







The following is the established format for referencing this article:
Newcombe, P. B., A. Forsyth, H. Brumberg, and A. Whitworth. 2023. Climate-resilient conservation strategies for an endemic forest bird, the Black-cheeked Ant-Tanager. Journal of Field Ornithology 94(2):2.ABSTRACT
As biodiversity declines and climate change causes shifts in species distribution, the knowledge of species’ ecological needs is vital to conserve biodiversity. On Costa Rica’s Osa Peninsula and its adjacent forests, a rich mosaic of ecosystems hosting numerous threatened and endemic species, conservationists lack clarity on the basic habitat requirements of the endemic Black-cheeked Ant-Tanager (Habia atrimaxillaris). Numerous attempts have been made to understand its habitat requirements, resulting in contradictory conclusions. This study integrates new field data, thousands of community science observations, and comments in historical literature ranging over 50 years to complement the more localized studies. We explore the species’ habitat requirements, diet, and distribution in protected areas and biological corridors to better understand the species’ conservation needs and, in doing so, suggest a strategy to protect the Black-cheeked Ant-Tanager in a changing climate. We illustrate that Black-cheeked Ant-Tanagers occur in secondary forests, which they also use for foraging and nesting, suggesting that the conservation and restoration of secondary forests may help protect this range-restricted forest bird, especially through a targeted conservation strategy within biological corridors to build connectivity with higher elevations.
RESUMEN
A medida que la biodiversidad disminuye y el cambio del clima causa cambios en las distribuciones de las especies, el conocimiento de las necesidades ecológicas de las especies es vital para la conservación de la biodiversidad. En la península de Osa en Costa Rica y sus bosques adyacentes, un mosaico diverso de ecosistemas en el cual habitan un alto número de especies amenazadas y endémicas, los conservacionistas no tienen claridad de los requerimientos básicos de hábitat de la especie endémica Estrategias de conservación resilientes al clima para un ave de bosque endémica, Habia atrimaxillaris. Ha habido muchos intentos para entender sus requerimientos de hábitat, con resultados contradictorios. Este estudio integra nueva información de campo, miles de observaciones de ciencia comunitaria y comentarios en la literatura histórica con un rango de más de 50 años para complementar los estudios más localizados. Exploramos los requerimientos de hábitat de la especie, su dieta y la distribución en áreas protegidas y corredores biológicos con el fin de comprender mejor las necesidades de conservación de la especie y sugerir una estrategia para proteger a Habia atrimaxillaris en un clima cambiante. Mostramos que Habia atrimaxillaris ocurre en bosques secundarios, los cuales también usan para forrajear y anidar, sugiriendo que la conservación y la restauración de bosques secundarios puede ayudar a proteger esta especie de ave de bosque con un rango restringido, especialmente a través de estrategias de conservación focalizadas dentro de corredores biológicos para construir conectividad con elevaciones mayores.
INTRODUCTION
The Black-cheeked Ant-Tanager (Habia atrimaxillaris) is a species endemic to the Osa Peninsula and adjacent forests of southwestern Costa Rica, an area that retains wet forest, large expanses of mangroves, and numerous threatened and endemic species (Rocha et al. 2006, Lobo et al. 2008, Cornejo et al. 2012). The region has experienced deforestation, especially conversion to pastureland, over the past century. In recent decades, however, forest cover has increased in the Osa, mostly because of forest regeneration on abandoned pasture lands (Sanchez-Azofeifa et al. 2002; Furey et al., https://doi.org/10.6084/m9.figshare.19337912.v1).
In a changing climate, many birds’ ranges are shifting along elevational gradients (Freeman and Class Freeman 2014, Freeman et al. 2018). These shifts require bird dispersal to new areas, and forested corridors have been shown to facilitate dispersal in forest specialist birds (Gillies and St. Clair 2008). In the context of a sixth mass extinction, biodiversity conservation efforts are highly important (Ceballos et al. 2020). Conservation corridors have received increasing focus on the Osa Peninsula, but in order to devise a strategic, successful corridor system, conservationists must understand the habitat needs of range-restricted species. Therefore, better understanding Black-cheeked Ant-Tanager ecology could improve efforts to conserve this endemic species, especially since range-restricted forest birds, like the Black-cheeked Ant-Tanager, tend to be most vulnerable to climate change (Harris and Pimm 2008). Such an effort may help protect a host of other species that use the same habitat.
Numerous habitat assessments have yet to lead to consensus on the requirements of the Black-cheeked Ant-Tanager. One study reported that Black-cheeked Ant-Tanagers were restricted to old-growth forest (including primary forest, mature forest, and old secondary forest), adding that Black-cheeked Ant-Tanagers avoid forest edges and do not use young secondary growth or gallery forest, even when adjacent to closed forest (Cornils et al. 2015). Other studies have reported that Black-cheeked Ant-Tanagers prefer mature forest, but also use secondary forest: a report suggested that Black-cheeked Ant-Tanagers are more abundant in early successional than late successional forest, but at their greatest density in primary forest (Leavelle 2013). Another report suggested that Black-cheeked Ant-Tanagers preferred forest with closed canopy and forest edge, but speculated that steep terrain and presence of streams or rivers could be important habitat characteristics (Jones et al. 2009). One study reported that they occur in fragmented forest (defined as having < 80% forest cover), though less densely than in forests with greater cover (Wilson et al. 2011). Another paper reported that Black-cheeked Ant-Tanagers had not recolonized one particular site after six years of reforestation and is a “true forest specialist,” although the authors find that the species does occur in other secondary forest sites in the area (Schulze et al. 2019). In fact, Black-cheeked Ant-Tanager habitat classifications sweep the entire spectrum of forest types, with one paper including the Black-cheeked Ant-Tanager on a list of “specialists on early successional vegetation” (Sandoval et al. 2019). We suggest that combining diverse data sources from across the geographic range may help resolve some of the contradictions of these site-restricted surveys.
Because the multiple quantitative assessments of Black-cheeked Ant-Tanagers have offered few concrete conclusions, we aim to advance the understanding of Black-cheeked Ant-Tanager ecology and conservation biology by complementing the existing surveys and population estimates with observational evidence from two field seasons, thousands of community science observations, and reference to historical and anecdotal literature. This multi-datum and range-wide analysis may allow for a more complete snapshot of the species’ habitat tolerances. We evaluate the Black-cheeked Ant-Tanagers habitat requirements and climate resiliency across a partially protected landscape and suite of biological corridors in order to advise ongoing conservation actions (Guillermo 2017, Sistema Nacional de Áreas de Conservación 2018; Fig. 1, Appendix 3).
METHODS
Black-cheeked Ant-Tanagers were observed anecdotally during two fields seasons of two weeks each, in June 2017 and June–July 2018. The observer (PBN) documented food sources whenever possible, along with GPS coordinates, forest type, proximity to the nearest river, and other data.
Habitat
Lead author PBN noted the forest type of every encounter of a Black-cheeked Ant-Tanager individual at the Osa Conservation Campus (formerly known as Piro Biological Station; 8.40388 N, 83.336618 W), managed by the nonprofit organization Osa Conservation (see Whitworth et al. 2021 for a detailed description of the site), while walking the entire trail system, on the trails, with Zeiss Terra ED 8x42 binoculars and listening for vocalizations. Forest was classified as secondary or mature based on examination of the surrounding vegetation and confirmed with Osa Conservation’s forest map (Villalobos and Ramírez, date unknown); when the observer was not certain, generally because the sighting was on or close to the border, the forest type was classified as “border.” Separately, using the coordinates in Google Earth, we measured the distance to the nearest river and classified whether each was within 200 m of a river or in forest edge (whether of primary or secondary forest). Identifications were made with visual confirmation, allowing for certainty. We also included verifiable observations from experienced birders during June and July 2018. No effort was made to survey habitats proportionally; rather, we simply gathered as many sightings as possible with the goal of showing which habitats the Black-cheeked Ant-Tanagers occurred in.
We searched online for literature on the Black-cheeked Ant-Tanager for qualitative mention of habitat in studies, books, or other literature (including historical literature, with a focus on specific diet or nesting information) in order to develop a collection of relevant literature. Nesting observations were included to determine the use of various types of forest by more than just transient individuals. For this collection of literature (also referred to in the following diet section), which we used to more comprehensively understand Black-cheeked Ant-Tanager habitat needs, we included the following sources: Slud 1964, Willis 1972, Stiles and Skutch 1989, Huber et al. 2008, and Sandoval and Gallo 2009.
Diet
We compiled information on Black-cheeked Ant-Tanager diet from a variety of observational sources. PBN collected field observations of Black-cheeked Ant-Tanagers consuming food at the Osa Conservation property during the same time range as described above, by observing the Black-cheeked Ant-Tanagers for as long as possible (typically, 1-15 minutes per group, but staying near the trail while following them at a distance of, generally, about 20 ft) and noting any time that an individual engaged in an obvious foraging behavior (e.g., sallying for insects, holding an arthropod in its bill) or eating fruit. This analysis is not meant to be complete; we recognize that observations of insect consumption, in particular, may not always have been apparent, and we do not seek to compare relative consumption. We incorporated community science data through photos submitted to the Cornell Lab of Ornithology’s Macaulay Library, in which PBN viewed each Black-cheeked Ant-Tanager photo submitted to the library as of 24 January 2021 and noted any time a Black-cheeked Ant-Tanager was photographed foraging. We also searched literature on the Black-cheeked Ant-Tanager for mention of diet in a similar way as described in the previous section on habitat, and used the same sources collected in the habitat section.
Spatial analysis
Complementing field observations, 3344 community science observations of Black-cheeked Ant-Tanagers were drawn from the eBird database in early 2021, with no date range filter. All eBird checklists were used, including ones with short observation periods and from repeated locations, in order to include all data available. At each of the points (i.e., the location of a checklist recording a sighting of one or multiple birds) of the 3344 observations where Black-cheeked Ant-Tanagers were recorded in eBird, land use was extracted in QGIS by using a regional land use map created in collaboration with NASA DEVELOP (Shrestha et al. 2018; Furey et al., https://doi.org/10.6084/m9.figshare.19337912.v1). Each of the points was also classified by whether it fell within a Costa Rican protected area (national park, forest reserve, or wildlife refuge) or biological corridor, in order to inform basic analysis on protected area status and connectivity of Black-cheeked Ant-Tanager occurrence, by using the Digital Atlas of Costa Rica and SINAC (National System of Conservation Areas) maps. The observations are likely biased in frequency by the locations where birders tend to submit checklists from (as well as a multitude of other sampling effort factors), but still represent occurrence, if not relative occurrence, in those habitats.
RESULTS
Habitat
Our field observations consisted of 41 encounters with Black-cheeked Ant-Tanager individuals. Of these, 27 individuals occurred in secondary forest (65.9%), seven in primary forest (17.1%), and seven on the border (17.1%). Of all Black-cheeked Ant-Tanager observations, 17 individuals occurred in forest edge (41.5%) and 38 were within 200 m of a river (92.7%).
The 3344 community science observations of Black-cheeked Ant-Tanagers drawn from the eBird database represent birds from across the range, many years, and a wide array of habitats (Fig. 1, Appendix 1). A total of 68.87% (2303) of eBird checklists that included Black-cheeked Ant-Tanagers were in secondary forest. Of these, 214 were stationary checklists. In addition, 10.05% (336) of checklists with Black-cheeked Ant-Tanagers were based in mature rainforest, 9.54% (319) in grassland, and 8.43% (282) in exposed/urban areas. Few Black-cheeked Ant-Tanager checklists occurred in monocultures, such as teak/melina (.27%, or nine) and oil palm (.39%, or 13).
Anecdotal observations dating back over 50 years provide another perspective on Black-cheeked Ant-Tanager habitat use, including for nesting habitat. A description of the Black-cheeked Ant-Tanager’s nest noted that one was found “at the edge of secondary forest,” a second “in young secondary forest” 1.5 m from a river, and a third “at the top of a creek bank inside a riverside secondary forest” (Sandoval and Gallo 2009). The authors wrote that, based on the habitat of these nest observations, “disturbed habitats (e.g., forest edges, secondary forest)” are “apparent habitat preferences” (Sandoval and Gallo 2009). Another nest description paper described a nest in a “primary gorge forest” (Huber et al. 2008). Historical literature also included mention of Black-cheeked Ant-Tanager habitat. Willis described the species as tending to inhabit “broken forests and tall second growth” more than “unbroken” forest (Willis 1972). In his book about the birds of Costa Rica, Slud described it as occurring in primary forest, advanced second growth, and “a little in broken, selectively logged forest” (Slud 1964). Another book states that Black-cheeked Ant-Tanagers depend on large or interconnected intact forest (Stiles and Skutch 1989).
Diet
Several observations were made of foraging Black-cheeked Ant-Tanagers. A Black-cheeked Ant-Tanager was photographed foraging on the fruit of Carludovica palmata, a member of the Cyclanthaceae family with pulpy, orange berries common in degraded and secondary habitat. It had been foraging with a mixed species flock and calling in sync with other members of the flock, but became silent and remained perched on a stalk, foraging on the fruit for an extended period of time while the flock continued on. The plant was situated in secondary forest, just three feet from the research station’s clearing. Black-cheeked Ant-Tanagers were observed on the same species of plant repeatedly, sometimes foraging in dry leaves at the end of the Carludovica palmata stalk and often working their way along stalks of similar plants. Black-cheeked Ant-Tanagers were also observed gleaning insects from a wide variety of plants, including Piper guanacastense, sallying for insects and consuming arthropods that looked like ants. Separately, Black-cheeked Ant-Tanagers were observed visiting army ant swarms.
The search on the Macaulay Library (Appendix 2), which found foraging photos from 2007 to 2020, yielded two other photos of Black-cheeked Ant-Tanagers consuming food from a stalked, fruiting plant while perched on its stalk; in these photos they were exposed to sunlight, suggesting a degree of habitat openness. Two more photos showed the Black-cheeked Ant-Tanager perched next to another decorative fruit; two additional images showed the Black-cheeked Ant-Tanager consuming other organisms, including an arthropod. The search also yielded photos from 19 observers over 17 dates, ranging from 2007 to 2020 and concentrated from December to June, of Black-cheeked Ant-Tanagers at a feeding station with human-provided fruits, appearing to be bananas or plantains (Appendix 2). This suggests a high degree of reliability in their consumption of fruit, at least during part of the year.
Slud described Black-cheeked Ant-Tanagers as squeezing juice from small fruits as well as being “probably animalivorous” (Slud 1964). Stiles and Skutch went on to describe that the species feeds on melastomes and solanaceous fruits (Stiles and Skutch 1989). Others noted that their diet includes a wide range of animals, including lizards, caterpillars, crickets, grasshoppers, cicadas, and butterflies (Huber et al. 2008). Willis notes that, while visiting army ant swarms, Black-cheeked Ant-Tanagers consumed “spiders, roaches, and long-horned grasshoppers” (Willis 1972).
Range-wide protected area status and connectivity
Black-cheeked Ant-Tanagers showed widespread use of protected areas, with 2544 (76.08%) observations occurring in those areas. Further, about 40% of Black-cheeked Ant-Tanager observations occurred in biological corridors (18.93%, or 633, in Osa Biological Corridor and 22.43%, or 750, in AmistOsa Biological Corridor).
DISCUSSION
Habitat
Our range-wide, multi-datum source assessment suggests that the Black-cheeked Ant-Tanager occurs in secondary forest, riverside forest, and forest edge in addition to old-growth forest. Since nests were found in both forest types as well as in forest edges, it appears even more likely that secondary forest can provide suitable habitat even if closed canopy rainforest of some form may be required (Huber et al. 2008, Jones et al. 2009, Sandoval and Gallo 2009). The presence of nests and foraging observations suggests that secondary forest is used not merely by transient individuals but for substantial portions of the lifecycle.
Importantly, eBird and GIS data analysis support the use of secondary habitat on a range-wide scale because the large majority of observations were based in secondary rainforest. These percentages are likely biased by sampling effort and perhaps proximity to other habitats but nonetheless indicate a clear presence of Black-cheeked Ant-Tanagers in degraded-recovering habitats, particularly because many were stationary observations. The small number of checklists in teak, melina, and oil palm could represent transient individuals or imprecise checklists and suggests that these often pesticide-rich monoculture habitats are likely unsuitable habitat for Black-cheeked Ant-Tanagers.
Consequently, even if other factors still influence distribution of the species (i.e., presence of water, food source availability, interspecific competitive interactions, or land gradient), extensive swaths of secondary forest outside of protected areas may provide habitat for Black-cheeked Ant-Tanagers. These observations highlight the value of forest restoration and regeneration to provide habitat for an endemic species as well as the importance of protection of both mature and secondary forest.
Diet
Foraging observations from the array of sources we analyzed indicate that Black-cheeked Ant-Tanagers are generalists that draw from many food sources, including three families of fruits, numerous invertebrates, and one vertebrate. As such, they should be considered foraging opportunists without a clear specialization. Fruit could comprise an important role in the diet of the species, and the categorization of the Black-cheeked Ant-Tanager as an insectivore is incomplete.
Relationship between habitat and diet
These fruiting plants are characteristic of degraded habitat, so it is possible that Black-cheeked Ant-Tanager habitat use may be tied to the availability of fruiting plants. This possibility could explain the variation in results found by other studies, as fruit availability could depend on any number of other variables, including time of year, temperature, rainfall, and complex synergies of those. It is important to note that these observations included observations from secondary forest.
The documentation of Black-cheeked Ant-Tanagers foraging on decorative plants and feeders suggests that ecolodges, conservation organizations, and landowners could consider planting fleshy fruits that have long stalks, such as Carludovica palmata, to provide a food source that is also visually appealing. This intervention tends to be positively impactful on a population level when food limits reproduction or survival (Williams et al. 2013).
The range of dietary choices for the Black-cheeked Ant-Tanager may suggest a degree of resilience to climate change. In other contexts, dietary specialization has been shown to contribute to climate change vulnerability (Leclerc et al. 2020), and plasticity in foraging behavior may enable the Black-cheeked Ant-Tanager to better persist in its current distribution (Davey et al. 2012, Evans and Moustakas 2018). Theory predicts that generalists will be better able to colonize new locations, even though its explanatory power for conservation and management may be limited (Angert et al. 2011).
Additionally, because the species consumes some insects, it may be at risk from insecticide use, common in Costa Rican agriculture (Galt 2008, Brown et al. 2020). This threat is of particular concern because Black-cheeked Ant-Tanagers were at times reported to eBird in proximity to monocultures, and herbicide and pesticide use can lead to bioaccumulation and food resource loss (Sánchez-Bayo 2011). Future studies could assess the extent of this threat in the Black-cheeked Ant-Tanager range.
Range-wide protected area status, population connectivity, and distribution
A species’ ability to move on an elevational gradient is an important consideration in devising climate-resilient conservation strategies (Forero-Medina et al. 2011, Elsen et al. 2018, Freeman et al. 2018). In addition to its diet, the Black-cheeked Ant-Tanager’s use of both mature and secondary forest may facilitate an added degree of resilience to climate change, because swaths of secondary forest may serve as corridors along an elevational gradient. Many Black-cheeked Ant-Tanager sightings already occurred in biological corridors. Ensuring the corridors’ protection and restoration (where degraded) could be vital to Black-cheeked Ant-Tanager survival in a changing climate. Indeed, much of the connectivity sought from Costa Rica’s system of biological corridors is and will continue to be derived from secondary forest.
Given Black-cheeked Ant-Tanager use of secondary forest, forest restoration may also be a powerful tool to create corridors for the species. Secondary forest regeneration in the Osa has been especially effective on former grassland (Furey et al., https://doi.org/10.6084/m9.figshare.19337912.v1). Thus, habitat regeneration efforts resulting from the implementation of the Amistosa Biological Corridor could be impactful. The Black-cheeked Ant-Tanager stands to benefit from forest restoration that creates a large-scale corridor, connecting and restoring forests across elevations and thereby ensuring the peninsula’s ecosystems, in particular its endemic species, are resilient to climate change.
In addition to corridors, protected areas may be important reservoirs for the species. The prevalence of Black-cheeked Ant-Tanager community science observations from protected areas is not necessarily representative of their proportional distribution because of observer bias, but it suggests that Black-cheeked Ant-Tanagers occur in those protected areas. As such, their protection should be continued and enhanced.
The eBird data yielded other insights into the distribution of the Black-cheeked Ant-Tanager (Fig. 1). Presence of Black-cheeked Ant-Tanagers on the north end of the Golfo Dulce (between the two main strongholds) is a favorable indicator of this connectivity, although more research is needed on Black-cheeked Ant-Tanager use of that region. The Golfo Dulce Forest Reserve connects the two populations, so strengthened conservation measures, including reforestation, should be prioritized there to ensure the species’ survival. The presence of Black-cheeked Ant-Tanager eBird locations far to the south and west of the population near Golfito, as well as on the northeastern rim of its range, could indicate strong capabilities for persistence in parts of its historic range (Fig. 1). Should enough closed-canopy forest exist to connect the populations, these observations are promising for the possibility of connectivity with the higher elevation Fila de Costeña (which reaches 1000–1500 m, as opposed to the maximum elevation of roughly 745 m on the Osa; Fisher et al. 2004, Cornejo et al. 2012). Increasing secondary forest cover on this margin of its range, which has been heavily impacted by land conversion to grassland and oil palm, is therefore a priority. This paper therefore presents another example of how citizen science can inform our understanding of species distribution (Yu et al. 2010).
CONCLUSION
As the biodiversity crisis worsens and climate change and deforestation cause shifts in bird populations, advancing the understanding of habitat requirements and ecology for endemic species is paramount. By synthesizing field observations, community science, and historical observations, we applied a range-wide, multi-datum approach to study the ecological needs of the Black-cheeked Ant-Tanager.
Leveraging these data sources, we determined that the Black-cheeked Ant-Tanager uses secondary forest, on the basis of occurrence data and supported by nesting and foraging observations. The use of secondary forest suggests that secondary forest habitats could enable connectivity between population strongholds. The Black-cheeked Ant-Tanager occurs in key habitat corridors on the Osa Peninsula, pointing to the importance of preserving existing forest and promoting secondary forest restoration in these corridors to facilitate the species’ connectivity in a changing climate.
This study provides an improved basis for a corridor-centered conservation strategy under climate change and provides a model for targeted climate adaptation strategies, which could also provide habitat for many other species.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
We thank Osa Conservation and their supportive staff for hosting this research at the Osa Conservation Campus, Ruthmery Pillco Huarcaya for kindly identifying plants that served as Black-cheeked Ant-Tanager food sources, Thomas Meinzen for generously contributing Black-cheeked Ant-Tanager observations, Eleanor Flatt for her valuable guidance, and María Andreína Pacheco, Max Villalobos, and Pascale Poussart for generous advice and encouragement. We thank the American Birding Association and members of the Washington, D.C., birding community for their financial support. We thank two anonymous reviewers and, in particular, editor Mark Hauber for constructive feedback that made the paper substantially stronger. The Office of Undergraduate Research at Princeton University contributed funding toward the publication fee. I dedicate this paper to Jonathan Franzen for his efforts to support young people engaging in avian conservation and field biology.
DATA AVAILABILITY
The data that support the findings of this study are openly available in figshare at https://doi.org/10.6084/m9.figshare.17912324.
LITERATURE CITED
Angert, A. L., L. G. Crozier, L. J. Rissler, S. E. Gilman, J. J. Tewksbury, and A. J. Chunco. 2011. Do species’ traits predict recent shifts at expanding range edges? Ecology Letters 14(7):677-689. https://doi.org/10.1111/j.1461-0248.2011.01620.x
Brown, J., T. Flint, and J. LaMay. 2020. The politics of pineapple: examining the inequitable impacts of southern Costa Rica’s pineapple industry. Journal of Public and International Affairs. https://jpia.princeton.edu/news/politics-pineapple-examining-inequitable-impacts-southern-costa-ricas-pineapple-industry
Ceballos, G., P. R. Ehrlich, and P. H. Raven. 2020. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proceedings of the National Academy of Sciences 117(24):13596-13602. https://doi.org/10.1073/pnas.1922686117
Cornejo, X., S. A. Mori, R. Aguilar, H. Stevens, and F. Douwes. 2012. Phytogeography of the trees of the Osa Peninsula, Costa Rica. Brittonia 64:76-101. https://doi.org/10.1007/s12228-011-9194-0
Cornils, J. S., I. Riedl, J. Fricke, M. Katz, and C. H. Schulze. 2015. Population density and habitat preferences of the Black-cheeked Ant-tanager Habia atrimaxillaris. Bird Conservation International 25(3):306-321. https://doi.org/10.1017/S0959270914000215
Davey, C. M., D. E. Chamberlain, S. E. Newson, D. G. Noble, and A. Johnston. 2012. Rise of the generalists: evidence for climate driven homogenization in avian communities. Global Ecology and Biogeography 21:568-578. https://doi.org/10.1111/j.1466-8238.2011.00693.x
Elsen, P. R., W. B. Monahan, and A. M. Merenlender. 2018. Global patterns of protection of elevational gradients in mountain ranges. Proceedings of the National Academy of Sciences 115(23):6004-6009. https://doi.org/10.1073/pnas.1720141115
Evans, M. R., and A. Moustakas. 2018. Plasticity in foraging behaviour as a possible response to climate change. Ecological Informatics 47:61-66. https://doi.org/10.1016/j.ecoinf.2017.08.001
Fisher, D. M., T. W. Gardner, P. B. Sak, J. D. Sanchez, K. Murphy, and P. Vannucchi. 2004. Active thrusting in the inner forearc of an erosive convergent margin, Pacific coast, Costa Rica. Tectonics 23(2):TC2007. https://doi.org/10.1029/2002TC001464
Forero-Medina, G., J. Terborgh, S. J. Socolar, and S. L. Pimm. 2011. Elevational ranges of birds on a tropical montane gradient lag behind warming temperatures. PLoS ONE 6(12):e28535. https://doi.org/10.1371/journal.pone.0028535
Freeman, B. G., and A. M. Class Freeman. 2014. Rapid upslope shifts in New Guinean birds illustrate strong distributional responses of tropical montane species to global warming. Proceedings of the National Academy of Sciences 111(12):4490-4494. https://doi.org/10.1073/pnas.1318190111
Freeman, B. G., M. N. Scholer, V. Ruiz-Gutierrez, and J. W. Fitzpatrick. 2018. Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. Proceedings of the National Academy of Sciences 115(47):11982-11987. https://doi.org/10.1073/pnas.1804224115
Galt, R. E. 2008. Pesticides in export and domestic agriculture: reconsidering market orientation and pesticide use in Costa Rica. Geoforum 39(3):1378-1392. https://doi.org/10.1016/j.geoforum.2007.12.003
Gillies, C. S., and C. C. St. Clair. 2008. Riparian corridors enhance movement of a forest specialist bird in fragmented tropical forest. Proceedings of the National Academy of Sciences 105(50):19774-19779. https://doi.org/10.1073/pnas.0803530105
Guillermo, J. B. 2017. Mapa de corredores biológicos. Ministerio del Ambiente y Energía, Sistema Nacional de Áreas de Conservación, Costa Rica. https://www.sinac.go.cr/EN-US/correbiolo/Pages/default.aspx
Harris, G., and S. L. Pimm. 2008. Range size and extinction risk in forest birds. Conservation Biology 22(1):163-171. https://doi.org/10.1111/j.1523-1739.2007.00798.x
Huber, W., A. Weissenhofer, and G. Aubrecht. 2008. First observations of nest and nestling of the Black-cheeked Ant-Tanager Habia atrimaxillaris (Dwight & Griscom 1924), endemic to the Golfo Dulce rainforests, Costa Rica. Brenesia 70:53-56.
Jones, E., A. Gallo, and D. Lebbin. 2009. Ecology and distribution of endemic birds of the Osa Peninsula. Evergreen Foundation and Friends of the Osa, Puerto Jiménez, Costa Rica.
Leavelle, K. M. 2013. Avian inventory and monitoring report for Osa conservation properties at Cerro Osa and Piro Research Stations, Osa Peninsula, Costa Rica. Osa Conservation, Washington, D.C., USA.
Leclerc, C., F. Courchamp, and C. Bellard. 2020. Future climate change vulnerability of endemic island mammals. Nature Communications 11:4943. https://doi.org/10.1038/s41467-020-18740-x
Lobo, J., R. Aguilar, E. Chacón, and E. Fuchs. 2008. Phenology of tree species of the Osa Peninsula and Golfo Dulce region, Costa Rica. Stapfia 88:547-555.
Rocha, O. J., B. Vílchez, and A. L. Araya Anchetta. 2006. A mast fruiting episode of the tropical tree Peltogyne purpurea (Caesalpinaceae) in the Osa Peninsula, Costa Rica. Revista de Biología Tropical 54(4):1151-1155.
Sanchez-Azofeifa, G. A., B. Rivard, J. Calvo, and I. Moorthy. 2002. Dynamics of tropical deforestation around national parks: remote sensing of forest change on the Osa Peninsula of Costa Rica. Mountain Research and Development 22(4):352-358. https://doi.org/10.1659/0276-4741(2002)022[0352:DOTDAN]2.0.CO;2
Sánchez-Bayo, F. 2011. Impacts of agricultural pesticides on terrestrial ecosystems. Pages 63-87 in F. Sánchez-Bayo, P. J. van den Brink, and R. M. Mann, editors. Ecological impacts of toxic chemicals. Bentham Science Publishers, United Arab Emirates.
Sandoval, L., and A. Gallo. 2009. Description of the nest and eggs of the Black-cheeked Ant Tanager (Habia atrimaxillaris). Wilson Journal of Ornithology 121(3):635-637. https://doi.org/10.1676/08-130.1
Sandoval, L., C.-O. Morales, J.-D. Ramírez-Fernández, P. Hanson, L.-R. Murillo-Hiller, and G. Barrantes. 2019. The forgotten habitats in conservation: early successional vegetation. Revista de Biología Tropical 67(S2):S36-S52. https://doi.org/10.15517/rbt.v67i2SUPL.37202
Schulze, C. H., P. Leidinger, B. Paces, and A. F. Reyes Páez. 2019. The importance of reforested and naturally regenerating young forest patches as secondary habitats for forest birds in the Biological Corridor La Gamba, Costa Rica. Acta ZooBot Austria 156:79-98.
Shrestha, S., M. Bouffard, J. Langstaff, C. Lee, E. Pauline, and S. Tingle. 2018. Assessing threats to river water quality and mangrove health based on watershed land use on the Osa Peninsula, Costa Rica. AGU Fall Meeting Abstracts: GC11H-1000.
Sistema Nacional de Áreas de Conservación. 2018. Plan estratégico 2018-2025 del programa nacional de corredores biológicos de Costa Rica (informe final). Programa Nacional de Corredores Biológicos: San José, Costa Rica. https://biocorredores.org/biodiver-city-sanjose/sites/default/files/docs/PLAN%20ESTRAT%C3%89GICO%202018%20%E2%80%93%202025%20Programa%20Nacional%20de%20Corredores%20%20Biol%C3%B3gicos%20de%20Costa%20Rica2%20%20.pdf
Slud, P. 1964. The birds of Costa Rica: distribution and ecology. Bulletin of the American Museum of Natural History. Volume 128. New York, New York, USA.
Stiles, G., and A. Skutch. 1989. A guide to the birds of Costa Rica. Comstock Publishing Associates, Ithaca, New York, USA.
Villalobos, M., and M. Ramírez. [Date unknown.] Trail system. Map at Piro Biological Station. Osa Conservation, Puerto Jiménez, Costa Rica.
Whitworth, A., C. Beirne, E. Flatt, G. Froese, G. Nuñez, and A. Forsyth. 2021. Recovery of dung beetle biodiversity and traits in a regenerating rainforest: a case study from Costa Rica’s Osa Peninsula. Insect Conservation and Diversity 14(4):439-454. https://doi.org/10.1111/icad.12470
Williams, D. R., R. G. Pople, D. A. Showler, L. V. Dicks, M. F. Child, E. K. H. J. zu Ermgassen, and W. J. Sutherland. 2013. Bird conservation: global evidence for the effects of interventions. Pelagic Publishing, Exeter, UK.
Willis, E. O. 1972. Taxonomy, ecology, and behavior of the Sooty Ant-Tanager (Habia gutturalis) and other Ant-Tanagers (Aves). American Museum Novitates: no. 2480. American Museum of Natural History, New York, New York, USA.
Wilson, S., D. M. Collister, and A. G. Wilson. 2011. Community composition and annual survival of lowland tropical forest birds on the Osa Peninsula, Costa Rica. Ornitología Neotropical 22:421-436.
Yu, J., W.-K. Wong, and R. A. Hutchinson. Edited by G. I. Webb, B. Liu, C. Zhang, D. Gunopulos, and X. Wu. 2010. Modeling experts and novices in citizen science data for species distribution modeling. Presented 13-17 December. 2010 IEEE International Conference on Data Mining. Pages 1157-1162. Institute of Electronic and Electronical Engineers, Sydney, New South Wales, Australia. https://doi.org/10.1109/ICDM.2010.103
Fig. 1
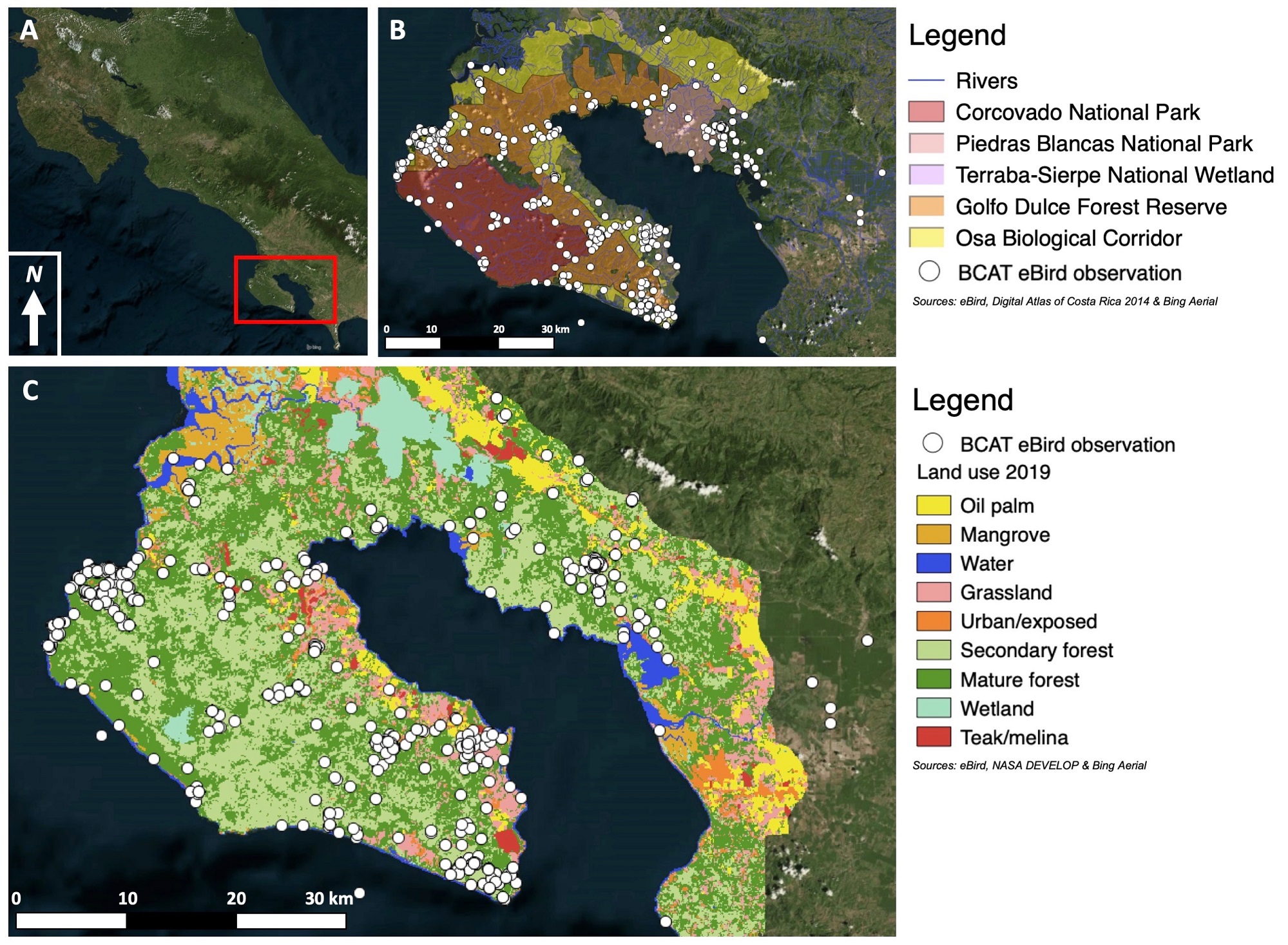
Fig. 1. (A) Location of the study region, Osa Conservation Area (ACOSA), in Costa Rica. (B) eBird Black-cheeked Ant-Tanager observations in relation to key protected areas, corridors, and rivers. (C) eBird observations and land use classifications.