







The following is the established format for referencing this article:
Cantu de Leija, A., R. E. Mirzadi, J. M. Randall, M. D. Portmann, E. J. Mueller, and D. E. Gawlik. 2023. A meta-analysis of disturbance caused by drones on nesting birds. Journal of Field Ornithology 94(2):3.ABSTRACT
The use of drones for monitoring nesting birds is rapidly increasing given their affordability and efficiency in bird detection and quantification across habitats. Reports of disturbance caused by drones on different bird species have been mixed, with no consensus on the degree to which different factors affect disturbance responses. Given the lack of systematic assessments of disturbance from drones on nesting birds, we conducted a formal meta-analysis to quantify the degree of disturbance caused by multi-rotor drones on nesting birds, with a particular focus on the effects of altitude of flights and species nesting traits. Seventeen studies met our criteria for inclusion in the analysis, from which we extracted 31 effect sizes in the form of log-odds ratio. Drones showed a small disturbance effect (-1.54; 95% CI: -2.83, -0.26) on nesting birds overall, but heterogeneity was large. Drone flights > 50 m showed no evidence of disturbance on nesting birds. Conversely, flights at lower altitudes (≤ 50 m) showed stronger evidence of disturbance effects, with the largest odds of disturbance observed on ground solitary and non-ground solitary nesters. Only ground colonial nesters showed no evidence of disturbance regardless of the drone altitude. We conclude that the use of drones can be an efficient and safe means of surveying nesting birds if altitude and nesting traits are considered in survey protocols.
RESUMEN
El uso de drones para monitorear las aves que anidan está aumentando rápidamente dada su asequibilidad y eficiencia en la detección y cuantificación de aves en todos los hábitats. Los informes de perturbaciones causadas por drones en diferentes especies de aves han sido mixtos, sin consenso sobre el grado en que los diferentes factores afectan las respuestas de perturbación. Dada la falta de evaluaciones sistemáticas de la perturbación de los drones en las aves que anidan, realizamos un metanálisis formal para cuantificar el grado de perturbación causada por drones multirotor en las aves que anidan, haciendo foco en particular en los efectos de la altitud de los vuelos y los rasgos de anidación de las especies. Diecisiete estudios cumplieron los criterios de inclusión en el análisis, de los cuales se extrajeron 31 tamaños del efecto en forma de relación logarítmica de probabilidades. Los drones mostraron un pequeño efecto de perturbación (-1,54; IC del 95%: -2,83, -0,26) en las aves que anidan en general, pero la heterogeneidad fue grande. Los vuelos de drones > 50 m no mostraron evidencia de perturbación en las aves que anidan. Por el contrario, los vuelos a altitudes más bajas (≤ 50 m) mostraron una mayor evidencia de efectos de perturbación, con las mayores probabilidades de perturbación observadas en anidadores solitarios y no terrestres. Solo los anidadores coloniales terrestres no mostraron evidencia de perturbación, independientemente de la altitud del dron. Concluimos que el uso de drones puede ser un medio eficiente y seguro para inspeccionar las aves que anidan si la altitud y los rasgos de anidación se consideran en los protocolos de monitoreo.
INTRODUCTION
Many bird surveys are focused on measuring reproductive parameters such as nest abundance, nest survival, and brood size at fledging because this information helps track fluctuations in populations and environmental conditions to guide conservation and restoration of habitats (Powell and Powell 1986, Götmark 1992). Thus, significant efforts and resources have been and will continue to be invested in monitoring nesting birds globally. But, to accurately assess fluctuations in reproductive performance, monitoring methods must be accurate, feasible, and consistent (Martin and Geupel 1993, Sutherland et al. 2004, Afán et al. 2018).
Often, bird species nest in areas that are inaccessible by land, are spaced large distances apart and may nest sympatrically with other bird species (e.g., Frederick et al. 1996). Traditionally, nesting birds have been surveyed using crewed aerial and ground-based monitoring methods. Ground-based surveys are often used to obtain productivity measures and estimate nest abundance of populations that are difficult to observe aerially, whereas crewed aerial survey methods are often necessary to monitor the size, composition, and status of nesting birds (Tremblay and Ellison 1979, Gibbs et al. 1988, Frederick et al. 1996). Ground monitoring techniques are expensive and may cause disturbance resulting in negative effects on reproduction (Tremblay and Ellison 1979, Frederick et al. 1996). Crewed aerial surveys are expensive, can have significant observer and detection biases, and often cannot accurately measure productivity (Frederick et al. 1996, Rodgers et al. 2005). Additionally, crewed aerial surveys are the leading cause of job-related mortality for wildlife biologists (Sasse 2003).
Recently, interest has increased in the use of small uncrewed aircraft systems (sUAS, also known as unmanned aircraft systems, UAVs), hereafter drones, for environmental and wildlife monitoring applications (Linchant et al. 2015, Pimm et al. 2015, Lyons et al. 2019). Drones have proven effective in several wildlife monitoring applications, most notably in monitoring avian species. For example, drone count surveys detected > 93% of ground survey counts of Common Terns (Sterna hirundo), but remarkably, ground counts required 12 surveyors and 4 hours to complete whereas the drone survey took 2 surveyors and about 90 minutes total (Chabot et al. 2015). Drones outperformed ground count surveys of breeding Eurasian Oystercatchers (Heamatopus ostralegus) in significantly less time and resulting in an overall 88% cost reduction compared to ground surveys (Valle and Scarton 2020). Drones have also achieved higher detection rates of birds than traditional surveys for species of waterfowl (McEvoy et al. 2016) and for nesting Glossy Ibis (Plegadis falcinellus; Afán et al. 2018). Moreover, drones allowed the modeling of nest survival in Western Grebes (Aechmophorus occidentalis), which had not been possible using traditional methods given the difficulty to observe nests and the high risk of nest or colony abandonment due to observer disturbance (Lachman et al. 2020).
However, studies differ on the degree to which drones caused disturbance to nesting birds (Barr et al. 2020, Valle and Scarton 2020, Gallego and Sarasola 2021), with no consensus on the traits that affect the magnitude of disturbance. Nesting birds often respond to disturbances by increasing vigilant behavior, agonistic behavior, standing at or walking away from the nest, and escape behavior (e.g., flushing; Weimerskirch et al. 2018, Barr et al. 2020), respectively. When these behavioral responses are severe, they may lead to the abandonment of young and increased energy expenditure, resulting in breeding failures (Borrelle and Fletcher 2017). Additionally, physiological responses to disturbance, including increased heart rate and hormonal fluctuations, have been previously linked to elevated metabolic rates that may cause declines in condition (Borrelle and Fletcher 2017). These adverse outcomes are ultimately influenced by the extent and severity of the disturbance, as well as the sensitivity of the nesting bird species.
Drones are often cited as causing less disturbance to nesting birds than traditional survey methods, with recent studies suggesting that the magnitude of disturbance associated with drone surveys may vary by species, life-history traits, the type of drone used (quadcopter vs. fixed-wing), and the altitude at which the drone is flown (Chabot et al. 2015, Borrelle and Fletcher 2017, Barr et al. 2020). For example, several studies have recorded low or no disturbance, either behaviorally or reproductively, on wading birds (Barr et al. 2020), seabirds (Brisson-Curadeau et al. 2017, Fudala and Bialik 2022), and Chaco Eagles (Buteogallus coronatus; Gallego and Sarasola 2021). In contrast, drone flights have been shown to significantly exacerbate disturbance behavior for species such as the Common Redshank (Tringa totanus; Valle and Scarton 2020) and Osprey (Pandion haliaetus; Junda et al. 2016). Additionally, the altitude of drone surveys may influence behavioral responses by nesting birds (Barr et al. 2020). For example, swifts (Mesquita et al. 2021) and Franklin’s Gulls (Leucophaeus pipixcan; McKellar et al. 2021) show stronger disturbance when the drone is flying below 50 m from the nest compared to > 50 m. In contrast, species like Siberian Cranes (Leucogeranus leucogeranus; Wen et al. 2021) and Western Grebes (Lachman et al. 2020) showed low or no disturbance regardless of altitude. However, no systematic effort has been made to quantify the degree to which drones cause disturbance in nesting birds and how surveying practices (e.g., altitude of flights) and species life-history traits may influence the response by birds.
In this study, we conducted a meta-analysis to quantify the degree to which drones cause disturbance on nesting birds. Additionally, we examined the degree to which the magnitude of disturbance varied relative to the altitude at which drones are flown and by species nesting traits (i.e., nest location and companionship behavior). We hypothesized that drones would have an overall small disturbance effect on nesting birds; however, we expected large heterogeneity across studies. Furthermore, we predicted that the altitude of drone flights will best account for heterogeneity across studies regardless of species nesting traits. Given the increased use of drones for surveying nesting birds, we aim to provide information that will help researchers in their study design process and apply surveying practices that will reduce disturbance on nesting birds.
METHODS
Study selection criteria
Following the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) Statement (Fig. 1; Page et al. 2021) to identify a pool of relevant studies, we began with database searches through Google Scholar (https://scholar.google.com/) and Web of Science (https://www.webofscience.com) using combinations of the following keywords: (UAV OR sUAS OR drone) AND (birds OR wading bird OR shorebird OR raptor OR waterfowl OR passerine OR upland game bird OR crane OR vulture OR waterbird OR seabird) AND (disturbance OR behavioral response) AND (nesting OR breeding).
Disturbance is generally defined as any behavioral response that includes standing up and moving off the nest, alert behavior, or increased vigilance, and flushing from nests, returning or not (Klein 1993). However, this definition can be rather subjective and can be interpreted differently for different species or research goals (e.g., Weimerskirch et al. 2018). In this study, disturbance was considered to occur if nesting birds moved off the nest, returning or not, as a response to drone presence, including, but not limited to flushing, swimming away, or escape behavior. Leaving the nest in response to drones is considered the strongest reaction because it involves leaving nests or chicks unprotected (Rümmler et al. 2018). This definition allows disturbance responses to be compared equally across studies regardless of how each individual study defined disturbance and accounts for studies that do not define what is considered as disturbance.
Once all the potential studies were identified through our database search, we began the screening process by first extracting all the duplicates and non-peer reviewed articles. We then removed those studies that did not survey actively nesting or breeding bird species. We further restricted our studies to those using multi-rotor drones because fixed-wing drones have been found to increase disturbance in some species due to their similarity to predators (e.g., raptors; McEvoy et al. 2016), thereby holding constant the effects of drone type.
We removed studies that did not provide a measure of bird disturbance and/or the sample size of birds evaluated (direct count, percentage, or proportion), studies that did not explicitly measure and provide the altitudes at which the drone surveys were conducted, as well as those that did not provide the altitudes at which a disturbance response was elicited. For measurements not explicitly stated in the text but presented in figures, we used values (or closest values) from figures. We contacted authors if measurements were unclear or referenced in the study but not presented and publicly available. In the final step of the study selection process, we compiled the relevant data from each study into a comprehensive table for statistical analyses (Table 1).
After the study selection was complete, we incorporated the quantitative measurements necessary to extract the effect sizes. This constituted the total sample size of the birds observed and a quantification of the number of birds disturbed during the survey trials, reported as direct counts, proportions, averages, or percentages. Each disturbance metric was converted into an effect size thereby providing standardization across studies.
Moderator variables
In meta-analyses, moderator variables are covariates that act at the study level (i.e., systematic difference among studies) used to assess the impact of covariates and to predict the effect size in studies with specific characteristics (Borenstein et al. 2009). We defined the altitude of the drone and bird species nesting trait as two moderator variables that may explain heterogeneity in the size of the disturbance effect by nesting birds to drones. The altitude of a drone from the nest during observational surveys (considered here as a measurement of the distance between the nest and the drone, vertical or diagonal) has been previously shown to have a large effect on the level of disturbance exhibited by nesting birds. Barr et al. (2020) found that disturbance increased significantly for nesting colonies of waterbirds when drones were flown at 46 m, as opposed to altitudes of 61 m, 91 m, and 122 m. Rümmler et al. (2016) observed increased disturbance in penguins during take-off at 50 m, with vigilance remaining elevated at altitudes between 20 m to 50 m. Moreover, related studies have recommended conducting drone surveys at distances greater than 50 m to avoid moderate to high levels of disturbance in nesting birds (Mesquita et al. 2021). Given the repeated reference to an approximate altitude of 50 m as a threshold or boundary for an effect, and to provide adequate sample sizes to test for an effect of altitude, we classified the altitude of each effect size as either ≤ 50 m or > 50 m.
The nesting traits of avian taxa have also been reported to result in differential sensitivity to disturbances (Blackmer et al. 2004, Carey 2009). Colonial nesting species are often considered to be at higher risk of certain disturbances because the high density and proximity of nests may result in disturbance to many birds simultaneously, and possibly favoring predation by opportunistic nest predators (Götmark 1992, Carney and Sydeman 1999). Moreover, nest vulnerability has been shown to vary depending on nest location (Burger 1981). For example, nesting on the ground may influence the effects of researcher disturbance because ground nest predators are mainly mammals, and ground-nesting species may show an increased reaction to ground intrusions (Richardson et al. 2009). Ground-nesting birds such as Brown Pelicans (Pelecanus occidentalis) have been shown to be particularly vulnerable to disturbances (Blackmer et al. 2004). Therefore, we tested for differences in the size of the disturbance effect caused by drones based on coloniality and nesting strata. Bird species or groups of species were designated to one of the following categories: ground colonial, ground solitary, non-ground colonial, and non-ground solitary. Studies reporting groups of multiple species (e.g., wading birds, swifts) with varying nesting traits were split and the effect size was calculated for each.
Effect-size extraction and meta-analysis
We used the odds ratio statistic to calculate the effect size for each species or group of species by the altitude at which the drone was flown during observational surveys (≤ 50 m or > 50 m) and by nesting traits. The odds ratio (i.e., the probability of one event occurring rather than an alternative event) allowed us to quantitatively compare the relationship between birds that were disturbed by drones and those that were not (i.e., the size of the disturbance effect; Borenstein et al. 2009, Viechtbauer 2010). The odds ratio was transformed to the natural log scale to standardize effect sizes among studies (Borenstein et al. 2009). We measured 95% confidence intervals (CIs) for each effect size to depict the precision with which the effect size was calculated in each study (Borenstein et al. 2009). We considered there to be strong evidence of a disturbance effect when the upper and lower CIs were both positive log-odds values, whereas CIs that were both negative indicated strong evidence of no disturbance. Effect sizes with both positive and negative CIs suggested that there was weak evidence of a disturbance or no disturbance because CIs overlapping zero may be due to a small sample size of observed birds or to a small ratio of one event being observed over the other (e.g., equal or near equal number of disturbed and non-disturbed birds).
Effect sizes were used to construct three models to assess the overall degree of disturbance (model 1) and how much heterogeneity could be explained by altitude (model 2) and species nesting traits (model 3). First, to evaluate the overall degree to which drones caused a disturbance to nesting birds, we constructed a random-effects meta-analysis model using the maximum-likelihood estimate of heterogeneity (Borenstein et al. 2009, Gurevitch et al. 2018) based on the log-odds effect sizes of birds that were reportedly disturbed by drones compared to the total number of birds observed. Random-effects meta-analysis assumes that the true effect sizes are not identical across studies, and the magnitude of the effects can vary due to “random” factors and characteristics in studies (Borenstein et al. 2009). Then, to specifically assess if the altitude of the drones (model 2; 2 levels) or the species’ nesting traits (model 3; 4 levels) could account for heterogeneity in the size of the disturbance effect, we constructed a mixed-effects meta-regression model including each variable as a moderator (López-López et al. 2014). This approach allowed the modeling of heterogeneity among studies by including these moderators as “fixed” effects in addition to the random (between-studies variance) effect (hence “mixed-effects meta-regression;” Borenstein et al. 2009, López-López et al. 2014). All models included the test for heterogeneity represented by Cochran’s Q and P < 0.05 indicating large heterogeneity, calculated as the weighted sum of squared differences between each study’s effect and the pooled effect across studies (Borenstein et al. 2009). Additionally, effect sizes were assessed and visualized through boxplots partitioned by altitude and species’ nesting traits to represent the pooled distribution of individual study’s effect sizes for each moderator variable. Boxplots indicate the median, upper (75%), and lower (25%) interquartile range, max, and min values (whiskers), and outliers (dots; if any).
Quantile-quantile normal plots (Q-Q plots) were used to assess the goodness of fits on all three models, which show the theoretical quantiles of a normal distribution against the observed quantiles of the standardized residuals that should approximate a straight vertical line (Viechtbauer 2010). We assessed publication bias (i.e., if included studies are a biased sample of all relevant studies) visually by the asymmetry of effect sizes against their corresponding standard errors in funnel plots (Gurevitch et al. 2018) and statistically by Egger’s test (Egger et al. 1997) with P < 0.05 indicating the presence of publication bias. If publication bias is detected, a “trim and fill” method would be applied as a non-parametric approach to estimate missing studies from the meta-analysis (Viechtbauer 2010). All statistical analysis and plotting were conducted in program R (R Core Team 2021) using the “metafor” (Viechtbauer 2010) and “tidyverse” (Wickham et al. 2019) packages.
RESULTS
From a total of 79 studies originally identified, 17 studies evaluating nesting bird disturbance caused by drones met our selection criteria (Fig. 1). From these 17 studies, we extracted 31 effect sizes, including 22 in which the drone was flown at or below 50 m and 9 flown above 50 m, and representing 10 ground colonial, 4 ground solitary, 10 non-ground solitary, and 7 non-ground colonial nesters across 19 species and 4 groups of species (n = 40,135 individuals observed; Table 1).
The overall log-odds effect size across all studies was -1.54 (95% CI: -2.83, -0.26), suggesting no strong evidence of a disturbance effect on nesting birds caused by drones (Fig. 2; Appendix 1). However, heterogeneity in the overall model was large (Q = 8726.55; df = 30; P < 0.001). The mixed-effects meta-regression analysis indicated that altitude accounted for 13.05% of the heterogeneity, with > 50 m showing no strong evidence of disturbance (-2.72; 95% CI: -5.35, -0.08), whereas ≤ 50 m (-0.73; 95% CI: -2.17, 0.70) showed weak evidence of disturbance (Fig. 2; Appendix 1). Species nesting traits accounted for 25.45% of the heterogeneity. Ground colonial nesters showed no strong evidence of disturbance effect (-2.95; 95% CI: -4.88, -1.02), whereas non-ground solitary nesters showed strong evidence of a disturbance effect (3.89; 95% CI: 1.13, 6.65), indicating a clear difference in the odds of disturbance caused by drones depending on species nesting traits (Fig. 2).
We found no evidence of publication bias in the overall model (P = 0.21), the altitude model (P = 0.33), nor the nesting traits model (P = 0.32). Additionally, the funnel plot showed horizontal symmetry in each model (Appendix 2, Fig. A2.2). Moreover, the residual heterogeneity in the true effects followed a normal distribution as shown by a normal quantile-quantile (QQ) plot (Appendix 2, Fig. A2.1), further suggesting that model assumptions were met and were a good fit for the data.
We detected no evidence of a disturbance effect (< 0 log odds) on any species nesting trait group when the drone was flown above 50 m (Fig. 3). Conversely, when the drone was flown at or below 50 m, only ground colonial nesters show no strong evidence of disturbance (Fig. 3). The strongest disturbance effect size was observed in ground solitary and non-ground nesters when drones were flown at or below 50 m.
DISCUSSION
The results of this study indicate that the use of drones has an overall small disturbance effect on nesting birds. Individual studies’ effect sizes indicated strong evidence of no disturbance for any species group when the drone altitude was > 50 m, whereas the disturbance from drones at an altitude of ≤ 50 m depended on nesting traits. Disturbance effects were strongest for ground solitary and non-ground solitary nesters at altitudes of ≤ 50 m. Conversely, ground colonial nesters showed no evidence of disturbance effect regardless of the drone altitude. Two conclusions from these results are that the use of drones flown above 50 m provides a consistently low disturbance means to monitor nesting birds, supporting recommendations in Mesquita et al. (2021) and conclusions by Barr et al. (2020) and Rümmler et al. (2018), whereas studies in which flights were ≤ 50 m had mixed effect sizes, with the magnitude of the disturbance varying by species nesting traits.
Despite our finding of overall small disturbance effect on nesting birds caused by drones, heterogeneity was large across studies, with several studies reporting high levels of disturbance for some species (see Appendix 1). Thus, we expected these large effect sizes to increase the disturbance effect size in the overall model. However, this was not the case, likely due to most effects (22 out of 32) having small effect sizes (< 0 log odds). The largest effect size obtained in this analysis (5.98; 95% CI: 3.21, 8.76) was on a study conducted for breeding Common Redshanks, but ironically, the study concluded that drones provide less disturbance than other traditional surveying methods (Valle and Scarton 2020). However, results for altitude and species nesting traits as moderators did show larger effect sizes or confidence intervals overlapping zero, suggesting that these factors should be considered in drone survey methodology.
Nesting traits accounted for higher heterogeneity (25.45%) than did altitude (13.05%). This new finding was not expected because studies consistently attribute disturbance to drone altitude. However, no studies that we are aware of have assessed drone disturbance across species nesting traits, so the effects could have been masked by other variables not considered. Only ground colonial nesters showed strong evidence of no disturbance effects by drones regardless of altitude. The degree of disturbance exhibited by the ground-nesting species in our study is lower than that observed via traditional methods (e.g., ground surveys) in other studies in which breeding failure is commonly reported (Götmark 1992, Carney and Sydeman 1999). This may be due to ground predators (e.g., mammals) being more important to most ground-nesting birds than aerial predators (Richardson et al. 2009). Moreover, it has been suggested that traditional ground surveys create trails that lead to bird nests, facilitating access to predators (Skagen et al. 1999, Bushaw et al. 2020), which may further increase disturbance to ground nesters compared to drone surveys.
Our analysis was limited to 18 studies despite identifying at least 47 studies that examined disturbance of nesting birds. This is because, surprisingly, most studies did not report sample sizes for observed species, did not specify the number of birds that were disturbed out of the total, or did not provide the altitude at which the drone flew. We urge authors to report these factors at a minimum, to increase the utility of their work for future meta-analyses. Although a limited number of studies can lead to an under-representation of species and systems, and therefore to potential publication and reporting bias (Gurevitch et al. 2018), our models did not provide evidence of any biases. Importantly, our included studies were not restricted to a few species or locations (12 countries represented), and the cumulative number of birds observed across studies was large (n = 40,135). Notably, most included studies focused on large-sized birds, presumably because they are easier to detect with drone cameras and/or because they nest in areas more accessible to drone cameras. Nevertheless, the use of drones for monitoring nesting birds is becoming increasingly popular due to their efficacy and accessibility (Chabot et al. 2015, Lachman et al. 2020), and we expect the increasing number of studies using drones will continue to grow and expand to more species. In this sense, it is important that studies include a definition of disturbance or specify what is being considered as disturbance so the magnitude of effects and interpretations can be more easily tracked. Additionally, studies should consider (if applicable) assessing or reporting potential disturbance effects on chicks in nests, particularly when adults are flushed off nests.
The rapid advancement of technology incorporated into drones could reduce the extent of disturbance on nesting birds in future monitoring surveys. For example, throughout the relatively short time (last ~10 years) drones have been used in wildlife monitoring applications, the cost of high-end camera drones has dropped considerably and there have been increases in the battery life, GPS accuracy, and camera resolution of commercially available models (Abdelmaboud 2021). As camera resolution improves on commercially available cameras incorporated by drones, researchers may fly their survey missions at higher altitudes and capture images with an equal resolution to what would have previously necessitated flying missions at a lower altitude (Barnas et al. 2020). Distance between drones and nesting birds can be increased by using stabilized telephoto lenses in conjunction with high-resolution cameras, as smaller and higher quality cameras become available (Altena and Goedemé 2014, Borrelle and Fletcher 2017). The smaller and higher quality cameras can then be coupled with smaller drones to reduce visual impact, which has also been reported to influence disturbance in wildlife (Mulero-Pázmány et al. 2017). Also, propulsion systems may be selected to reduce noise signatures and may further reduce perceived threats (Sinibaldi and Marino 2013) thereby reducing disturbance on nesting birds.
Studies have shown empirically that there are negative effects on birds associated with investigator presence at nest sites (Borrelle and Fletcher 2017). These negative effects may result in abandonment of the nesting site, reduction in hatching success, reduction in breeding success, deaths of individual adults, and a reduction in local, regional, or total populations (Nisbet 2000). The use of drones to survey nesting birds does not eliminate the possibility for negative effects in some circumstances (Borrelle and Fletcher 2017); however, this meta-analysis demonstrates that if altitude and species traits are considered, drones can provide an effective means of collecting useful demographic and environmental data while reducing disturbance for nesting birds and reducing chances of injury or death for investigators.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
A. Cantu de Leija: writing, original draft and editing; conceptualization; investigation; methodology; data analysis; and visualization. R. E. Mirzadi: writing, original draft; conceptualization; investigation; and methodology. J. M. Randall: writing, original draft; conceptualization; and visualization. M. D. Portmann: writing, original draft; conceptualization; and investigation. E. J. Mueller: writing, original draft; conceptualization; and investigation. D. E. Gawlik: writing, review and editing.
ACKNOWLEDGMENTS
The authors were supported by Texas A&M University-Corpus Christi. The authors thank the editors and two anonymous reviewers for their detailed feedback and constructive comments. Funding for publication costs was provided by the Texas A&M University-Corpus Christi Mary and Jeff Bell Library, the Division of Research and Innovation, and the Office of the Provost.
DATA AVAILABILITY
The data that support the findings of this study are included in this article (Table 1). The R script used for data analysis is available by request to the corresponding author [AC].
LITERATURE CITED
Abdelmaboud, A. 2021. The internet of drones: requirements, taxonomy, recent advances, and challenges of research trends. Sensors 21:5718. https://doi.org/10.3390/s21175718
Afán, I., M. Máñez, and R. Díaz-Delgado. 2018. Drone monitoring of breeding waterbird populations: the case of the Glossy Ibis. Drones 2:42. https://doi.org/10.3390/drones2040042
Altena, B., and T. Goedemé. 2014. Assessing UAV platform types and optical sensor specifications. ISPRS Annals of Photogrammetry, Remote Sensing and Spatial Information Sciences II5:17-24. https://doi.org/10.5194/isprsannals-II-5-17-2014
Barnas, A. F., D. Chabot, A. J. Hodgson, D. W. Johnston, D. M. Bird, and S. N. Ellis-Felege. 2020. A standardized protocol for reporting methods when using drones for wildlife research. Journal of Unmanned Vehicle Systems 8:89-98. https://doi.org/10.1139/juvs-2019-0011
Barr, J. R., M. C. Green, S. J. DeMaso, and T. B. Hardy. 2020. Drone surveys do not increase colony-wide flight behaviour at waterbird nesting sites, but sensitivity varies among species. Scientific Reports 10:3781. https://doi.org/10.1038/s41598-020-60543-z
Bevan, E., S. Whiting, T. Tucker, M. Guinea, A. Raith, and R. Douglas. 2018. Measuring behavioral responses of sea turtles, saltwater crocodiles, and crested terns to drone disturbance to define ethical operating thresholds. PLOS ONE 13(3):e0194460. https://doi.org/10.1371/journal.pone.0194460
Blackmer, A. L., J. T. Ackerman, and G. A. Nevitt. 2004. Effects of investigator disturbance on hatching success and nest-site fidelity in a long-lived seabird, Leach’s Storm-Petrel. Biological Conservation 116:141-148. https://doi.org/10.1016/S0006-3207(03)00185-X
Borenstein, M., L. V. Hedges, J. P. T. Higgins, and H. R. Rothstein. 2009. Introduction to meta-analysis. John Wiley and Sons, Chichester, U.K. https://doi.org/10.1002/9780470743386
Borrelle, S. B., and A. T. Fletcher. 2017. Will drones reduce investigator disturbance to surface-nesting birds? Marine Ornithology 45:89-94. http://www.marineornithology.org/PDF/45_1/45_1_89-94.pdf
Brisson-Curadeau, É., D. Bird, C. Burke, D. A. Fifield, P. Pace, R. B. Sherley, and K. H. Elliott. 2017. Seabird species vary in behavioural response to drone census. Scientific Reports 7:17884. https://doi.org/10.1038/s41598-017-18202-3
Burger, J. 1981. The effect of human activity on birds at a coastal bay. Biological Conservation 21:231-241. https://doi.org/10.1016/0006-3207(81)90092-6
Bushaw, J. D., K. M. Ringelman, M. K. Johnson, T. Rohrer, and F. C. Rohwer. 2020. Applications of an unmanned aerial vehicle and thermal-imaging camera to study ducks nesting over water. Journal of Field Ornithology 91:409-420. https://doi.org/10.1111/jofo.12346
Carey, M. J. 2009. The effects of investigator disturbance on procellariiform seabirds: a review. New Zealand Journal of Zoology 36:367-377. https://doi.org/10.1080/03014220909510161
Carney, K. M., and W. J. Sydeman. 1999. A review of human disturbance effects on nesting colonial waterbirds. Waterbirds 22:68-79. https://doi.org/10.2307/1521995
Chabot, D., S. R. Craik, and D. M. Bird. 2015. Population census of a large Common Tern colony with a small unmanned aircraft. PLOS ONE 10:e0122588. https://doi.org/10.1371/journal.pone.0122588
Egger, M., G. D. Smith, M. Schneider, and C. Minder. 1997. Bias in meta-analysis detected by a simple, graphical test. BMJ 315:629-634. https://doi.org/10.1136/bmj.315.7109.629
Frederick, P. C., T. Towles, R. J. Sawicki, and G. T. Bancroft. 1996. Comparison of aerial and ground techniques for discovery and census of wading bird (Ciconiiformes) nesting colonies. Condor 98:837-841. https://doi.org/10.2307/1369865
Fudala, K., and R. J. Bialik. 2022. The use of drone-based aerial photogrammetry in population monitoring of Southern Giant Petrels in ASMA 1, King George Island, maritime Antarctica. Global Ecology and Conservation 33:e01990. https://doi.org/10.1016/j.gecco.2021.e01990
Gallego, D., and J. H. Sarasola. 2021. Using drones to reduce human disturbance while monitoring breeding status of an endangered raptor. Remote Sensing in Ecology and Conservation 7:550-561. https://doi.org/10.1002/rse2.206
Gibbs, J. P., S. Woodward, M. L. Hunter, and A. E. Hutchinson. 1988. Comparison of techniques for censusing great blue heron nests. Journal of Field Ornithology 59:130-134.
Götmark, F. 1992. The effects of investigator disturbance on nesting birds. Pages 63-104 in D. M. Power, editor. Current ornithology. Springer, Boston, Massachusetts, USA. https://doi.org/10.1007/978-1-4757-9921-7_3
Gurevitch, J., J. Koricheva, S. Nakagawa, and G. Stewart. 2018. Meta-analysis and the science of research synthesis. Nature 555:175-182. https://doi.org/10.1038/nature25753
Junda, J. H., E. Greene, D. Zazelenchuk, and D. M. Bird. 2016. Nest defense behaviour of four raptor species (Osprey, Bald Eagle, Ferruginous Hawk, and Red-tailed Hawk) to a novel aerial intruder - a small rotary-winged drone. Journal of Unmanned Vehicle Systems 4:217-227. https://doi.org/10.1139/juvs-2016-0004
Klein, M. L. 1993. Waterbird behavioral responses to human disturbances. Wildlife Society Bulletin 21:31-39.
Lachman, D., C. Conway, K. Vierling, and T. Matthews. 2020. Drones provide a better method to find nests and estimate nest survival for colonial waterbirds: a demonstration with Western Grebes. Wetlands Ecology and Management 28:837-845. https://doi.org/10.1007/s11273-020-09743-y
Linchant, J., J. Lisein, J. Semeki, P. Lejeune, and C. Vermeulen. 2015. Are unmanned aircraft systems (UASs) the future of wildlife monitoring? A review of accomplishments and challenges. Mammal Review 45:239-252. https://doi.org/10.1111/mam.12046
López-López, J. A., F. Marín-Martínez, J. Sánchez-Meca, W. Van den Noortgate, and W. Viechtbauer. 2014. Estimation of the predictive power of the model in mixed-effects meta-regression: a simulation study. British Journal of Mathematical and Statistical Psychology 67:30-48. https://doi.org/10.1111/bmsp.12002
Lyons, M. B., K. J. Brandis, N. J. Murray, J. H. Wilshire, J. A. McCann, R. T. Kingsford, and C. T. Callaghan. 2019. Monitoring large and complex wildlife aggregations with drones. Methods in Ecology and Evolution 10:1024-1035. https://doi.org/10.1111/2041-210X.13194
Martin, T. E., and G. R. Geupel. 1993. Nest-monitoring plots: methods for locating nests and monitoring success. Journal of Field Ornithology 64:507-519.
McEvoy, J. F., G. P. Hall, and P. G. McDonald. 2016. Evaluation of unmanned aerial vehicle shape, flight path and camera type for waterfowl surveys: disturbance effects and species recognition. PeerJ 4:e1831. https://doi.org/10.7717/peerj.1831
McKellar, A. E., N. G. Shephard, and D. Chabot. 2020. Dual visible‐thermal camera approach facilitates drone surveys of colonial marshbirds. Remote Sensing in Ecology and Conservation 7:214-226. https://doi.org/10.1002/rse2.183
Mesquita, G. P., J. D. Rodríguez-Teijeiro, S. A. Wich, and M. Mulero-Pßzmány. 2021. Measuring disturbance at swift breeding colonies due to the visual aspects of a drone: a quasi-experiment study. Current Zoology 67:157-163. https://doi.org/10.1093/cz/zoaa038
Mulero-Pázmány, M., S. Jenni-Eiermann, N. Strebel, T. Sattler, J. J. Negro, and Z. Tablado. 2017. Unmanned aircraft systems as a new source of disturbance for wildlife: a systematic review. PLoS ONE 12(6):e0178448. https://doi.org/10.1371/journal.pone.0178448
Nisbet, I. C. T. 2000. Disturbance, habituation, and management of waterbird colonies. Waterbirds 23:312-332.
Page, M. J., J. E. McKenzie, P. M. Bossuyt, I. Boutron, T. C. Hoffmann, C. D. Mulrow, L. Shamseer, J. M. Tetzlaff, E. A. Akl, S. E. Brennan, R. Chou, J. Glanville, J. M. Grimshaw, A. Hróbjartsson, M. M. Lalu, T. Li, E. W. Loder, E. Mayo-Wilson, S. McDonald, L. A. McGuinness, L. A. Stewart, J. Thomas, A. C. Tricco, V. A. Welch, P. Whiting, and D. Moher. 2021. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 2021 372:n71. https://doi.org/10.1136/bmj.n71
Pimm, S. L., S. Alibhai, R. Bergl, A. Dehgan, C. Giri, Z. Jewell, L. Joppa, R. Kays, and S. Loarie. 2015. Emerging technologies to conserve biodiversity. Trends in Ecology and Evolution 30:685-696. https://doi.org/10.1016/j.tree.2015.08.008
Powell, G. V. N., and A. H. Powell. 1986. Reproduction by Great White Herons Ardea herodias in Florida Bay as an indicator of habitat quality. Biological Conservation 36:101-113. https://doi.org/10.1016/0006-3207(86)90001-7
R Core Team. 2021. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Richardson, T. W., T. Gardali, and S. H. Jenkins. 2009. Review and meta-analysis of camera effects on avian nest success. Journal of Wildlife Management 73:287-293. https://doi.org/10.2193/2007-566
Rodgers, J. A., Jr., P. S. Kubilis, and S. A. Nesbitt. 2005. Accuracy of aerial surveys of waterbird colonies. Waterbirds 28:230-237. https://doi.org/10.1675/1524-4695(2005)028[0230:AOASOW]2.0.CO;2
Rümmler, M.-C., O. Mustafa, J. Maercker, H.-U. Peter, and J. Esefeld. 2016. Measuring the influence of unmanned aerial vehicles on Adélie Penguins. Polar Biology 39:1329-1334. https://doi.org/10.1007/s00300-015-1838-1
Rümmler, M.-C., O. Mustafa, J. Maercker, H.-U. Peter, and J. Esefeld. 2018. Sensitivity of Adélie and Gentoo penguins to various flight activities of a micro UAV. Polar Biology 41:2481-2493. https://doi.org/10.1007/s00300-018-2385-3
Ryckman, M. D., K. Kemink, C. J. Felege, B. Darby, G. S. Vandeberg, and S. N. Ellis-Felege. 2022. Behavioral responses of Blue-winged Teal and Northern Shoveler to unmanned aerial vehicle surveys. PLOS ONE 17(1):e0262393. https://doi.org/10.1371/journal.pone.0262393
Sasse, D. B. 2003. Job-related mortality of wildlife workers in the United States, 1937-2000. Wildlife Society Bulletin 31:1015-1020.
Scholten, B. D., A. R. Beard, H. Choi, D. M. Baker, M. E. Caulfield, and D. S. Proppe. 2020. Short-term exposure to unmanned aerial vehicles does not alter stress responses in breeding Tree Swallows. Conservation Physiology 8(1):coaa080. https://doi.org/10.1093/conphys/coaa080
Shewring, M. P., and J. O. Vafidis. 2021. Using UAV-mounted thermal cameras to detect the presence of nesting Nightjar in upland clear-fell: a case study in South Wales, UK. Ecological Solutions and Evidence 2(1):e12052. https://doi.org/10.1002/2688-8319.12052
Sinibaldi, G., and L. Marino. 2013. Experimental analysis on the noise of propellers for small UAV. Applied Acoustics 74:79-88. https://doi.org/10.1016/j.apacoust.2012.06.011
Skagen, S. K., T. R. Stanley, and M. B. Dillon. 1999. Do mammalian nest predators follow human scent trails in the shortgrass prairie? Wilson Bulletin 111:415-420. https://sora.unm.edu/sites/default/files/journals/wilson/v111n03/p0415-p0420.pdf
Sutherland, W. J., I. Newton, and R. Green. 2004. Bird ecology and conservation: a handbook of techniques. Oxford University Press, New York, New York, USA. https://doi.org/10.1093/acprof:oso/9780198520863.001.0001
Tremblay, J., and L. N. Ellison. 1979. Effects of human disturbance on breeding of Black-crowned Night Herons. Auk 96:364-369. https://sora.unm.edu/sites/default/files/journals/auk/v096n02/p0364-p0369.pdf
Valle, R. G., and F. Scarton. 2020. Drones improve effectiveness and reduce disturbance of censusing Common Redshanks Tringa totanus breeding on salt marshes. Ardea 107:275-282. https://doi.org/10.5253/arde.v107i3.a3
Viechtbauer, W. 2010. Conducting meta-analyses in R with the metafor Package. Journal of Statistical Software 36. https://doi.org/10.18637/jss.v036.i03
Weimerskirch, H., A. Prudor, and Q. Schull. 2018. Flights of drones over sub-Antarctic seabirds show species- and status-specific behavioural and physiological responses. Polar Biology 41:259-266. https://doi.org/10.1007/s00300-017-2187-z
Wen, D., L. Su, Y. Hu, Z. Xiong, M. Liu, and Y. Long. 2021. Surveys of large waterfowl and their habitats using an unmanned aerial vehicle: a case study on the Siberian Crane. Drones 5:102. https://doi.org/10.3390/drones5040102
Wickham, H., M. Averick, J. Bryan, W. Chang, L. D. McGowan, R. François, G. Grolemund, A. Hayes, L. Henry, J. Hester, M. Kuhn, T. L. Pedersen, E. Miller, S. M. Bache, K. Müller, J. Ooms, D. Robinson, D. P. Seidel, V. Spinu, K. Takahashi, D. Vaughan, C. Wilke, K. Woo, and H. Yutani. 2019. Welcome to the Tidyverse. Journal of Open Source Software 4:1686. https://doi.org/10.21105/joss.01686
Zbyryt A., L. Dylewski, F. Morelli, T. H. Sparks, and P. Tryjanowski. 2021. Behavioural responses of adult and young White Storks Ciconia cinocia in nests to an unmanned aerial vehicle. Acta Ornithologica 55(2):243-251. https://doi.org/10.3161/00016454AO2020.55.2.009
Fig. 1
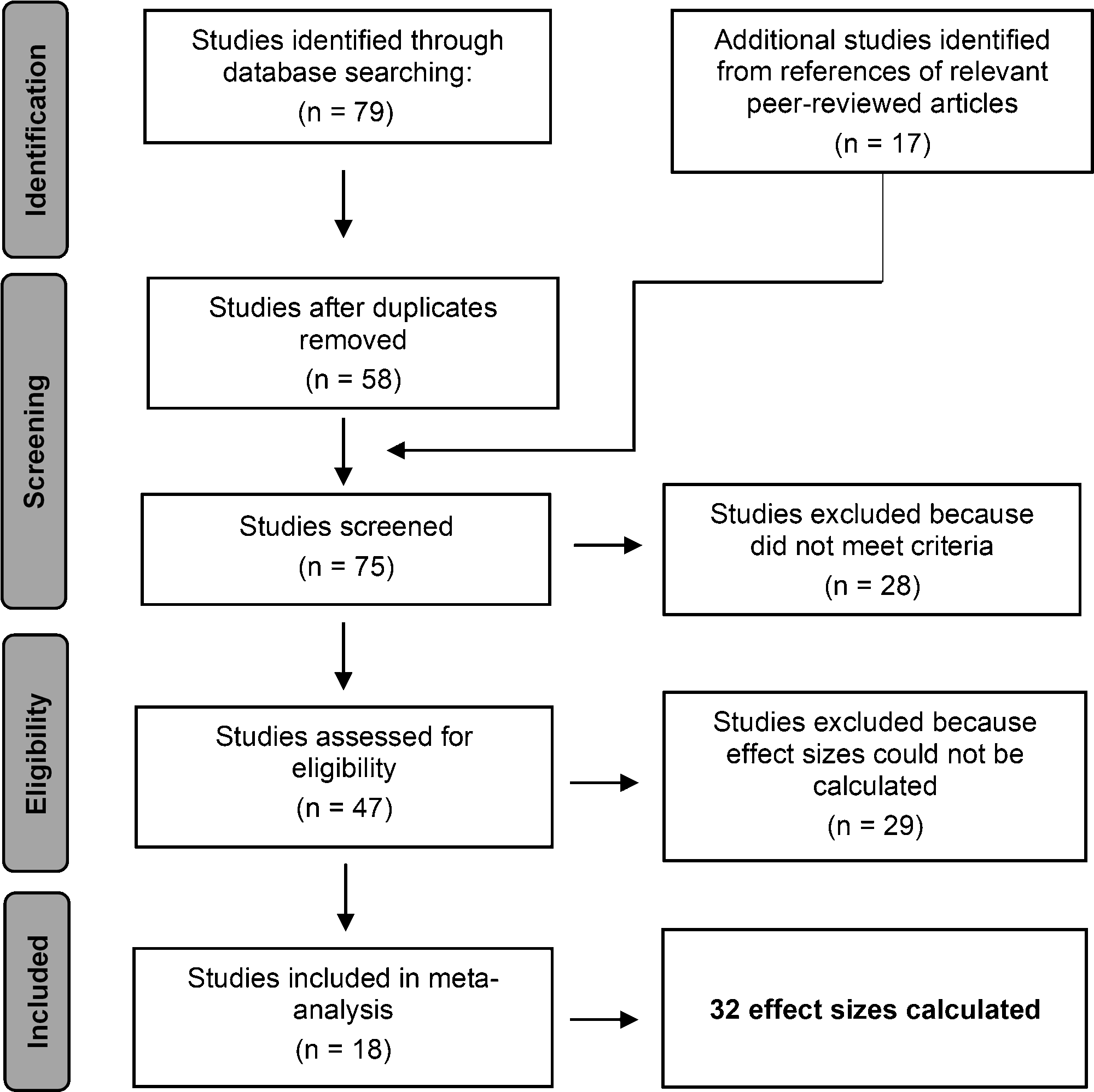
Fig. 1. Preferred reporting items for systematic reviews and meta-analyses (PRISMA) flow diagram displaying the number of studies considered at each step of the study selection process for meta-analysis.<.span>
Fig. 2
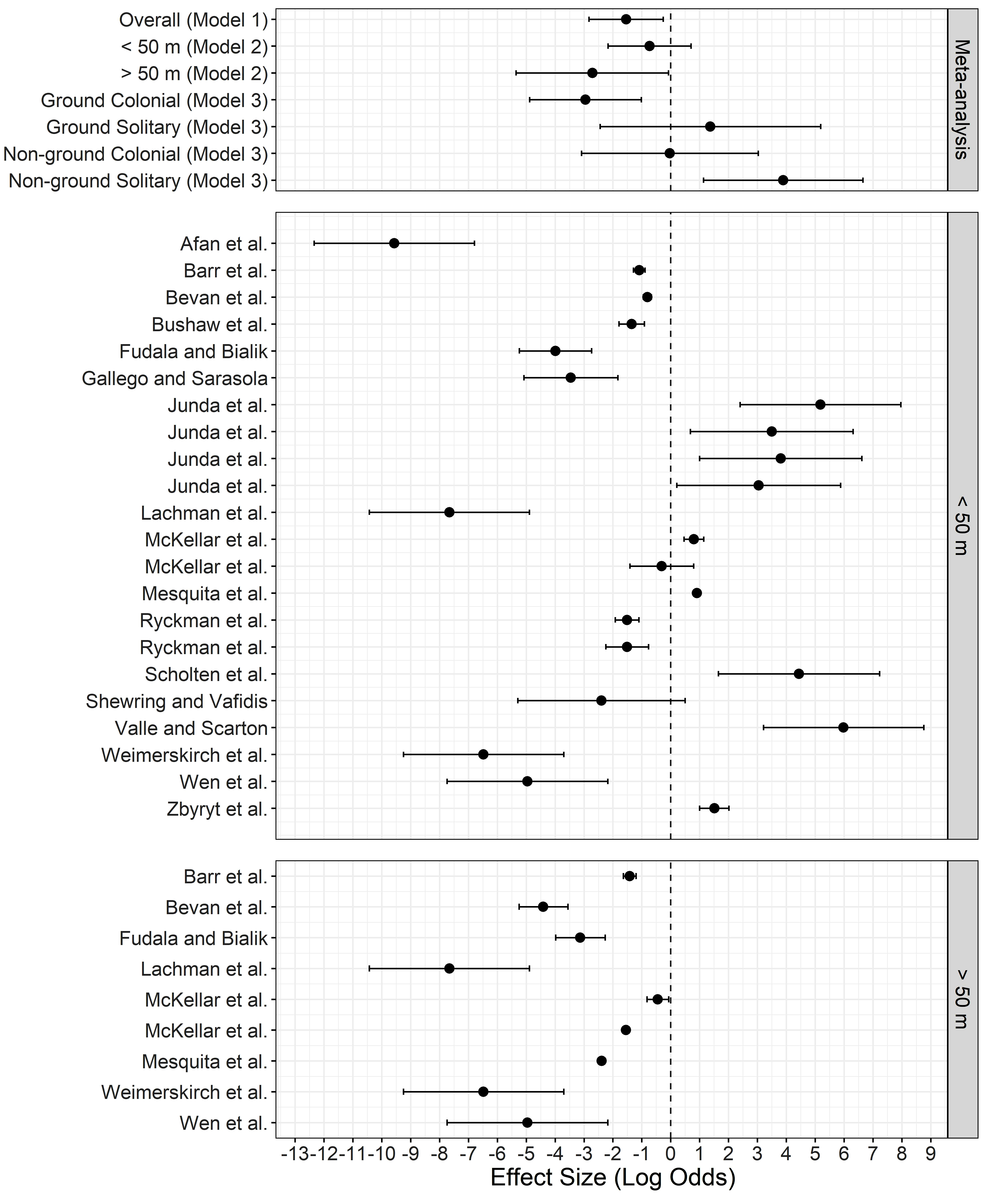
Fig. 2. Forest plot of effect sizes (points) and 95% confidence intervals (whiskers) for overall random effects meta-analysis and meta-regression models (top panel) and for each study partitioned by drone altitude (mid and bottom panels). Effect size is calculated as the log-odds ratio of reported disturbed vs. non-disturbed nesting birds in each study. Increasing positive log-odds values indicate stronger evidence of drone disturbance effects and increasing negative values indicate stronger evidence of no disturbance effect. Confidence intervals crossing zero suggest weak evidence of disturbance/no disturbance effect.
Fig. 3
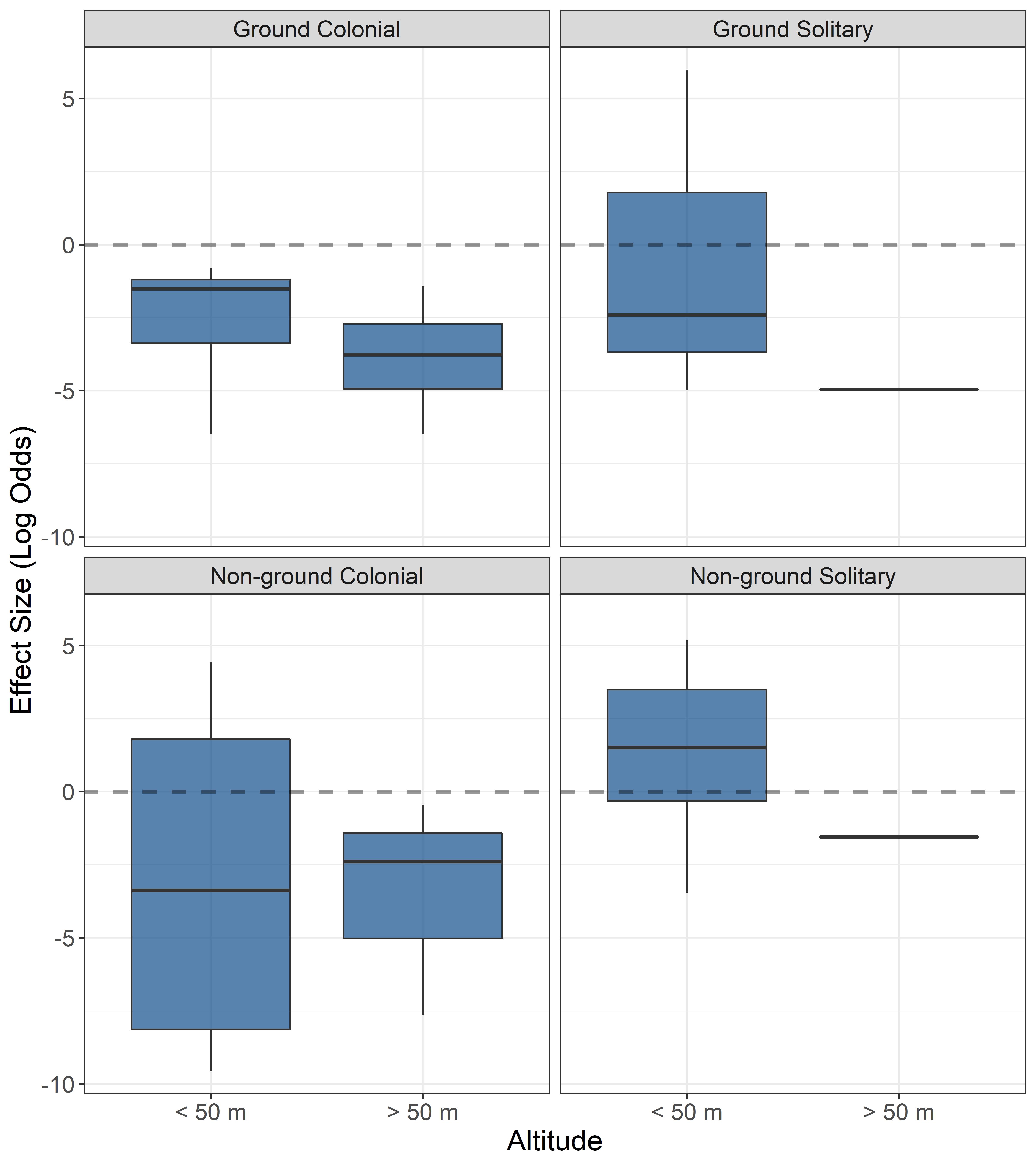
Fig. 3. Summary of individual studies’ disturbance effect sizes on nesting birds by altitude and species nesting traits. Effect size is calculated as the log-odds ratio of reported disturbed vs. non-disturbed nesting birds in each study. Increasing positive log-odds values indicate stronger evidence of drone disturbance effects and increasing negative values indicate stronger evidence of no disturbance effect. Values crossing zero suggest weak evidence of disturbance/no disturbance effect. For each boxplot, the medium line indicates the median; top and bottom lines of the box indicate the upper and lower interquartile range; whiskers indicate the max and min values.
Table 1
Table 1. List of included studies and data collected for meta-analysis.
| Study | Year | Species | Group | Total birds | Disturbed | †Altitude flown (m) | Altitude category | ||
| Afán et al. | 2018 | Glossy Ibis (Plegadis falcinellus) |
Non-ground Colonial | 7134 | 0 | 50 | ≤ 50 m | ||
| Barr et al. | 2020 | Wading birds | Ground Colonial | 510 | 128 | 46 | ≤ 50 m | ||
| Bevan et al. | 2018 | Crested tern (Thalasseus bergii) |
Ground Colonial | 926 | 285 | 50, 45, 40 | ≤ 50 m | ||
| Bushaw et al. | 2020 | Waterfowl | Non-ground Solitary | 118 | 24 | 31, 5 | ≤ 50 m | ||
| Fudala and Bialik | 2022 | Southern Giant Petrel (Macronectes giganteus) |
Ground Colonial | 137 | 2 | 50, 30 | ≤ 50 m | ||
| Gallego and Sarasola | 2021 | Chaco Eagle (Buteogallus coronatus) |
Non-ground Solitary | 48 | 1 | 5-10 | ≤ 50 m | ||
| Junda et al. | 2016 | Osprey (Pandion haliaetus) |
Non-ground Solitary | 88 | 88 | 3 | ≤ 50 m | ||
| Junda et al. | 2016 | Bald Eagle (Haliaeetus leucocephalus) |
Non-ground Solitary | 16 | 16 | 3-6 | ≤ 50 m | ||
| Junda et al. | 2016 | Ferruginous Hawk (Buteo regalis) |
Non-ground Solitary | 22 | 22 | 3-6 | ≤ 50 m | ||
| Junda et al. | 2016 | Red-tailed Hawk (Buteo jamaicensis) |
Non-ground Solitary | 10 | 10 | 3-6 | ≤ 50 m | ||
| Lachman et al. | 2020 | Western Grebe (Aechmophorus occidentalis) |
Non-ground Colonial | 1059 | 0 | 50-10 | ≤ 50 m | ||
| McKellar et al. | 2020 | Franklin’s Gull (Leucophaeus pipixcan) |
Non-ground Solitary | 152 | 105 | 45 | ≤ 50 m | ||
| McKellar et al. | 2020 | Black Tern (Chlidonias niger) |
Non-ground Solitary | 12 | 5 | 45 | ≤ 50 m | ||
| Mesquita et al. | 2021 | Swifts | Non-ground Colonial | 12000 | 8550 | 50, 47.17, 43.01, 29.15, 26.93, 25.50, 25 | ≤ 50 m | ||
| Ryckman et al. | 2022 | Blue-winged Teal (Anas discors) |
Ground Colonial | 151 | 27 | 45 | ≤ 50 m | ||
| Ryckman et al. | 2022 | Northern Shoveler (Spatula clypeata) |
Ground Colonial | 46 | 8 | 45 | ≤ 50 m | ||
| Scholten et al. | 2020 | Tree Swallow (Tachycineta bicolor) |
Non-ground Colonial | 42 | 42 | 1.5 | ≤ 50 m | ||
| Shewring and Vafidis | 2021 | European Nightjar (Caprimulgus europaeus) |
Ground Solitary | 5 | 0 | 50-5 | ≤ 50 m | ||
| Valle and Scarton | 2020 | Redshank (Tringa totanus) |
Ground Solitary | 198 | 198 | 10 | ≤ 50 m | ||
| Weimerskirch et al. | 2018 | Seabirds | Ground Colonial | 324 | 0 | 50, 25, 10, 10-3 | ≤ 50 m | ||
| Wen et al. | 2021 | Siberian Crane (Grus leucogeranus) |
Ground Solitary | 71 | 0 | 50 | ≤ 50 m | ||
| Zbyryt et al. | 2021 | White Stork (Ciconia ciconia) |
Non-ground Solitary | 101 | 83 | 20-1 | ≤ 50 m | ||
| Barr et al. | 2020 | Wading birds | Ground Colonial | 510 | 99 | 122, 91, 61 | > 50 m | ||
| Bevan et al. | 2018 | Crested tern | Ground Colonial | 455 | 5 | 70, 60 | > 50 m | ||
| Fudala and Bialik | 2022 | Southern Giant Petrel | Ground Colonial | 130 | 5 | 70, 100, 130, 200 | > 50 m | ||
| Lachman et al. | 2020 | Western Grebe | Non-ground Colonial | 1059 | 0 | 60-51 | > 50 m | ||
| McKellar et al. | 2021 | Western Grebe | Non-ground Colonial | 113 | 44 | 60 | > 50 m | ||
| McKellar et al. | 2021 | Franklin’s Gull | Non-ground Solitary | 3803 | 665 | 60, 120 | > 50 m | ||
| Mesquita et al. | 2021 | Swifts | Non-ground Colonial | 10500 | 881 | 64.03, 61.03, 52.20, 50.99, 50.25 | > 50 m | ||
| Weimerskirch et al. | 2018 | Seabirds | Ground Colonial | 324 | 0 | > 50 | > 50 m | ||
| Wen et al. | 2021 | Siberian Crane | Ground Solitary | 71 | 0 | 150, 100 | > 50 m | ||
| †Altitude flown (m) = reported values or range of values used in each study. | |||||||||