddy Turnstones to this site are not documented, but year-round surveys in South Carolina found Ruddy Turnstone numbers peak in May and decline in June and July, with some birds present throughout the year (Wallover et al. 2015).
Bird capture and tag deployment
Ruddy Turnstones were captured during the day using cannon nets on a falling tide within two hours of high tide when birds were feeding on horseshoe crab eggs. Birds were removed from under the net and moved to keeping cages for immediate processing. All birds were acquiring breeding plumage (definitive prealternative molt; Pyle 2008). Each individual was measured, weighed, and fitted with a uniquely inscribed three-character green leg flag and a USGS numbered metal band. We attached coded very high frequency (VHF) radio-transmitters (166.38 MHz with 4.7 or 8.9 second burst intervals; models NTQB-4-2 and NTQB-4-2S; Lotek Wireless Inc., Newmarket, Ontario, Canada) to each individual by parting the feathers near the bird’s rump above the preen gland and clipping a small area of feathers about the size of the transmitter. This project focused on migration patterns in the month following capture thus the tags were simply glued to the feather stubble with a cyanoacrylate gel adhesive (Loctite super glue gel control, Henkel, USA), leaving the antenna leading straight down the tail. The bird was released once the tag was firmly attached and the glue was completely dry (Warnock and Warnock 1993). Tags weighed 0.90 g or < 1% of individual body mass.
Detection data collection and processing
We processed tag detection data similarly to previously published studies using Motus (Crysler et al. 2016, Duijns et al. 2017, 2019). The primary data associated with each detection are tag identity, detection date and time, receiver station coordinates and the orientation of the receiving antenna (when available), and a measure of signal strength. False detections were eliminated during post-processing by examining all detections with fewer than three consecutive pulses and considering several derived metrics of detection structure related to a tag’s frequency, burst interval, and other signal qualities as well as considering the noise context of the receiving station and the timing and locations of other valid detections of the tag.
Departure dates
To estimate the departure window from coastal South Carolina to northern destinations, we used two methods. First, for birds that were detected by a station away from the coast within South Carolina or North Carolina (n = 13), we considered the date of first detection away from the capture site as the initiation of migration. At the time of this study, only two (2018) or five (2019) receiving stations were operational in South Carolina and none within 75 km of the capture location. The lack of stations near the capture location made it difficult to estimate departure dates precisely for most individuals, although their detection elsewhere in the network (typically the Great Lakes region 2-10 days after tag deployment) was indicative of the range of potential dates they could have departed South Carolina. To make use of all birds detected during northbound migration, we assigned the 34 turnstones with uncertain departure dates a range of potential departure dates. We assigned each day between the tag deployment date and the date of first detection away from the capture site a percentage that reflected the possibility the bird departed on that day. For example, a Ruddy Turnstone with a known departure date made its full contribution (1 individual) to a date, whereas a Ruddy Turnstone with a five-day potential departure window contributed 0.2 “individuals” to each of the days in that date range. Summing these departure date weights for all 47 individuals produced an estimated departure timeline for our full data set while accounting for uncertainty in exact departure dates.
Migratory routes
To compare the relative use of the Atlantic Coastal route through Delaware Bay versus an inland route through the Great Lakes, we examined patterns of tag detections in these watersheds. We defined Delaware Bay as any station within 30 km of the Delaware Bay hydrological unit (HUC 02040204) and the Great Lakes as the Great Lakes and their associated watersheds or the Great Lakes Basin (Great Lakes Restoration Initiative 2010; Fig. 1). To explore how the prevailing wind conditions encountered by migrants during flight may associate with migratory routes through the Great Lakes, we estimated the association between the longitude of first detection in the Great Lakes with the east-west wind component of the detection station near or just prior to detection (see below for more details on wind assistance).
Stopovers
We followed a modified approach of Crysler et al. (2016) to identify potential stopovers. Considering that shorebirds migrating during the day may elect to temporarily rest or refuel i.e., mixing the acts of migration and stopover (Linscott and Senner 2021), we relaxed Crysler et al.’s (2016) detection timeframes and considered stopovers as any detections at a single station spanning > 4 hours (with no intervening detections elsewhere) or spanning > 6 hours among multiple adjacent stations (e.g., stations located within 30 km of each other).
Speed of travel and wind assistance
We estimated migration speeds for Ruddy Turnstones during migratory flights from detection times at receiving stations separated by at least 150 km to exclude local movements and reduce bias from uncertainty in an individual’s precise location during a detection (Duijns et al. 2019) and restricted estimates of migration speed to flights of 18 hours or less. This produced 34 flight trajectories from 29 individuals for analysis of travel speed and wind assistance.
We calculated total trajectory length as the shortest distance connecting all detection receiving stations during the flight between the beginning and ending receiving stations and trajectory displacement length as the shortest distance between the beginning and ending receiving station, using the “trajr” package (McLean and Skowron Volponi 2018) in R (R Core Team 2020). We calculated net ground speed for a flight as the trajectory displacement length divided by the time between the last detection at the beginning receiving station and the first detection at the ending receiving station. We permitted an individual to contribute multiple flights, provided there was no receiver station overlap in the trajectories. For each migratory flight, we also estimated tailwind support at departure using surface wind conditions (i.e., 1000-hPa pressure level) at the measurement time closest to departure. We assumed the “preferred” direction of movement (as required by the calculation of tailwind support) was the bearing that would take the bird to the ending receiving station following a great-circle route. We acquired wind component data for each migratory flight from the NCEP/NCAR reanalysis project (Kalnay et al. 1996) via the “RNCEP” package (Kemp et al. 2012) in R (R Core Team 2020).
All code and data necessary to reproduce the analyses and figures are available via figshare (https://doi.org/10.6084/m9.figshare.22714642).
RESULTS
We tagged 84 Ruddy Turnstones in late May 2018 (24 May) and 2019 (21-22 May). The average mass at transmitter deployment was 150 ± 13 g (median = 152 g). Forty-seven individuals (56%) were detected in the Motus network during northbound migration (i.e., deployment date through 15 June).
Departure dates
The median departure date of 13 individuals with known departure dates from the coast was 27 May (Fig. 2A). Using detection data from all Ruddy Turnstones but incorporating uncertainty in departure date for 34 individuals, the weighted median departure date was 25 May (Fig. 2B).
Migratory routes
Of the 46 Ruddy Turnstones that provided northbound detections from which we could infer migratory route (only one individual was detected along the South Carolina coast after release), none was detected by any stations within Delaware Bay or any of the more than a dozen coastal stations between South Carolina and New Jersey, although most of these stations were operational and detecting other shorebirds at the time of these migrations (www.motus.org; Fig. 1). Most Ruddy Turnstones (43; 93%) were detected by at least one Great Lakes station (Fig. 1). Within the context of active receiver stations in the Great Lakes, most movement occurred through central to eastern Lake Erie or in western Lake Erie and the lower peninsula of Michigan (Fig. 3). The Motus infrastructure in the western Great Lakes (i.e., most of Lakes Michigan and Superior), at the time of this study, was insufficient to evaluate movements through that region. The longitude of first detection in the Great Lakes was associated with the east-west wind component (Pearson’s r = 0.44, p = 0.003; Fig. 4). Stronger winds from the east were associated with initial detections farther west in the Great Lakes and vice versa for western winds. The median days between tag deployment and the first detection in the Great Lakes region was four days (range 1-10 days, n = 43).
Stopovers
Most detection windows in the Great Lakes were short (< 1 day); only four (9%) Ruddy Turnstones were detected for greater than one day, and thus presumed to stopover, in the Great Lakes (Fig. 5A; green highlighted areas in Fig. 3). Three of the four Great Lakes stopovers were in the western Lake Ontario/eastern Lake Erie area; one was in western Lake Erie. Passage through the Great Lakes was concentrated in the last week of May (Fig. 5B). Our estimate of short duration stays in the Great Lakes was also supported by subsequent detections of eight individuals in James Bay or Hudson Bay, for which the median time between last Great Lakes detection and first James/Hudson Bay detection was 15.5 hours (range: 6.9-69.6 hours). Ten Ruddy Turnstones (21%) were detected in James Bay (n = 5) from 25 May - 31 May or Hudson Bay (n = 4) from 27 May - 7 June or both (n = 1; Fig. 1). We could identify only a single stopover in James Bay.
Speed of travel and wind assistance
The median trajectory length of measured migratory flights (n = 34) was 478 km (range: 168-1314 km); the median net displacement (i.e., the orthodrome distance) between beginning and ending receiver stations was 413 km (range: 154-1280 km). Estimated ground speed along 34 flight trajectories representing 29 individuals was unimodal (median: 17.0 m/s; Fig. 6A). There was a small correlation between net ground speed and trajectory length (r = 0.38, p = 0.03). Ground speeds appeared to have some geographical association, whereby the slowest ground speeds were associated with flight trajectories involving one or more receiving stations in the eastern Great Lakes basin (Fig. 6C). Trajectory ground speeds correlated positively but weakly with tailwind support toward the ending receiving station (Pearson’s r = 0.24, p = 0.17; Fig. 6B); however, nearly 75% of migratory trajectories were associated with tailwinds at departure.
DISCUSSION
Motus tracking revealed Ruddy Turnstones using the South Atlantic Bight in late May did not migrate northward along the Atlantic coast but rather over land through the Great Lakes toward Arctic breeding grounds. The short time (median four days) between tag deployment and first detection in the Great Lakes supported the tracking data, suggesting little time for a major stop elsewhere on the Atlantic coast. Our inference of the primary movement routes through the Great Lakes was constrained by the availability of active stations, particularly in the western Great Lakes. However, patterns of detection and the availability of receiving stations suggest that Ruddy Turnstones departing South Carolina in late May typically remain west of Lake Ontario. Ruddy Turnstones leaving Delaware Bay typically pass through the eastern Great Lakes (Duijns et al. 2019, Niles and Porter, unpublished geolocator data), further supporting the finding that Ruddy Turnstones departing South Carolina in late May follow a direct path to and through the Great Lakes.
Most Ruddy Turnstones tracked did not stop in the Great Lakes, thus making South Carolina the last presumed stopover before reaching Arctic habitats. Four Ruddy Turnstones made a brief (< 1 day) stopover in the Great Lakes, and slower ground speeds for some Ruddy Turnstones in the Great Lakes (Fig. 6) suggest Ruddy Turnstones may occasionally punctuate flights with resting or foraging stops. Making brief stopovers is not necessarily a sign of inadequate preparation for migration. Red Knots (Calidris canutus) in Delaware Bay with higher fat stores departed for the breeding grounds later and made shorter stopovers “en route” to still arrive to the breeding grounds earlier than knots in poorer body condition (Duijns et al. 2017). A single Ruddy Turnstone was detected upon entering the Great Lakes Basin and then spent four additional days outside of network detection prior to departing over the lower peninsula of Michigan. This bird may have been using areas without adequate Motus coverage (e.g., western Lake Huron shoreline or Saginaw Bay, possibly Lake St. Clair). An expanded study of Ruddy Turnstones using the South Atlantic Bight in spring, along with expanded and strategic Motus infrastructure throughout the Great Lakes Basin, may more completely reveal the varying ways Ruddy Turnstones and other shorebirds combine use of the South Atlantic Bight with additional stopovers in the Great Lakes to complete their northbound flights to the breeding grounds.
We found evidence to suggest that prevailing wind conditions play a role in the path Ruddy Turnstones take through the Great Lakes (Fig. 6). More intensive flight path modeling (Baldwin et al. 2018) coupled with expanded Motus infrastructure between the South Atlantic Bight and the Great Lakes along an inland route will make it possible to examine the wind conditions encountered along the entire flight path preceding entry into the Great Lakes and may provide additional clarity on the role of wind in migratory paths through this region. Moreover, most Ruddy Turnstones made use of tailwinds at departure from the South Atlantic Bight. Wind is thought to be the most important abiotic driver of global migration routes (Kranstauber et al. 2015), and wind support can reduce energy needed to travel great distances (Duijns et al. 2017). Tailwind support has been the most important factor in explaining migratory distance in raptors (Mellone et al. 2012) and departure date in passerines that predominately make use of favorable winds to migrate (Åkesson and Hedenström 2000). Ruddy Turnstones using the South Atlantic Bight as their final stopover area may deliberately time their departure with conditions that will help them reach the Great Lakes.
This research demonstrates the usefulness of Motus for tracking the migration of smaller shorebirds. The number of stations in South Carolina has grown from the 2-5 stations active during our study to 21 stations in 2022, an increase that is partly attributable to the support generated in response to the novel information gained from this project. The percentage of tagged turnstones we detected (56%) is low compared to the detection rate of Red Knots tagged in the spring in Delaware Bay (84%), however, 42% of those tagged knots were only detected once, mostly at a station near the capture location (Duijns et al. 2017, 2019). Only 29% of the Sanderlings (Calidris alba) tagged on the Gulf coast of the United States were detected in northern latitudes, likely due to minimal Motus coverage through the Midcontinental Flyway at the time of the study (Bianchini et al. 2020). Similarly, the lack of inland Motus stations between South Carolina and the Great Lakes likely affected the percentage of tags detected during our study compared to Motus tracking projects along the Atlantic Flyway (Birds Canada 2022). Research on the effects of nanotags glued to the backs of Piping Plovers (Charadrius melodus) found no differences in survival rates and productivity between birds with or without transmitters (Stantial et al. 2019). Although the effect of nanotags on Ruddy Turnstone’s survival has not been investigated, we expect the 37 Ruddy Turnstones in our study were not detected for a variety of reasons including migration flights through areas with no Motus coverage, nanotag attachment or transmission failure, and possibly death of some birds due to natural causes. For example, one turnstone we tagged was detected on August 11 in James Bay, likely migrating south from the Arctic, but it was never detected moving north. Our study illustrates the potential of Motus to explore and understand the full life cycle of migratory shorebirds. The Motus network has grown significantly in the years since our study, and continued strategic expansion in underrepresented areas of the Western Hemisphere will substantially increase capacity to fill key information needs for priority migratory species.
Our results emphasize that the South Atlantic Bight is a critical final stopover site for some northbound Ruddy Turnstones, presumably where shorebirds can gain adequate energy to fly directly to Arctic habitat. Arctic-breeding shorebirds depend on plentiful and predictable food at a small network of stopover sites that they return to year after year (Mizrahi et al. 2012, Tucker et al. 2022), and the degradation of habitat quality at these sites can impact subsequent migratory performance, reproductive success, and survivorship (Studds et al. 2017). Shorebirds may exhibit high intra- and interannual site fidelity to stopover areas (Buchanan et al. 2012, Neima et al. 2020, Sanders et al. 2021) increasing the importance of protecting critical stopover areas. Identifying a network of key sites for shorebirds is the first step in protecting essential and reliable habitats for shorebirds traveling across a hemisphere (WHSRN 2022). Future conservation planning for shorebirds must include the full network of stopover sites supporting the varied migratory routes and strategies of declining shorebird species.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
We thank the many volunteers who assisted with field work. We appreciate the support of Bird Studies Canada with the Motus Wildlife Tracking System. We are grateful for housing that was provided by Pinckney Island National Wildlife Refuge. Thanks to Rebecca Poulson for support from the Southeast Cooperative Wildlife Disease Study. This research was supported by the South Carolina Department of Natural Resources and federal funding from South Carolina State Wildlife Grants (SC-T-F20AF11231-00). Additional funding was provided by the National Institute of Allergy and Infectious Diseases, NIH, Department of Health and Human Services, under contract HHSN272201400006C, and the U.S. Fish and Wildlife Service, National Wildlife Refuge System, Southeast Inventory and Monitoring Branch.
DATA AVAILABILITY
Nanotag attachment was approved by the U.S. Geological Survey, bird banding laboratory, federal bird banding permit number: 06658, issued to the South Carolina Department of Natural Resources. All code and data necessary to reproduce the analyses and figures are available via Github (https://github.com/adamdsmith/RUTU).
LITERATURE CITED
Åkesson, S., and A. Hedenström. 2000. Wind selectivity of migratory flight departures in birds. Behavioral Ecology and Sociobiology 47:140-144. https://doi.org/10.1007/s002650050004
Andres, B., P. Smith, R. Morrison, C. Gratto-Trevor, S. Brown, and C. Friis. 2012. Population estimates of North American shorebirds. Wader Study Group Bulletin 119(3):178-194. https://www.shorebirdplan.org/wp-content/uploads/2013/03/ShorePopulationAndresEtAl2012.pdf
Baker, A. J., P. M. González, T. Piersma, L. J. Niles, I. de Lima Serrano do Nascimento, P. W. Atkinson, N. A. Clark, C. D. T. Minton, M. K. Peck, and G. Aarts. 2004. Rapid population decline in Red Knots: fitness consequences of decreased refuelling rates and late arrival in Delaware Bay. Proceedings of the Royal Society B: Biological Sciences 271(1541):875-882. https://doi.org/10.1098/rspb.2003.2663
Baldwin, J. W., K. Leap, J. T. Finn, and J. R. Smetzer. 2018. Bayesian state-space models reveal unobserved off-shore nocturnal migration from Motus data. Ecological Modelling 386:38-46. https://doi.org/10.1016/j.ecolmodel.2018.08.006
Bianchini, K., D. J. Newstead, and C. A. Morrissey. 2020. Differences in migration timing along the Midcontinental Flyway in Sanderling (Calidris alba) from three Gulf of Mexico staging areas. Waterbirds 43(3-4):225-356. https://doi.org/10.1675/063.043.0301
Birds Canada. 2022. Motus: fetch and use data from the Motus Wildlife Tracking System. Birds Canada, Port Rowan, Ontario, Canada. https://motus.org/motusRBook/archives/MotusRBook2018-01.pdf
Brown, S., C. Hickey, B. Harrington, and R. E. Gill. 2001. The U.S. shorebird conservation plan. Second edition. Manomet Center for Conservation Sciences, Manomet, Massachusetts, USA.
Buchanan, J. B., J. E. Lyons, L. J. Salzer, R. Carmona, N. Arce, G. J. Wiles, K. Brady, G. E. Hayes, S. M. Desimone, G. Schirato, and W. Michaelis. 2012. Among-year site fidelity of Red Knots during migration in Washington. Journal of Field Ornithology 83(3):282-289. https://doi.org/10.1111/j.1557-9263.2012.00376.x
Clark, K. E., L. J. Niles, and J. Burger. 1993. Abundance and distribution of migrant shorebirds in Delaware Bay. Condor 95(3):694-705. https://doi.org/10.2307/1369612
Conklin, J. R., and P. F. Battley. 2011. Impacts of wind on individual migration schedules of New Zealand Bar-tailed Godwits. Behavioral Ecology 22(4):854-861. https://doi.org/10.1093/beheco/arr054
Crysler, Z. J., R. A. Ronconi, and P. D. Taylor. 2016. Differential fall migratory routes of adult and juvenile Ipswich Sparrows (Passerculus sandwichensis princeps). Movement Ecology 4:3. https://doi.org/10.1186/s40462-016-0067-8
Cushman, E. L., A. E. Fowler, J. D. Robinson, R. Walsh, and T. L. Darden. 2019. Population structure and genetic diversity of spawning Atlantic horseshoe crabs (Limulus polyphemus) along the South Carolina coast. Journal of Shellfish Research 38(3):655-663. https://doi.org/10.2983/035.038.0317
Department of the Interior. 2021. Endangered and threatened wildlife and plants; designation of critical habitat for Rufa Red Knot (Calidris Canutus Rufa). Federal Register, U.S. Fish and Wildlife Service, Washington, D.C., USA. https://www.govinfo.gov/content/pkg/FR-2021-07-15/pdf/2021-14406.pdf
Duijns, S., A. M. Anderson, Y. Aubry, A. Dey, S. A. Flemming, C. M. Francis, C. Friis, C. Gratto-Trevor, D. J. Hamilton, R. Holberton, S. Koch, A. E. McKellar, D. Mizrahi, C. A. Morrissey, S. G. Neima, D. Newstead, L. Niles, E. Nol, J. Paquet, J. Rausch, L. Tudor, Y. Turcotte, and P. A. Smith. 2019. Long-distance migratory shorebirds travel faster towards their breeding grounds, but fly faster post-breeding. Scientific Reports 9:9420. https://doi.org/10.1038/s41598-019-45862-0
Duijns, S., L. J. Niles, A. Dey, Y. Aubry, C. Friis, S. Koch, A. M. Anderson, and P. A. Smith. 2017. Body condition explains migratory performance of a long-distance migrant. Proceedings of the Royal Society B: Biological Sciences 284:20171374. https://doi.org/10.1098/rspb.2017.1374
Great Lakes Restoration Initiative. 2010. Great Lakes Subbasins. U.S. Department of Interior, U.S. Geological Services, Washington, D.C. USA. https://www.sciencebase.gov/catalog/item/530f8a0ee4b0e7e46bd300dd
Harrington, B., and C. Flowers. 1996. The flight of the Red Knot. W. W. Norton, New York, USA.
Hobson, K. A., and J. R. Jehl. 2010. Arctic waders and the capital-income continuum: further tests using isotopic contrasts of egg components. Journal of Avian Biology 41:565-572. https://doi.org/10.1111/j.1600-048X.2010.04980.x
International Shorebird Survey. 2022. International Shorebird Survey Mapping Tool. Manomet, Plymouth, Massachusetts, USA. https://www.manomet.org/iss-map
Kalnay, E., M. Kanamitsu, R. Kistler, W. Collins, D. Deaven, L. Gandin, M. Iredell, S. Saha, G. White, J. Woollen, Y. Zhu, M. Chelliah, W. Ebisuzaki, W. Higgins, J. Janowiak, K. C. Mo, C. Ropelewski, J. Wang, A. Leetmaa, R. Reynolds, R. Jenne, and D. Joseph. 1996. The NCEP/NCAR 40-Year reanalysis project. Bulletin of the American Meteorological Society 77(3):437-472. https://doi.org/10.1175/1520-0477(1996)077<0437:TNYRP>2.0.CO;2
Kemp, M. U., E. Emiel van Loon, J. Shamoun-Baranes, and W. Bouten. 2012. RNCEP: global weather and climate data at your fingertips. Methods in Ecology and Evolution 3(1):65-70. https://doi.org/10.1111/j.2041-210X.2011.00138.x
Klaassen, R. H. G., M. Hake, R. Strandberg, B. J. Koks, C. Trierweiler, K.-M. Exo, F. Bairlein, and T. Alerstam. 2014. When and where does mortality occur in migratory birds? Direct evidence from long-term satellite tracking of raptors. Journal of Animal Ecology 83(1):176-184. https://doi.org/10.1111/1365-2656.12135
Kranstauber, B., R. Weinzierl, M. Wikelski, and K. Safi. 2015. Global aerial flyways allow efficient travelling. Ecology Letters 18(12):1338-1345. https://doi.org/10.1111/ele.12528
Linscott, J. A., and N. R. Senner. 2021. Beyond refueling: investigating the diversity of functions of migratory stopover events. Ornithological Applications 123(1):1-14. https://doi.org/10.1093/ornithapp/duaa074
McGowan, C. P., J. E. Hines, J. D. Nichols, J. E. Lyons, D. R. Smith, K. S. Kalasz, L. J. Niles, A. D. Dey, N. A. Clark, P. W. Atkinson, C. D. T. Minton, and W. Kendall. 2011. Demographic consequences of migratory stopover: linking Red Knot survival to horseshoe crab spawning abundance. Ecosphere 2(6):1-22. https://doi.org/10.1890/ES11-00106.1
McLean, D. J., and M. A. Skowron Volponi. 2018. trajr: an R package for characterisation of animal trajectories. Ethology 124(6):440-448. https://doi.org/10.1111/eth.12739
Mellone, U., R. H. G. Klaassen, C. García-Ripollés, R. Limiñana, P. López-López, D. Pavón, R. Strandberg, V. Urios, M. Vardakis, and T. Alerstam. 2012. Interspecific comparison of the performance of soaring migrants in relation to morphology, meteorological conditions and migration strategies. PLoS ONE 7(7):e39833. https://doi.org/10.1371/journal.pone.0039833
Mizrahi, D. S., K. A. Peters, and P. A. Hodgetts. 2012. Energetic condition of Semipalmated and Least Sandpipers during northbound migration staging periods in Delaware Bay. Waterbirds 35(1):135-145. https://doi.org/10.1675/063.035.0113
Munro, M. 2017. What’s killing the world’s shorebirds? Nature 541:16-20. https://doi.org/10.1038/541016a
Myers, J. P., R. I. G. Morrison, P. de Tarso Z. Antas, B. A. Harrington, T. E. Lovejoy, M. Sallaberry, S. Senner, and A. Tarak. 1987. Conservation strategy for migratory species. American Scientist 75(1):19-26. https://doi.org/10.1179/019713687806027861
Neima, S. G., D. J. Hamilton, C. L. Gratto-Trevor, and J. Paquet. 2020. Intra- and interannual regional fidelity of Semipalmated Sandpipers (Calidris pusilla) during migratory stopover in the upper Bay of Fundy, Canada. Avian Conservation and Ecology 15(1):14. https://doi.org/10.5751/ACE-01561-150114
Nettleship, D. N. 2020. Ruddy Turnstone (Arenaria interpres). Version 1.0. In S. M. Billerman, editor. Birds of the world. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.rudtur.01
Niles, L. J., J. Bart, H. P. Sitters, A. D. Dey, K. E. Clark, P. W. Atkinson, A. J. Baker, K. A. Bennett, K. S. Kalasz, N. A. Clark, J. Clark, S. Gillings, A. S. Gates, P. M. GonzáLez, D. E. Hernandez, C. D. T. Minton, R. I. G. Morrison, R. R. Porter, R. K. Ross, and C. R. Veitch. 2009. Effects of horseshoe crab harvest in Delaware Bay on Red Knots: are harvest restrictions working? BioScience 59(2):153-164. https://bioone.org/journals/bioscience/volume-59/issue-2/bio.2009.59.2.8/Effects-of-Horseshoe-Crab-Harvest-in-Delaware-Bay-on-Red/10.1525/bio.2009.59.2.8.short
Piersma, T., and A. J. Baker. 2000. Life history characteristics and the conservation of migratory shorebirds. Pages 105-124 in L. M. Gosling and W. J. Sutherland, editors. Behaviour and conservation. Cambridge University Press, Cambridge, UK.
Piersma, T., T. Lok, Y. Chen, C. J. Hassell, H.-Y. Yang, A. Boyle, M. Slaymaker, Y.-C. Chan, D. S. Melville, Z.-W. Zhang, and Z. Ma. 2016. Simultaneous declines in summer survival of three shorebird species signals a flyway at risk. Journal of Applied Ecology 53(2):479-490. https://doi.org/10.1111/1365-2664.12582
Pyle, P. 2008. Identification guide to North American birds, part II. Slate Creek, Point Reyes Station, California, USA.
R Core Team. 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. r-project.org
Robinson, R. A., P. W. Atkinson, and N. A. Clark. 2003. Arrival and weight gain of Red Knots (Calidris canutus), Ruddy Turnstone (Arenaria interpres), and Sanderling (Calidris Alba) staging in Delaware Bay in spring. British Trust for Ornithology, Thetford, UK.
Rosenberg, K. V., A. M. Dokter, P. J. Blancher, J. R. Sauer, A. C. Smith, P. A. Smith, J. C. Stanton, A. Panjabi, L. Helft, M. Parr, and P. P. Marra. 2019. Decline of the North American avifauna. Science 366(6461):120-124. https://doi.org/10.1126/science.aaw1313
Sanders, F. J., M. C. Handmaker, A. S. Johnson, and N. R. Senner. 2021. Nocturnal roost on South Carolina coast supports nearly half of Atlantic coast population of Hudsonian Whimbrel (Numenius hudsonicus) during northward migration. Wader Study Group Bulletin 128(2):117-124. https://doi.org/10.18194/ws.00228
Senner, N. R., M. Stager, M. A. Verhoeven, Z. A. Cheviron, T. Piersma, and W. Bouten. 2018. High-altitude shorebird migration in the absence of topographical barriers: avoiding high air temperatures and searching for profitable winds. Proceedings of the Royal Society B: Biological Sciences 285:20180569. https://doi.org/10.1098/rspb.2018.0569
Stantial, M., J. Cohen, P. Loring, and P. Paton. 2019. Radio transmitters did not affect apparent survival rates of adult Piping Plovers (Charadrius melodus). Waterbirds 42(2):205-209. https://doi.org/10.1675/063.042.0207
Studds, C. E., B. E. Kendall, N. J. Murray, H. B. Wilson, D. I. Rogers, R. S. Clemens, K. Gosbell, C. J. Hassell, R. Jessop, D. S. Melville, D. A. Milton, C. D. T. Minton, H. P. Possingham, A. C. Riegen, P. Straw, E. J. Woehler, and R. A. Fuller. 2017. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nature Communications 8:14895. https://doi.org/10.1038/ncomms14895
Takahashi, F., F. J. Sanders, and P. G. R. Jodice. 2021. Spatial and temporal overlap between foraging shorebirds and spawning horseshoe crabs (Limulus polyphemus) in the Cape Romain-Santee Delta Region of the U.S. Atlantic coast. Wilson Journal of Ornithology 133(1):58-72. https://doi.org/10.1676/21-00009
Taylor, P. D., T. L. Crewe, S. A. Mackenzie, D. Lepage, Y. Aubry, Z. Crysler, G. Finney, C. M. Francis, C. G. Guglielmo, D. J. Hamilton, R. L. Holberton, P. H. Loring, G. W. Mitchell, D. R. Norris, J. Paquet, R. A. Ronconi, J. R. Smetzer, P. A. Smith, L. J. Welch, and B. K. Woodworth. 2017. The Motus Wildlife Tracking System: a collaborative research network to enhance the understanding of wildlife movement. Avian Conservation and Ecology 12(1):8. https://doi.org/10.5751/ACE-00953-120108
Tucker, A. M., C. P. McGowan, J. E. Lyons, A. DeRose-Wilson, and N. A. Clark. 2022. Species-specific demographic and behavioral responses to food availability during migratory stopover. Population Ecology 64(1):19-34. https://doi.org/10.1002/1438-390X.12094
Wallover, N. J., M.-C. Martin, and F. J. Sanders. 2015. Monthly abundance and seasonal spatial distribution of shorebirds in Cape Romain National Wildlife Refuge, South Carolina. Chat 79(2):61-79. https://whsrn.org/wp-content/uploads/2019/03/shorebirds-in-cape-romain.pdf
Warnock, N. 2010. Stopping vs. staging: the difference between a hop and a jump. Journal of Avian Biology 41:621-626. https://doi.org/10.1111/j.1600-048X.2010.05155.x
Warnock, N., and S. Warnock. 1993. Attachment of radio-transmitters to sandpipers: review and methods. Wader Study Group Bulletin 70:28-30. https://sora.unm.edu/sites/default/files/journals/iwsgb/n070/p00028-p00030.pdf
Watts, B. D., F. M. Smith, D. J. Hamilton, T. Keyes, J. Paquet, L. Pirie-Dominix, J. Rausch, B. Truitt, B. Winn, and P. Woodard. 2019. Seasonal variation in mortality rates for Whimbrels (Numenius phaeopus) using the Western Atlantic Flyway. Condor 121(1):1-13. https://doi.org/10.1093/condor/duy001
Western Hemisphere Shorebird Reserve Network (WHSRN). 2022. About WHSRN. WHSRN, Manomet, Massachusetts, USA. https://whsrn.org/about-whsrn/.
Fig. 1
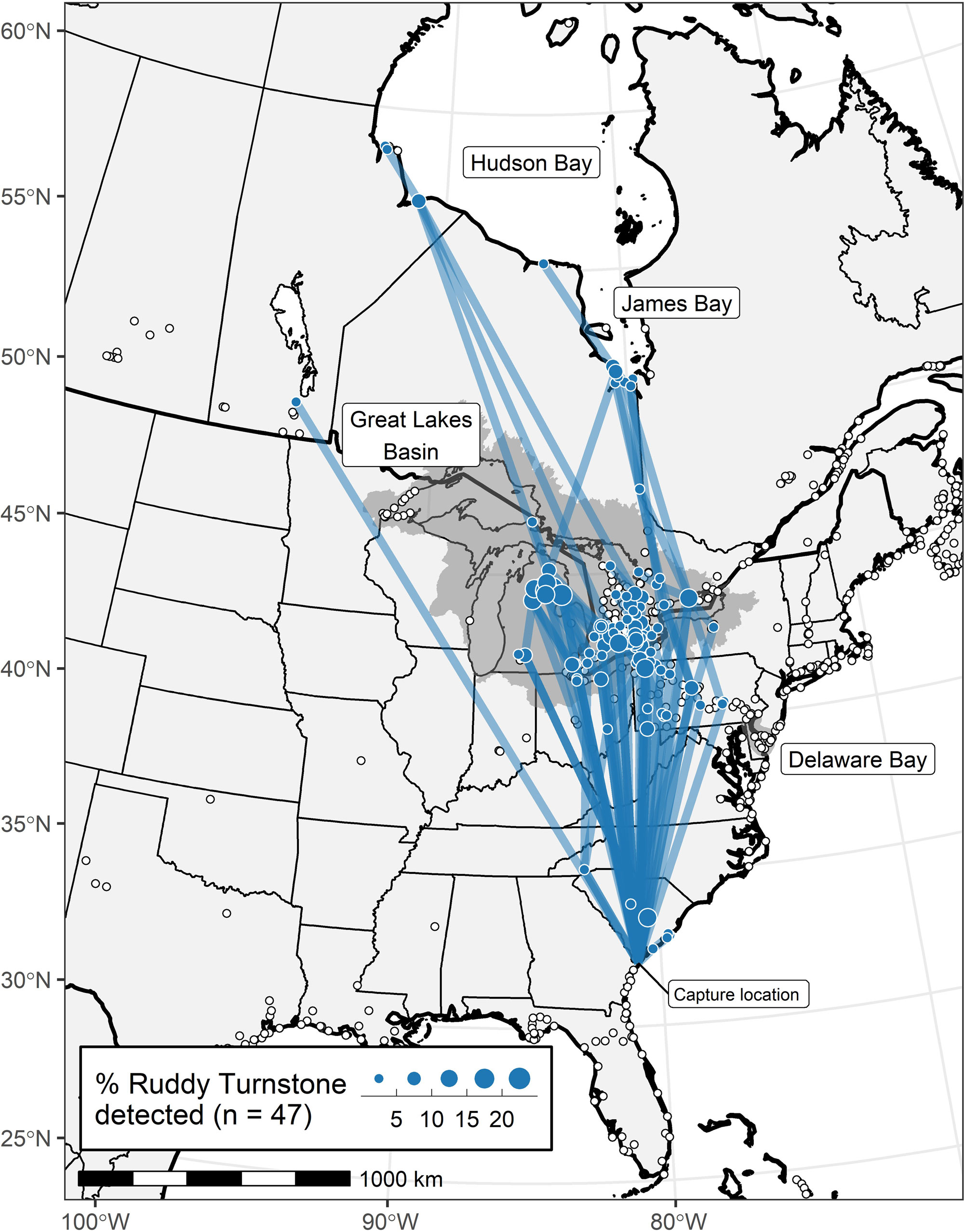
Fig. 1. Locations of active Motus receiver stations (white dots with black outlines) and the northern migratory trajectories of Ruddy Turnstones (Arenaria interpres) tagged in South Carolina in May 2018 and 2019. The lines connect detections of individuals between receiving stations or between the tagging location and a receiver station. These tracks represent simplified flight trajectories not necessarily actual flight paths. Circle diameters reflect the number of individual birds detected by a given receiving station. The Great Lakes Basin and the Delaware Bay hydrological unit are shaded in gray.
Fig. 2
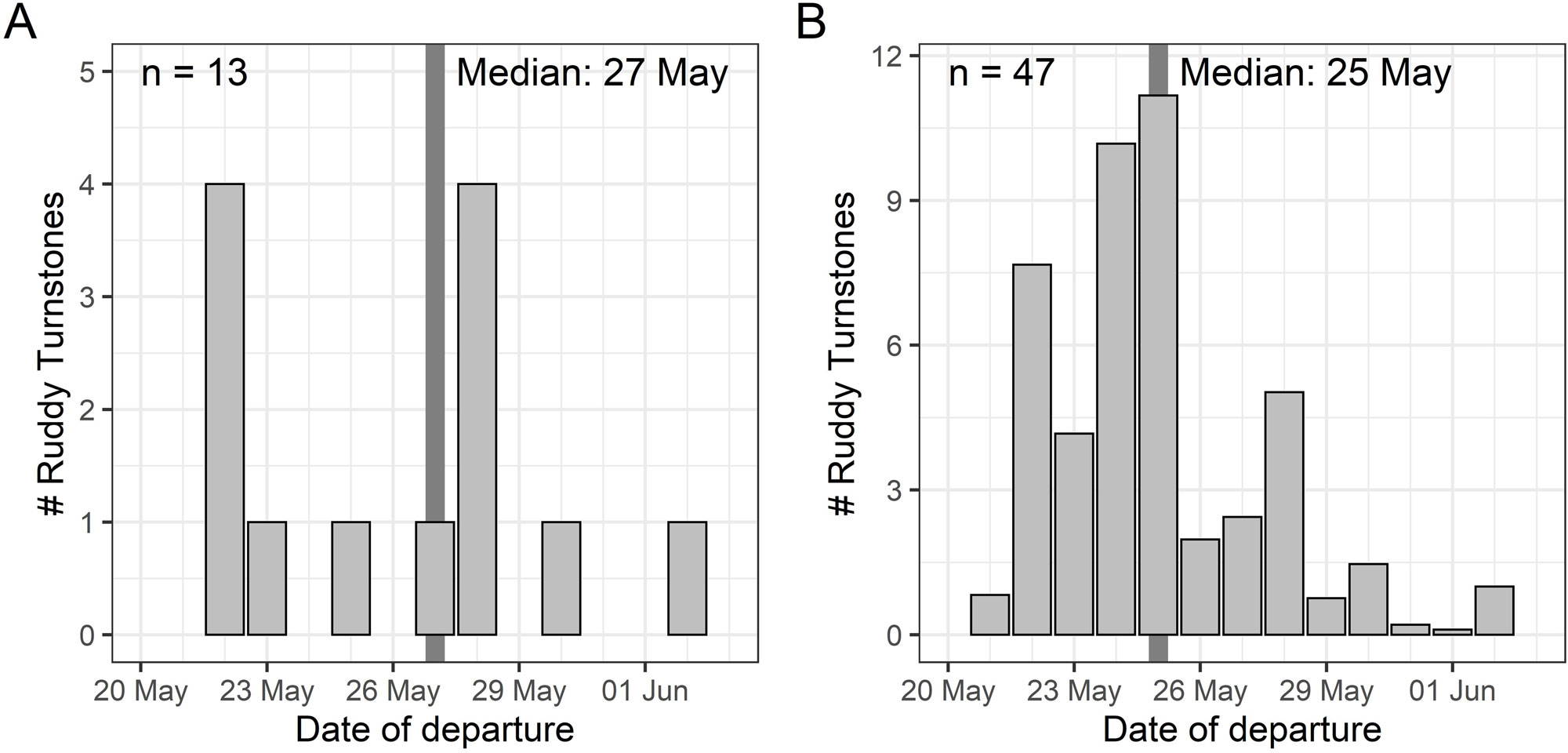
Fig. 2. The departure date (A) of 13 Ruddy Turnstones (Arenaria interpres) migrating north from coastal South Carolina determined by detection at inland stations on the night of departure. The lack of Motus stations near capture locations made it difficult to get precise departure dates for most Ruddy Turnstones, so we incorporated uncertainty in the departure date range for 34 additional individuals to estimate the weighted number of all Ruddy Turnstone departures (B) as described in the text.
Fig. 3
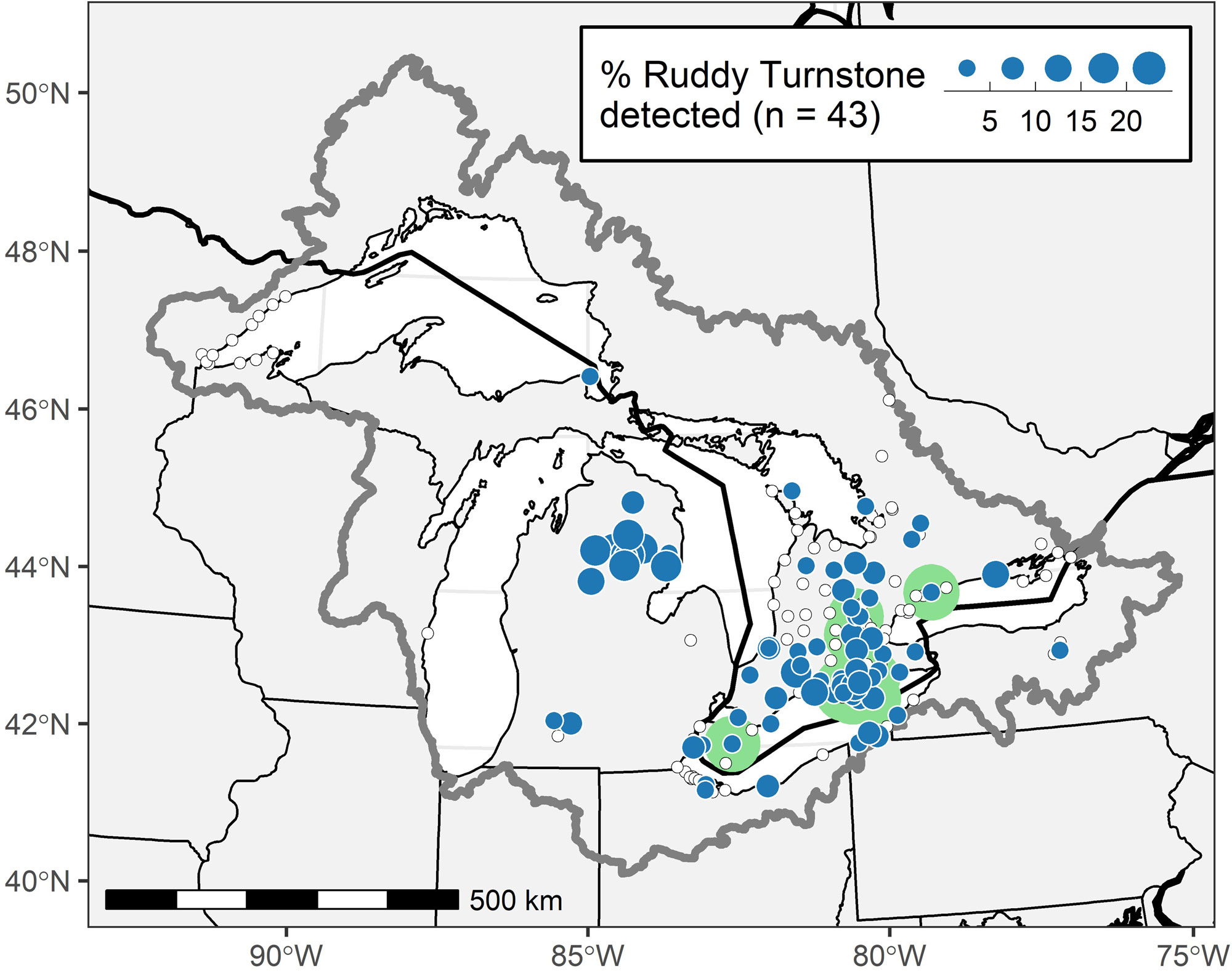
Fig. 3. Locations of active Motus receiver stations (white dots with black outlines), and receiver locations where Ruddy Turnstones (Arenaria interpres) were detected in the Great Lakes Basin during northern migration from May 22 to June 3 (blue circles). Circle diameters indicate the number of individuals detected by each station. Individuals can be detected by multiple stations. Green circles identify the general areas where four turnstones stopped over in the Great Lakes Basin for longer than one day.
Fig. 4
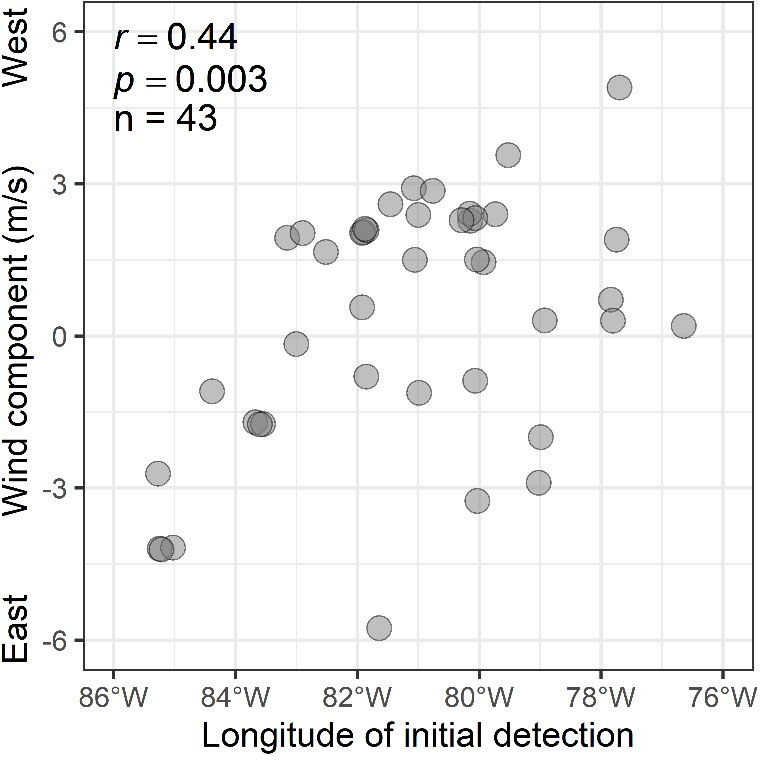
Fig. 4. The association (Pearson correlation) of the longitude of first detection in the Great Lakes Basin and the east-west wind component at the time of detection for Ruddy Turnstones (Arenaria interpres) tagged in South Carolina in May 2018 and 2019.
Fig. 5
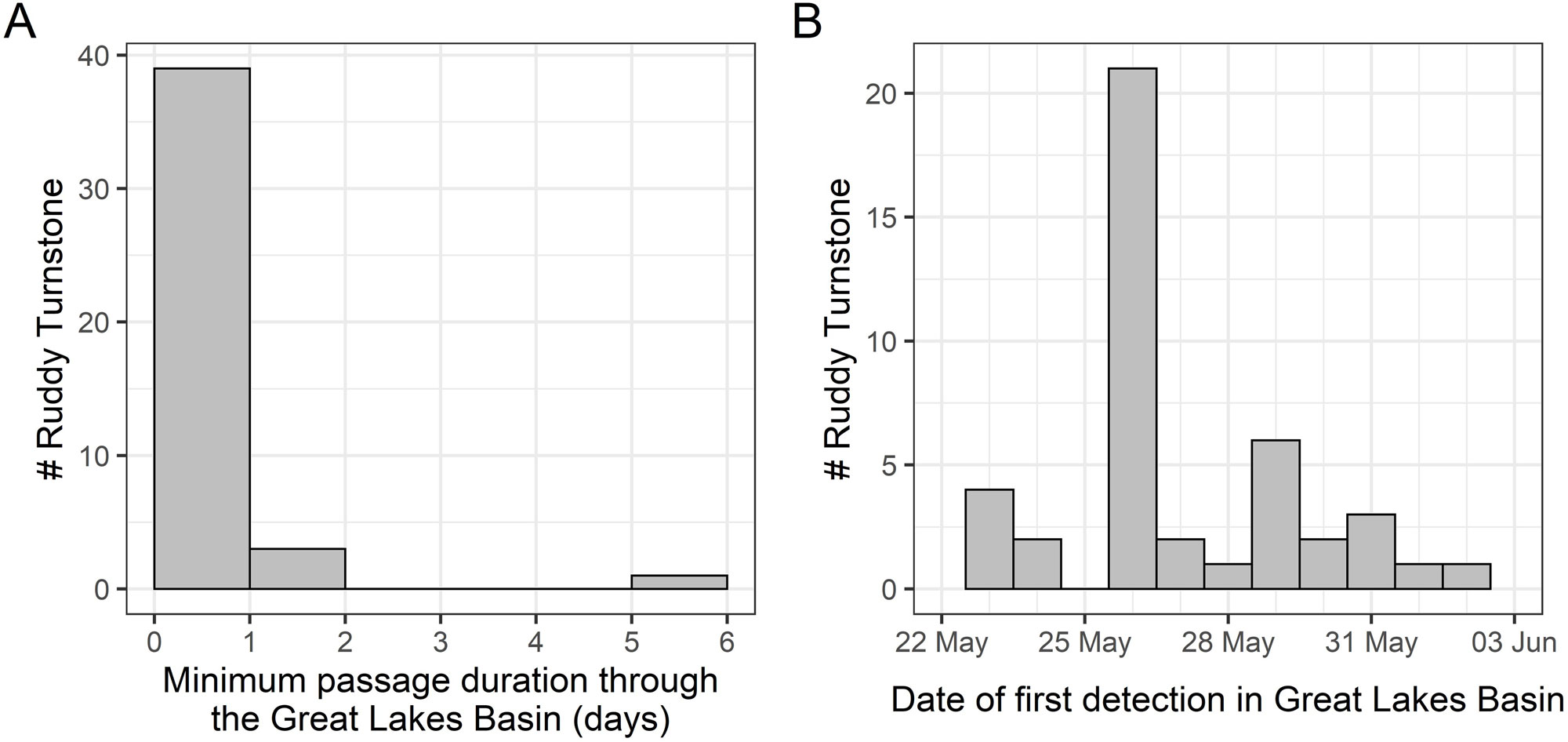
Fig. 5. Ruddy Turnstone (Arenaria interpres) minimum passage time (A) and dates of first detection (B) in the Great Lakes Basin during their northward migration from South Carolina.
Fig. 6
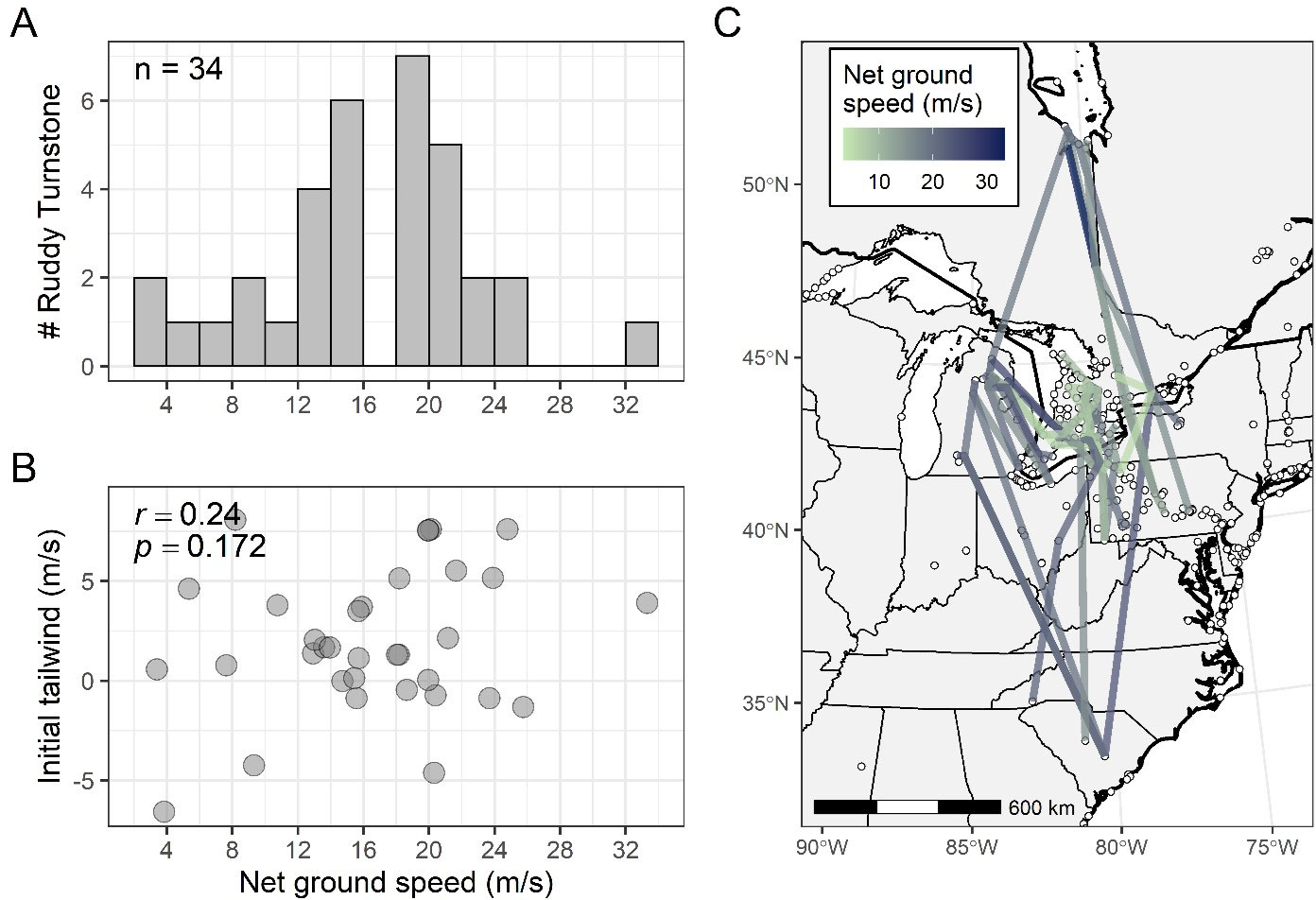
Fig. 6. Estimated ground speed along 34 flight trajectories of 29 Ruddy Turnstones (Arenaria interpres). Migration speed was calculated by restricting estimates to detection times at receiving stations separated by at least 150 km and continuous flights not exceeding 18 hours in duration. Estimated ground speed along 34 flight trajectories was unimodal (median = 17.0 m/s) (A), and the Pearson correlation of trajectory net ground speeds and tailwind support was positive but weak (B). Flight trajectory net ground speeds (C) suggest some Ruddy Turnstones use slower ground speeds as they pass through the eastern Great Lakes Basin relative to flights into, out of, and through the western half of the basin.