







The following is the established format for referencing this article:
Israel, A. M., S. Hayes, B. P. Boyd, and B. J. M. Stutchbury. 2023. Effects of nest concealment on nest predation and cowbird parasitism, flight initiation distance and female stress levels in the Wood Thrush. Journal of Field Ornithology 94(1):9.ABSTRACT
Temperate zone songbirds in North America can experience high levels of nest predation and Brown-headed Cowbird (Molothrus ater) parasitism, which may contribute to population declines, and an important question is whether nesting females can mitigate these threats through greater nest concealment. However, there is little known about the pros and cons of nest concealment to the incubating female, and whether concealment influences female corticosterone levels or nest escape behavior to potentially reduce risk of predation on females. This study investigates whether nest concealment resulted in reduced risk of nest predation and Brown-headed Cowbird parasitism in the Wood Thrush (Hylocichla mustelina), a forest bird that is declining across most of its range. We monitored nests (n = 186) in a highly fragmented forest landscape in southwestern Ontario, Canada and found high nest predation (53.2%) and moderate cowbird parasitism (25.3%). Overall nest concealment, nest height, and forest fragment size were not good predictors of daily nest survival or cowbird parasitism, but greater nest concealment one meter above the nest strongly reduced nest predation risk. We speculate that because most Wood Thrush nests are relatively low (< 2.5 m above ground), above-nest concealment may be effective against detection by avian nest predators. Overall nest concealment was not negatively correlated with flight initiation distance (FID) or positively related to baseline corticosterone level. This suggests that high nest concealment does not increase stress or personal risk to incubating females; however, nest concealment manipulations are needed to further explore whether any such trade-offs exist.
RESUMEN
Las aves Passeriformes de las zonas templadas en Norte América pueden experimentar altos niveles de depredación de nidos y parasitismo por parte de Molothrus ater, lo cual puede contribuir a la disminución de las poblaciones. Una pregunta importante es determinar si las hembras que anidan pueden mitigar las amenazas a través del ocultamiento de sus nidos. Sin embargo, poco es conocido sobre las ventajas y desventajas del ocultamiento de los nidos sobre la hembra que incuba y si el ocultamiento influye en los niveles de corticosterona de las hembras o el comportamiento de escape del nido lo que potencialmente disminuye el riesgo de depredación sobre las hembras. Este estudio investiga si el ocultamiento del nido resulta en un menor riesgo de depredación de los nidos y el parasitismo por parte de Molothrus ater en Hylocichla mustelina, una especie de bosque cuyas poblaciones están disminuyendo a través de todo su rango de distribución. Monitoreamos nidos (n=186) en un paisaje de bosque altamente fragmentado en el suroeste de Ontario, Canadá y encontramos una alta depredación de nidos (53.2%) y un parasitismo moderado por parte de Molothrus ater (25.3%). En general el ocultamiento de nidos, la altura de los nidos y el tamaño del fragmento del bosque no fueron buenos predictores de la tasa diaria de supervivencia o del parasitismo, pero un mayor ocultamiento un metro por encima del nido disminuyó significativamente el riesgo de depredación. Especulamos que debido a que la mayoría de los nidos de Hylocichla mustelina están relativamente bajos (< 2.5 m sobre el suelo), el ocultamiento por encima del nido puede ser efectivo en contra de la detección de los nidos por parte los depredadores aviares. En general, el ocultamiento del nido no estuvo negativamente correlacionado con la distancia del vuelo de iniciación (FID) ni positivamente relacionado con los niveles de línea base de corticosterona. Esto sugiere que un alto ocultamiento del nido no incrementa el estrés o el riesgo personal a las hembras incubantes; sin embargo, manipulaciones del ocultamiento de los nidos son necesarias para explorar en el futuro si existe alguno de estos compromisos.
INTRODUCTION
Predation is a ubiquitous selective pressure, and animals have evolved a wide variety of behavioral (e.g., hiding, fleeing), morphological (e.g., spines, coloration), physiological (e.g., toxins), and sensory defensive adaptations (Kats and Dill 1998, Langerhans 2007). For passerine birds, nest predation rates exceed 50% for most species (Robinson et al. 2000) and predation causes the large majority of all nest failures (Ibáñez-Álamo et al. 2015). Avian strategies to reduce nest predation include direct defense of the nest by the parents (Montgomerie and Weatherhead 1988, Thys et al. 2019) and/or indirect defense through concealment of the nest (Weidinger 2002, Borgmann and Conway 2015). To reduce the risk of nest predation, birds could choose nesting sites in areas with a low predation risk and/or attempt to conceal their nest with dense vegetation to avoid detection by visually searching predators (Martin 1993). Nest predation risk, and how this can be mitigated behaviorally, is evolutionarily important for understanding life history trade-offs (Martin 2014, Ibáñez-Álamo et al. 2015) and plays an important role in population declines of birds in disturbed landscapes (Faaborg et al. 2010). In this study, we test whether greater nest concealment is associated with reduced risk of nest predation or cowbird parasitism for the Wood Thrush (Hylocichla mustelina), and whether high nest concealment affects a female’s flight initiation distance or corticosterone levels.
The nest-concealment hypothesis (Martin 1993, Borgmann and Conway 2015) predicts: (1) songbirds should choose nesting sites with more dense foliage rather than random sites and (2) the probability of nest predation and parasitism should decrease with increased foliage density at the nest. Some studies have found support for the nest-concealment hypothesis but across more than 100 studies, support has been equivocal (Borgmann and Conway 2015). For instance, Canada Warblers (Wilsonia canadensis) choose nesting sites with significantly greater concealment and vegetation density when compared to randomly selected sites within the birds’ territory, and successful nests had greater nest concealment than those that were depredated (Goodnow and Reitsma 2011). Greater foliage density at Hermit Thrush (Catharus guttatus) nests was also associated with reduced nest predation (Martin and Roper 1988). However, many studies have found no effect of nest concealment on predation risk (e.g., Holway 1991, Burhans and Thompson 1998, Wilson and Cooper 1998). Nest concealment may be less effective as a predator defense when males are brightly colored, but more effective for species with bright female coloration (Borgmann and Conway 2015), neither of which apply to Wood Thrushes. Experimental studies that manipulate concealment have found that low nest concealment does not significantly increase nest predation (Howlett and Stutchbury 1996, Peak 2003, Li et al. 2018) perhaps because many nest predators do not hunt visually.
Previous studies of Wood Thrush have found mixed results for whether vegetation density near nests (e.g., concealment) predicts nesting success. In a large contiguous forest, Farnsworth and Simons (1999) found that percentage vegetation cover directly above the nest was significantly positively correlated with nesting success. In a small suburban woodlot, Johnson (1997) found that higher overall percentage concealment of nests reduced nest predation rate but in a highly fragmented landscape Newell and Kostalos (2007) found overall nest concealment did not predict predation rate. Such variation among studies of the same species could result from geographical and habitat-related differences in the abundance and types of nest predators.
Although most tests of the nest-concealment hypothesis focus on traditional nest predators (e.g., mammals, snakes, corvids) the brood parasitic Brown-headed Cowbird (Molothrus ater) is also a nest predator. Cowbirds often remove host eggs during the egg-laying period, and they have also been documented removing eggs during incubation (Benson et al. 2010, Friesen et al. 2013) and, less frequently, removing host young (Hoover and Robinson 2007, Conkling et al. 2012). In addition to reducing nest predation from cowbirds, hosts may benefit from concealing nests from cowbirds to reduce the fitness costs of raising cowbird young. The high demands on host parental care can reduce reproductive success of parasitized nests by 50% or more (Lorenzana and Sealy 1999). Cowbird parasitism can also increase the risk of nest predation (Hannon et al. 2009, Latif et al. 2012) through mechanisms such as increased detectability to predators because of loud and frequent begging calls by cowbird chicks and increased parental feeding rates to parasitized nests. Evidence for nest concealment reducing cowbird parasitism risk is also mixed, with some studies finding support (Saunders et al. 2003, Sharp and Kus 2006, Hackemack et al. 2016) while others do not (Burhans and Thompson 1998, Wilson and Cooper 1998, Banks and Martin 2000).
Although the nest concealment hypothesis expects females to conceal their nests as much as possible, in most species there is wide variation among females in extent of nest concealment. Why do many females not do a better job of hiding their nests? Nest site selection may involve a trade-off between concealing the nest from predators to increase survival of eggs/young versus having a more open nest site to increase a female’s ability to escape from predators (Götmark et al. 1995). This assumes that a highly concealed nest reduces a female’s ability to detect and monitor an approaching predator, and so puts the female’s own life at risk. This trade-off is reflected in a female’s decision to remain on the nest as a predator approaches so that she does not draw attention to the nest versus the need for her to flee at some point to protect herself. Several studies on ground-nesting waterfowl, shorebirds, and grouse have shown that high nest concealment reduces the distance at which a female flushes from the nest in response to a predator (the flight initiation distance, FID; Albrecht and Klvaňa 2004, Seltmann et al. 2013), and that high nest concealment increases risk of predation for incubating females (Wiebe and Martin 1998, Amat and Masero 2004, Miller et al. 2007, Öst and Steele 2010).
Another potential cost to females of choosing a highly concealed nest site could be the physiological costs of elevated stress hormones, specifically corticosterone (CORT). During the nesting attempt, increased predation risk for females with concealed nests could increase a female’s CORT levels, and this could have longer-term negative effects on individuals, such as delayed breeding (Schoech et al. 2009) and reduced reproductive success (Bonier et al. 2009). An alternative hypothesis is that short FID of females (i.e., boldness) will be associated with low CORT because this physiological mechanism allows individuals to better tolerate high risk environments such as urban areas (Atwell et al. 2012). Although a positive relationship between FID and CORT has been shown in comparisons between populations (e.g., rural versus urban; Atwell et al. 2012), it is not known to what extent this also applies to within-population variation among females, or whether FID and CORT are linked to nest concealment. In Eider ducks (Somateria mollissima) females that had a bold phenotype (short FID, low stress responsiveness) were more likely to select nest sites in which risk of predation on females was higher, but nest predation risk was lower (high concealment or farther from shore; Seltmann et al. 2013).
Little is known about the relationship between nest concealment, FID, and corticosterone in passerines. Incubating female Brown Thornbills (Acanthiza pusilla), which build closed dome nests, are vulnerable to predation if they become trapped in their nest. During and after experimental playback of predator calls, females spent more time looking out of their nest if it was highly concealed in dense vegetation, which suggests such females have a higher perceived risk of predation (Schneider and Griesser 2013). In the open-cup nesting Song Thrush (Turdus philomelos), flight initiation distance was shorter for females with highly concealed nests, but CORT was not measured (Götmark et al. 1995). At the population level, Song Sparrows (Melospiza melodia) that experienced experimentally reduced nest predation risk had lower baseline levels of CORT, but nest concealment was not examined (Clinchy et al. 2004).
We tested if the distance at which female Wood Thrush flush from the nest as a predator approaches is negatively correlated with nest concealment. We also tested if short FID or high nest concealment is associated with higher baseline or stress-induced corticosterone levels in incubating female Wood Thrush.
METHODS
Study species and study sites
The Wood Thrush is a long-distance migrant that has declined severely in the last 50 years (-1.3%/year with cumulative loss ~ 50%; Sauer et al. 2019). Wood Thrush often nest in low branches of the shrub layer, less than 2.5 m from the ground, making their nests easily accessible for monitoring (Evans et al. 2020). They nest in a variety of habitats ranging from mature forest interior to scrubby forest edge and readily occupy small forest fragments where they may experience high nest predation (Hoover et al. 1995). Wood Thrush are double brooded, and females will often re-nest after a nest failure.
In 2017, 2018, and 2019, Wood Thrush nests (n = 186) were found within forest fragments throughout Norfolk County, southwestern Ontario, Canada, a region with 17% forest cover located on the north shore of Lake Erie (Eng et al. 2011). Forest fragments were selected based on fragment size and distance between study sites for logistical purposes. Forest fragments were categorized as small (11-69 ha; n = 10) versus large (162-500 ha; n = 10) and consisted of deciduous and mixed forest. Within these fragments, forest cover nearly complete was interrupted only by walking trails, small streams, and ravines.
Video monitoring of Wood Thrush nest predation events in forest fragments < 100 km from our study site (Friesen et al. 2013) found that the 5 most frequent nest predators during the incubation or nestling periods were Brown-headed Cowbird (26.7%), Raccoon (Procyon lotor; 17.8%), Cooper’s Hawk (Accipiter cooperii; 16.1%), American Crow (Corvus brachyrynchos; 8.9%), and Blue Jay (Cyanocitta cristata; 7.1%). Other culprits included Red Squirrel (Tamiasciurus hudsonicus), Eastern Chipmunk (Tamias striatus), Eastern Gray Squirrel (Sciurus carolinensis), Red-tailed Hawk (Buteo jamaicensis), Sharp-shinned Hawk (Accipiter striatus), and Short-tailed Weasel (Mustela erminea).
Field methods
Nests were located May-Aug by: (1) finding singing males within the study sites and then intensively searching likely areas within their territories, (2) by finding females carrying nesting material and following them to their nesting location, or (3) searching typical nesting habitats. Female Wood Thrush incubate their eggs for about 13-14 days, and young will begin to fledge around 12 days after hatching (Evans et al. 2020). Nests were checked on average every eight days to minimize the possibility that human visits would attract predators, yet still provide sufficient estimates of nest survival probability. The final check occurred when nestlings were ~10 days old, to avoid causing premature fledging, which is likely to reduce fledgling survival. To check the nests as quickly and in the least invasive way possible, a cell phone with video recording capabilities was attached to a long stick and raised over top of each nest, while standing as far back from the nest as possible. The video was reviewed away from the nest and allowed us to determine nest contents, including cowbird eggs or nestlings. Indicators of predation included eggshells, feathers, or an empty nest that had previously contained eggs or nestlings. A nest was considered successful if at least one nestling survived to be 10 days old. For successful nests, the ~ 10-day-old nestlings were banded with aluminum Canadian Wildlife Service bands, and for many nests, one nestling was fitted a radio tag for a concurrent study on fledgling survival (Hayes, unpublished manuscript).
The concealment of a nest was measured at least one week after the nestlings fledged to avoid disturbance to the incubating/feeding parents and to fledglings who often remain in the nest vicinity for several days until they become more mobile. Because 75% of nests fledged after mid-June, when leaf growth rate has peaked, we assume that the change in nest concealment during the nest development period before we measured concealment was small compared to variation in concealment among nests. Concealment was measured at three different heights (ground level, nest height, and one meter above nest height) by taking digital photo(s) of the nest from one meter away at each of the four primary compass directions around the nest. For ground level, one additional photo was taken from the ground, standing immediately under the nest, and for one meter above the nest, one additional photo was taken from one meter distance immediately above the nest. For each image, the percentage of the nest that was concealed to the nearest 10% was estimated by one person (A.I.) for consistency. The overall concealment of each nest was calculated as the average of the 14 estimates of concealment (similar to Howlett and Stutchbury 1996). Concealment from one meter above was the average of five estimates, concealment at nest height the average of four estimates, and concealment at ground level the average of five estimates. Nests that were > 5 m above ground (4% of all nests located) were too high to measure all the concealment variables and so were excluded.
Flight initiation distance measurements were taken in 2018 and 2019 during regular nest checks, so as not to cause female Wood Thrush to flush from their nests more than necessary. If a female Wood Thrush was seen sitting on her nest during the incubation period, the nest was approached from a random direction at a steady walking pace and in a straight line. The point at which the female left her nest was the FID measurement and was recorded in meters, with observations beginning from a starting point about 20 m away.
Blood sampling for corticosterone (CORT) analysis occurred in 2018 and 2019 on 36 adult female Wood Thrush midway through their incubation stage; a different study found that CORT does not vary with nest stage in female Wood Thrush (Done et al. 2011). Blood-sampled females were radio-tagged as part of a concurrent study on the effects of habitat quality on female body condition and migration timing (Boyd 2022). All samples were collected at least three hours after sunrise and three hours before sunset. Adult females were caught by placing mist nets 5-10 m from nests; most females remained on nests during net set up. To obtain blood samples within three minutes of capture, as an estimate of baseline conditions (Romero and Reed 2005), incubating females were flushed toward the nets by slowly walking toward the nest. Most females (61%) were captured immediately, but those that were not we flushed a second time after they had returned to the nest to resume incubating. If females alarm called or were visually agitated at any time, the mist nets were dismantled, and capture was attempted on a different day.
Each bird underwent a capture-stress protocol that involved taking an initial blood sample within 3 min. of capture, holding the bird for 30 min. in a fabric bag, and then taking a second blood sample (Wingfield et al. 1994). Approximately 100 ul of blood was taken from the brachial vein of one wing; the vein was punctured with a 26 g needle and capillary tubes were used to collect the blood. Blood samples were immediately transferred into 400 ul Eppendorf tubes and stored on ice. The bird was then banded, measured, radio-tagged, and then placed in a cloth bag until 30 min from capture had elapsed. A second blood sample was then taken from the other wing to measure the induced stress response of the bird. Samples were centrifuged within eight hours and blood plasma was separated and stored in a freezer.
Corticosterone concentration was measured using the commercially available Enzo Life Sciences Corticosterone ELISA kit (ADI-900-097) using the protocol developed by Wada et al. (2007), including the process of validating and optimizing the kit using Wood Thrush plasma. Samples were run in duplicate and if both assays yielded a concentration that fell outside the standard curve, the sample was excluded from further analysis (n = 1). The intra-assay coefficient of variation was 9.3% for samples assayed in 2018, and 11.5% for samples assayed in 2019. The inter-assay coefficient of variation based on a standard concentration run in duplicate on each plate was 8.7% in 2018 and 6.4% in 2019.
Statistical and survival analysis
The nest concealment data were not normally distributed; therefore, the differences in nest concealment among successful/unsuccessful and non-parasitized/parasitized nests were calculated using non-parametric statistical tests (Mann-Whitney U-test and generalized linear models, GLMs). All statistical tests and figures were generated in RStudio (Version 1.1.456).
Daily nest survival rate (DSR) was analyzed using the R (R Core Team 2020) interface to run the nest survival model in the program MARK 6.0 (White and Burnham 1999) with the package RMARK version 2.2.7 (Laake 2013). To run this model, the following was needed for each nest: the day the nest was found, the last day the nest was checked and still alive, the last day the nest was checked, the fate of the nest (predated or successful), and the number of nests that had the same encounter history. In addition, nest age was included to test for effects on nest survival (Rotella et al. 2004).
A parallel study (Hayes, unpublished manuscript) on Wood Thrush found that nest age was overwhelmingly the best predictor for temporal sources of variation in nest predation. Therefore, models in this study were run using nest age as the base model to which the predictors were added. Additive and interactive models were run with each of the predictor variables of interest that included nest concealment (overall, ground level, nest level, and one meter above) and nest height. Forest fragment size (small vs large) and year were also included as categorical variables in the model set. Model support was determined using the Akaike Information Criterion corrected for small sample size (AICc) and cumulative model weights (Arnold 2010).
RESULTS
Nest concealment and predation/parasitism
Of all nests found, 53.2% (99 of 186) failed due to nest predation and 25.3% (47 of 186) were parasitized by cowbirds. Overall nest concealment (% nest not visible) varied widely among females, from < 5% to almost 80%, with an average 29.4% (Fig. 1A). Concealment from ground level was positively correlated with both nest level concealment (Spearman rank correlation, rs = 0.32, P < 0.001) and concealment from one meter above the nest (rs = 0.17, P = 0.02).
Of the 186 nests monitored, there were 141 nests with the complete information needed (e.g., first egg date, nest height, cowbird parasitism) to be included in the nest survival model. RMARK analysis showed that the model including nest age and nest concealment from one meter above the nest [S(NestAge + ConcealAbove)] was by far the best fitting model (Wi = 0.744; Table 1, Table 2). Daily nest survival rate increased strongly as concealment from one meter above the nest increased (Fig. 2), and nest survival was lower for older nests (e.g., those with nestlings). Nest concealment one meter above the nest ranged from 0-98% and averaged 41.5%. The second-best model [S(NestAge)] had a ΔAIC = 4.74 and a weighting of only 0.072 but this base model fit far better than the constant model S(1), which had ΔAIC = 22.70. Other nest concealment variables (e.g., nest level, ground level, overall) and other parameters (nest height, parasitism by cowbirds, fragment size, year) were poor predictors of nest survival (Table 1) compared with concealment one meter above the nest.
For cowbird parasitism, there was no strong pattern of different concealment between non-parasitized and parasitized nests (Fig. 1B). A binomial GLM to predict cowbird parasitism including overall concealment, concealment one meter above the nest, nest height, fragment size, and year found the model was not significant (Table 3). Fragment size was a significant predictor (p = 0.047) with nests in large forest fragments experiencing lower risk of parasitism.
We also tested if nest concealment increased over the nesting season, which could result from increasing leaf cover over time and/or experienced females choosing more concealed sites when renesting. Leaf growth rate of shrubs/trees in the study area is highest in late May and early June. Overall nest concealment increased significantly over the nesting season (gamma generalized linear model, t = 3.57, n = 142 nests, P < 0.001), but the effect size was modest. Over the 40-day period from 20 May to 30 June, overall nest concealment increased from an average of 25% to only 35%. During the peak time that first nests were being laid (20 May) nests ranged widely in concealment (0-75%), and at the end of June there was still a wide range of nest concealment (10-78%). Throughout the season, many females choose sites with low nest concealment. We could not test for changes in nest concealment over time for individual females because most nesting females had not been banded.
Concealment, corticosterone, and flight initiation distance (FID)
There was no relationship found between overall nest concealment and either initial CORT (within 3 minutes of capture) or response CORT (30 minutes after capture; Fig. 3). As expected, initial CORT concentrations were much lower than response CORT. A gamma GLM including overall concealment, concealment one meter above the nest, nest height, fragment size, and year for initial CORT and response CORT found the models were not significant (Table 4).
If highly concealed nests compromise a female’s ability to detect an approaching predator, then there should be a negative relationship between FID and concealment. Unexpectedly, the majority (24 of 30; 80%) of FID measurements were 0 m in 2018 (Fig. 4A) meaning the bird did not flush until the person was at the nest. Flight initiation distance measurements were more variable in 2019 and only 2 of 17 females (12%) had an FID of 0 m (Fig 4B). Nest concealment was not a strong predictor of FID (Fig. 4; n = 47).
To assess the roles of both concealment and CORT in predicting FID, we used a subset of nests (n = 14) for which we had measured all variables for a given female. The GLM analysis including nest concealment measures, FID, initial CORT, 30-min CORT, nest height, fragment size, and year revealed that only nest height was a significant predictor of FID (p = 0.001; Table 5). Females with higher nests exhibited shorter FID distances (i.e., allowed a terrestrial predator to approach more closely to the base of the nesting tree/shrub before flushing). Contrary to prediction, there was no relationship between FID and either initial CORT or response CORT concentration (Fig. 5, Table 5).
DISCUSSION
Nest concealment
In this study, Wood Thrush had high nest predation (53%), which was comparable to nest predation rates (52-62%) in fragmented landscapes for other studies of this species (Hoover et al. 1995, Driscoll et al. 2005, Etterson et al. 2014). The level of cowbird parasitism on Wood Thrush was moderate (25%) compared with other studies of this species in fragmented landscapes (20-50%; Phillips et al. 2005, Etterson et al. 2014). Our daily nest survival analysis found that being parasitized by cowbirds did not increase the risk of nest predation (Table 1). Despite the large variability among females, overall nest concealment did not predict likelihood of predation. Although nest concealment does differ between successful and unsuccessful nests for some forest birds (Goodnow and Reitsma 2011) including one study of Wood Thrush (Johnson 1997), support for the nest-concealment hypothesis is generally weak (Newell and Kostalos 2007, Borgmann and Conway 2015) even for experimental manipulations of concealment (Howlett and Stutchbury 1996). This is due in part to the high diversity in nest predators and how they locate nests. In the Superb Fairy-wren (Malurus cyaneus), for instance, nest concealment in open woodland habitat reduced predation risk from visually hunting avian predators but not from small mammals or snakes who use primarily olfactory cues (Colombelli-Négrel and Kleindorfer 2009).
We found that greater nest concealment from one meter above the nest was a strong predictor of higher daily nest survival rate (Fig. 2, Table 1). Although Farnsworth and Simons (1999) did not measure nest concealment, they also found that vegetation cover above the nest (i.e., across all heights from nest to the canopy) was positively related to nesting success in Wood Thrushes. Most (136/186; 73%) Wood Thrush nests in our study were < 2.5 m above the ground, and so concealment above the nest could be highly effective against avian predators. In forest fragments only 85 km from our study area, video monitoring found that birds accounted for 64% of all Wood Thrush nest predation events (Friesen et al. 2013). For American Redstarts (Setophaga ruticilla), concealment above the nest also reduced nest predation in a study population in which avian predators were responsible for most observed nest predation events (Hannon et al. 2009). Concealment manipulation experiments, combined with camera monitoring of nests, could test if changes in concealment above the nest affects nest predation risk from avian predators.
Wood Thrush nests are relatively large and bulky and thus may be inherently difficult to hide. Some studies have found that large-bodied passerines have a higher risk of predation and speculate that large nests are more easily detectable by visually hunting predators (Willson and Gende 2000, Biancucci and Martin 2010). However, Unzeta et al. (2020) found that large-bodied passerines had a slightly lower daily nest survival rate, which could result from such species being better able to defend their nest from large-bodied predators. Even so, nest defense entails energetic costs and possible risk of injury. For large-bodied passerines, the higher detectability of large nests for visually hunting predators would presumably increase the benefits of nest concealment and yet many female Wood Thrush chose sites with low concealment.
Another question is whether females learn to build more concealed nests based on past experience with nest predation, either within or between seasons (Marzluff 1988). For Wood Thrush, overall concealment was not higher for older females (Johnson 1997) and for Hooded Warblers (Setophaga citrina), overall concealment was not higher for females re-nesting after nest predation (Howlett and Stutchbury 1997). However, a study by Hatchwell et al. (1999) found that breeding experience was an important predictor of nest placement in the cooperative-breeding Long-tailed Tit (Aegithalos caudatus) in woodland habitat. Low nests were more successful than higher nests, and failed breeders who assisted conspecifics at successful nests built their subsequent nests lower than their previous failed nests. We found that overall nest concealment in Wood Thrush increased modestly over the breeding season. A follow-up study, ideally using radio tracking to locate all of a female’s nests (e.g., Gow and Stutchbury 2013), could test if individuals better conceal their subsequent nests after nest predation occurs.
Nest concealment, even from one meter above the nest, did not reduce the likelihood of cowbird parasitism (Table 3). Although some studies have found that nest concealment reduces cowbird parasitism (Saunders et al. 2003, Sharp and Kus 2006), others have found no correlation between parasitism and nest concealment (Burhans and Thompson 1998, Banks and Martin 2000). During the host nest building and egg-laying stage, female cowbirds locate nests and then revisit them to optimally time their parasitic egg laying. A recent experimental study by White (2020) found that once female cowbirds find a host nest, they can remember not just the location but also the amount of time that has elapsed between nest visits and the number of host eggs (see also Guigueno et al. 2014). Banks and Martin (2000) showed that cowbirds were more likely to parasitize nests of host species that spent more time at the nest site during the nest-building process, and those where males vocalized more frequently near the nest. If cowbirds are relying on the behavior of pre-incubating parents to find host nests, physical concealment by vegetation may not reduce a cowbirds’ ability to locate nests.
We measured nest concealment at least one week after young fledged to reduce the possibility of attracting predators to the nest and to minimize disturbance to fledglings when they are least mobile and most vulnerable to predators. Nest concealment increased but only modestly (10%) over the breeding season, and so changes in concealment during a given nesting attempt, before we measured concealment, was likely low for most nests. However, for nests started early in the season, nests could have been less concealed at the time that cowbirds found the nest compared with a month later after the young had fledged. Our measure of nest concealment, the percentage of the nest visible from the ground and one meter away, may not fully reflect the visibility of a nest to diurnal predators with different visual abilities (e.g., visual acuity, color vision, etc.) or the conspicuousness of a nest under different lighting conditions (e.g., extent and timing of sun exposure). It is also not known for open-cup nesting passerines to what extent physical concealment of a nest within vegetation could affect wind flow patterns and reduce the olfactory cues available to nest predators (e.g., Borgo and Conover 2016).
Corticosterone and flight initiation distance (FID)
Little is known about the relationship between nest concealment, FID, and corticosterone in passerines. High nest concealment could reduce nest predation but also limit a female’s view of approaching predators and so put her at higher risk of predation (Götmark et al. 1995). Studies of ground-nesting birds in other taxa have found evidence that highly concealed nests are associated with low FID and higher risk of predation on females (e.g., Albrecht and Klvaňa 2004, Miller et al. 2007, Öst and Steele 2010, Seltmann et al. 2013). In our study, however, overall nest concealment and concealment from one meter above the nest were not significant predictors of FID in Wood Thrush (Fig. 4, Table 5). A comparative study of European birds found that more cryptically colored species had consistently shorter FIDs (Møller et al. 2019). Incubating Wood Thrush may have enhanced nest concealment via their cryptically colored plumage, which may also explain the very low FID (0 m) for almost half of females. We found that females with relatively high nests, which were presumably less vulnerable to attack from terrestrial predators, had lower FID.
The expected relationship between nest concealment and CORT is less clear, however. On the one hand, high nest concealment may induce chronic stress and cause higher CORT for incubating females because their risk of predation is high. The link between higher predation risk for adults and higher CORT has been shown experimentally at the population level for Song Sparrows (Clinchy et al. 2004). On the other hand, variation in nest concealment among females may be a result of individual variation in risk-taking tolerance and underlying CORT levels. Eider Duck females that had a bold phenotype (short FID, low stress responsiveness) were more likely to select nest sites in which predation risk to the female was higher, but nest predation was lower (Seltmann et al. 2013). A positive relationship between FID and CORT has also been shown in comparisons between urban and rural populations of Dark-eyed Juncos (Junco hyemalis), with evidence for microevolution driving these changes (Atwell et al. 2012). For Wood Thrush, however, nest concealment, even at one meter above the nest, had no effect on the initial CORT concentration of incubating females (Fig. 3, Table 4).
We also found no relationship between FID and initial or 30-min CORT (Fig. 5). Although our sample sizes were small, this preliminarily suggests that variation in concealment among individual female Wood Thrush may not lie along a risk-prone versus risk-averse axis (e.g., Seltmann et al. 2013). Further studies could test whether FID behavior is consistent for a given female (e.g., across nesting attempts), whether low FID reduces nest predation risk and increases female mortality, and experimentally manipulate nest concealment to determine if this affects female FID and CORT.
Conclusion
Many studies have explored how edge effects, small fragment size, and low landscape forest cover reduce nesting success in Wood Thrushes and other forest birds (Hoover et al. 1995, Faaborg et al. 2010). Our study suggests that nest placement by females, specifically with high nest concealment from above, could partly mitigate the costs of this high predation pressure. Predator communities may vary greatly across forest fragments of different sizes, and this can vary by region and by year (Andren 1992, Chalfoun et al. 2002), which could impact the effectiveness of overhead concealment. Future research on the effectiveness of above-nest concealment in forest birds should test the mechanism by which concealment increases nest success, for instance whether this reduces predation by avian predators, and whether females change their nest placement strategy with experience. Above-nest concealment may be ineffective for some forest birds, like the ground-nesting Veery (Catharus fuscescens), because nest predation is almost entirely by mammalian predators that do not rely on vision for finding prey (Goguen and Murray 2021). Few studies of nest concealment in songbirds have investigated if this imposes negative physiological or survival costs on nesting females. We found that overall nest concealment was not correlated with flight initiation distance or initial corticosterone concentration of incubating female Wood Thrushes; however, nest concealment manipulations are needed to further explore whether such trade-offs exist.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
We gratefully acknowledge the excellent field assistance of Amy Wilson. We also thank the various public and private landowners who allowed us access to forest fragments for our study. This research was funded by a Discovery Grant and Research Tools and Instruments Grant (BJMS) from the Natural Sciences and Engineering Research Council of Canada, the Schad Foundation, and York University.
DATA AVAILABILITY
Data is available at https://osf.io/mh2f5/.
LITERATURE CITED
Albrecht, T., and P. Klvaňa. 2004. Nest crypsis, reproductive value of a clutch and escape decisions in incubating female mallards Anas platyrhynchos. Ethology 110:603-613. https://doi.org/10.1111/j.1439-0310.2004.00992.x
Amat, J. A., and J. A. Masero. 2004. Predation risk on incubating adults constrains the choice of thermally favourable nest sites in a plover. Animal Behaviour 67:293-300. https://doi.org/10.1016/j.anbehav.2003.06.014
Andren, H. 1992. Corvid density and nest predation in relation to forest fragmentation: a landscape perspective. Ecology 73:794-804. https://doi.org/10.2307/1940158
Arnold, T. W. 2010. Uninformative parameters and model selection using Akaike’s Information Criterion. Journal of Wildlife Management 74:1175-1178. https://doi.org/10.1111/j.1937-2817.2010.tb01236.x
Atwell, J. W., G. C. Cardoso, D. J. Whittaker, S. Campbell-Nelson, K. W. Robertson, and E. D. Ketterson. 2012. Boldness behavior and stress physiology in a novel urban environment suggest rapid correlated evolutionary adaptation. Behavioral Ecology 23:960-969. https://doi.org/10.1093/beheco/ars059
Banks, A. J., and T. E. Martin. 2000. Host activity and the risk of nest parasitism by Brown-headed Cowbirds. Behavioral Ecology 12:31-40. https://doi.org/10.1093/oxfordjournals.beheco.a000375
Benson, T. J., J. D. Brown, and J. C. Bednarz. 2010. Identifying predators clarifies predictors of nest success in a temperate passerine. Journal of Animal Ecology 79:225-234. https://doi.org/10.1111/j.1365-2656.2009.01604.x
Biancucci, L., and T. E. Martin. 2010. Can selection on nest size from nest predation explain the latitudinal gradient in clutch size? Journal of Animal Ecology 79:1086-1092. https://doi.org/10.1111/j.1365-2656.2010.01720.x
Bonier, F., P. R. Martin, I. T. Moore, and J. C. Wingfield. 2009. Do baseline glucocorticoids predict fitness? Trends in Ecology Evolution 24:634-642. https://doi.org/10.1016/j.tree.2009.04.013
Borgmann, K. L., and C. J. Conway. 2015. The nest-concealment hypothesis: new insights from a comparative analysis. Wilson Journal of Ornithology 127:646-660. https://doi.org/10.1676/14-162.1
Borgo, J. S., and M. R. Conover. 2016. Visual and olfactory concealment of duck nests: influence on nest site selection and success. Human-Wildlife Interactions 10(1):110-121. https://doi.org/10.26077/0jbj-yr29
Boyd, B. P. 2022. Carry-over effects of forest fragmentation on breeding adult Wood Thrushes (Hylocichla mustelina). Dissertation. York University, Toronto, Ontario, Canada. https://yorkspace.library.yorku.ca/xmlui/bitstream/handle/10315/40670/Boyd_Brendan_P_2022_PhD.pdf?sequence=2&isAllowed=y
Burhans, D. E., and F. R. Thompson, III. 1998. Effects of time and nest-site characteristics on concealment of songbird nests. Condor 100:663-672. https://doi.org/10.2307/1369747
Chalfoun, A. D., F. R. Thompson, III, and M. J. Ratnaswamy. 2002. Nest predators and fragmentation: a review and meta-analysis. Conservation Biology 16:306-318. https://doi.org/10.1046/j.1523-1739.2002.00308.x
Clinchy, M., L. Zanette, R. Boonstra, J. C. Wingfield, and J. N. M. Smith. 2004. Balancing food and predator pressure induces chronic stress in songbirds. Proceedings of the Royal Society London B 271:2473-2479. https://doi.org/10.1098/rspb.2004.2913
Colombelli-Négrel, D., and S. Kleindorfer. 2009. Nest height, nest concealment, and predator type predict nest predation in Superb Fairy-wrens (Malurus cyaneus). Ecological Research 24:921-928. https://doi.org/10.1007/s11284-008-0569-y
Conkling, T. J., T. L. Pope, K. N. Smith, H. A. Mathewson, M. L. Morrison, R. N. Wilkins, and J. W. Cain, III. 2012. Black-Capped Vireo nest predator assemblage and predictors for nest predation. Journal of Wildlife Management 76(7):1401-1411. https://10.1002/jwmg.388
Done, T., E. A. Gow, and B. J. M. Stutchbury. 2011. Corticosterone stress response and plasma metabolite levels during breeding and molt in a free-living migratory songbird, the Wood Thrush (Hylocichla mustelina). General and Comparative Endocrinology 171:176-182. https://doi.org/10.1016/j.ygcen.2011.01.006
Driscoll, M. J. L., T. Donovan, R. Mickey, A. Howard, and K. K. Fleming. 2005. Determinants of Wood Thrush nest success: a multi-scale, model selection approach. Journal of Wildlife Management 69:699-709. https://doi.org/10.2193/0022-541X(2005)069[0699:DOWTNS]2.0.CO;2
Eng, M. L., B. J. M. Stutchbury, D. M. Burke, and K. A. Elliot. 2011. Influence of forest management on pre- and post-fledging productivity of a Neotropical migratory songbird in a highly fragmented landscape. Canadian Journal of Forest Research 41:2009-2014. https://doi.org/10.1139/x11-119
Etterson, M. A., R. Greenberg, and T. Hollenhorst. 2014. Landscape and regional context differentially affect nest parasitism and nest predation for Wood Thrush in central Virginia. Condor 116:205-214. https://doi.org/10.1650/CONDOR-13-045.1
Evans M., E. Gow, R. R. Roth, M. S. Johnson, and T. J. Underwood. 2020. Wood Thrush species account. Birds of the World, Cornell Lab of Ornithology, Ithaca, New York, USA. https://birdsoftheworld.org/bow/species/woothr/cur/introduction
Faaborg, J., R. T. Holmes, A. D. Anders, K. L. Bildstein, K. M. Dugger, S. A. Gauthreaux, Jr., P. Heglund, K. A. Hobson, A. E. Jahn, D. H. Johnson, S. C. Latta, D. J. Levey, P. P. Marra, C. L. Merkord, E. Nol, S. I. Rothstein, T. W. Sherry, T. S. Sillett, F. R. Thompson, III, and N. Warnock. 2010. Conserving migratory land birds in the New World: do we know enough? Ecological Applications 20:398-418. https://doi.org/10.1890/09-0397.1
Farnsworth, G. L., and T. R. Simons. 1999. Factors affecting nesting success of Wood Thrushes in Great Smoky Mountains National Park. Auk 116:1075-1082. https://doi.org/10.2307/4089686
Friesen, L. E., G. Casbourn, V. Martin, and R. J. Mackay. 2013. Nest predation in an anthropogenic landscape. Wilson Journal of Ornithology 125:562-569. https://doi.org/10.1676/12-169.1
Goguen, C. B., and L. D. Murray. 2021. Nest survival, predator assemblage, and patterns of predation at Veery (Catharus fuscescens) nests in a mature forest landscape. Wilson Journal of Ornithology 132:284-294. https://doi.org/10.1676/1559-4491-132.2.284
Goodnow, M. L., and L. R. Reitsma. 2011. Nest-site selection in the Canada Warbler (Wilsonia canadensis) in central New Hampshire. Canadian Journal of Zoology 89:1172-1177. https://doi.org/10.1139/z11-094
Götmark, F., D. Blomqvist, O. C. Johansson, and J. Bergkvist. 1995. Nest site selection: a trade-off between concealment and view of surroundings? Journal of Avian Biology 26:305-312. https://doi.org/10.2307/3677045
Gow, E. A., and B. J. M. Stutchbury. 2013. Understanding sex differences in parental effort in a migratory songbird: examining a sex-specific trade-off between reproduction and molt. Condor 115:640-649. https://doi.org/10.1525/cond.2013.120091
Guigueno, M. F., D. A. Snow, S. A. MacDougall-Shackleton, and D. F. Sherry. 2014. Female cowbirds have more accurate spatial memory than males. Biology Letters 10:20140026. https://doi.org/10.1098/rsbl.2014.0026
Hackemack, T. F., Z. G. Loman, S. K. Riffell, and S. A. Rush. 2016. Do nest size and shape characteristics affect nest parasitism rates? Wildlife Society Bulletin 40:722-726. https://doi.org/10.1002/wsb.707
Hannon, S. J., S. Wilson, and C. A. McCallum. 2009. Does cowbird parasitism increase predation risk to American Redstart nests? Oikos 118:1035-1043. https://doi.org/10.1111/j.1600-0706.2008.17383.x
Hatchwell, B. J., A. F. Russel, M. K. Fowlie, and D. J. Ross. 1999. Reproductive success and nest-site selection in a cooperative breeder: effect of experience and a direct benefit of helping. Auk 116:355-363. https://doi.org/10.2307/4089370
Holway, D. A. 1991. Nest-site selection and the importance of nest concealment in the Black-throated Blue Warbler. Condor 93:575-581. https://doi.org/10.2307/1368189
Hoover, J. P., M. C. Brittingham, and L. J. Goodrich. 1995. Effects of forest patch size on nesting success of Wood Thrushes. Auk 112:146-155. https://doi.org/10.2307/4088774
Hoover, J. P., and S. K. Robinson. 2007. Retaliatory mafia behavior by a parasitic cowbird favors host acceptance of parasitic eggs. Proceedings of the National Academy of Sciences 104:4479-4483. https://doi.org/10.1073/pnas.0609710104
Howlett, J. S., and B. J. Stutchbury. 1996. Nest concealment and predation in Hooded Warblers: experimental removal of nest cover. Auk 113:1-9. https://doi.org/10.2307/4088930
Howlett, J. H., and B. J. M. Stutchbury. 1997. Within-season dispersal, nest-site modification, and predation in renesting Hooded Warblers. Wilson Bulletin 109:643-649.
Ibáñez-Álamo, J. D., R. D. Magrath, J. C. Oteyza, A. D. Chalfoun, T. M. Haff, K. A. Schmidt, R. L. Thompson, and T. E. Martin. 2015. Nest predation research: recent findings and future perspectives. Journal of Ornithology 156:247-262. https://doi.org/10.1007/s10336-015-1207-4
Johnson, M. S. 1997. The effect of age on nest concealment and its complimentary effect on production of Wood Thrush. Wilson Bulletin 109:68-73.
Kats, L. B., and L. M. Dill. 1998. The scent of death: chemosensory assessment of predation risk by prey animals. Ecoscience 5:361-394. https://doi.org/10.1080/11956860.1998.11682468
Laake, J. 2013. RMark: an R interface for analysis of capture-recapture data with MARK. AFSC Processed Rep. 2013-01, Alaska Fisheries Science Center, NOAA, National Marine Fisheries Service, Seattle, Washington, USA. https://repository.library.noaa.gov/view/noaa/4372
Langerhans, R. B. 2007. Evolutionary consequences of predation: avoidance, escape, reproduction, and diversification. Pages 177-220 in A. M. T. Elewa, editor. Predation in organisms. Springer, Berlin, Germany. https://doi.org/10.1007/978-3-540-46046-6_10
Latif, Q. S., S. K. Heath, and J. T. Rotenberry. 2012. Effects of parents and Brown-headed Cowbirds (Molothrus ater) on nest predation risk for a songbird. Ecology and Evolution 2(12):3079-3097. https://doi.org/10.1002/ece3.411
Li, S., J. Qin, Z. Jin, W. Li, and H. Yan. 2018. Experimental test of the concealment hypothesis using Oriental Skylark (Alauda gulgula) nests on the Tibet Plateau. Russian Journal of Ecology 49:588-590. https://doi.org/10.1134/S1067413618660049
Lorenzana J. C., and S. G. Sealy. 1999. A meta-analysis of the impact of parasitism by the Brown-headed Cowbird on its hosts. Studies in Avian Biology 18:241-253. https://sora.unm.edu/sites/default/files/SAB_018_1999%20P241-253_A%20Meta-Analysis%20of%20the%20Impact%20of%20Parasitism%20by%20the%20Brown-Headed%20Cowbird%20on%20Its%20Hosts_Janice%20C.%20Lorenzana%2C%20Spencer%20G.%20Sealy.pdf
Martin, T. E. 1993. Nest predation and nest sites. BioScience 43:523-532. https://doi.org/10.2307/1311947
Martin, T. E. 2014. A conceptual framework for clutch-size evolution in songbirds. American Naturalist 183:313-324. https://doi.org/10.1086/674966
Martin, T. E., and J. J. Roper. 1988. Nest predation and nest-site selection of a western population of the Hermit Thrush. Condor 90:51-57. https://doi.org/10.2307/1368432
Marzluff, J. M. 1988. Do Pinyon Jays alter nest placement based on prior experience? Animal Behaviour 36:1-10. https://doi.org/10.1016/S0003-3472(88)80244-6
Miller, D. A., J. B. Grand, T. F. Fondell, and R. M. Anthony. 2007. Optimizing nest survival and female survival: consequences of nest site selection for Canada Geese. Condor 109:769-780. https://doi.org/10.1093/condor/109.4.769
Møller, A. P., W. Liang, and D. S. M. Samia. 2019. Flight initiation distance, color, and camouflage. Current Zoology 65:535-540. https://doi.org/10.1093/cz/zoz005
Montgomerie, R. D., and P. J. Weatherhead. 1988. Risks and rewards of nest defense by parent birds. Quarterly Review of Biology 63:167-187. https://doi.org/10.1086/415838
Newell, F. L., and M. S. Kostalos. 2007. Wood Thrush nests in dense understory may be vulnerable to predators. Wilson Journal of Ornithology 119:693-702. https://doi.org/10.1676/05-036.1
Öst, M., and B. B. Steele. 2010. Age-specific nest-site preference and success in eiders. Oecologia 162:59-69. https://doi.org/10.1007/s00442-009-1444-4
Peak, R. G. 2003. An experimental test of the concealment hypothesis using American Goldfinch nests. Wilson Bulletin 115:403-408. https://doi.org/10.1676/03-024
Phillips, J., E. Nol, D. Burke, and W. Dunford. 2005. Impacts of housing developments on Wood Thrush nesting success in hardwood forest fragments. Condor 107:97-106. https://doi.org/10.1093/condor/107.1.97
R Core Team. 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project-org
Robinson, W. D., T. R. Robinson, S. K. Robinson, and J. D. Brawn. 2000. Nesting success of understory forest birds in central Panama. Journal of Avian Biology 31:151-164. https://doi.org/10.1034/j.1600-048X.2000.310207.x
Romero, M. L., and J. M. Reed. 2005. Collecting baseline corticosterone samples in the field: is under 3 min good enough? Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology 140(1):73-79. https://doi.org/10.1016/j.cbpb.2004.11.004
Rotella, J. J., S. J. Dinsmore, and T. L. Shaffer. 2004. Modeling nest-survival data: a comparison of recently developed methods that can be implemented in MARK and SAS. Animal Biodiversity and Conservation 27:187-205.
Sauer, J. R., D. K. Niven, J. E. Hines, D. J. Ziolkowski, Jr., K. L. Pardieck, J. E. Fallon, and W. A. Link. 2019. The North American Breeding Bird Survey, results and analysis 1966-2019. Version 2.07.2019. USGS Patuxent Wildlife Research Center, Laurel, Maryland, USA. https://www.usgs.gov/data/north-american-breeding-bird-survey-analysis-results-1966-2019
Saunders, C. A., P. Arcese, and K. D. O’Connor. 2003. Nest site characteristics in the Song Sparrow and parasitism by Brown-headed Cowbirds. Wilson Bulletin 115:24-28. https://doi.org/10.1676/02-057
Schoech, S. J., M. A. Rensel, E. S. Bridge, R. K. Boughton, and T. E. Wilcoxen. 2009. Environment, glucocorticoids, and the timing of reproduction. General Comparative Endocrinology 163:201-207. https://doi.org/10.1016/j.ygcen.2008.09.009
Schneider, N. A., and M. Griesser. 2013: Incubating females use dynamic risk assessment to evaluate the risk posed by different predators. Behavioral Ecology 24:47-52. https://doi.org/10.1093/beheco/ars134
Seltmann, M. W., K. Jaatinem, B. B. Steele, and M. Öst. 2013. Boldness and stress responsiveness as drivers of nest-site selection in a ground-nesting bird. Ethology 120:77-89. https://doi.org/10.1111/eth.12181
Sharp, B. L., and B. E. Kus. 2006. Factors influencing the incidence of cowbird parasitism of Least Bell’s Vireos. Journal of Wildlife Management 70:682-690. https://doi.org/10.2193/0022-541X(2006)70[682:FITIOC]2.0.CO;2
Thys, B., Y. Lambreghts, R. Pinxten, and M. Eens. 2019. Nest defence behavioural reaction norms: testing life-history and parental investment theory predictions. Royal Society Open Science 6:182180. https://doi.org/10.1098/rsos.182180
Unzeta, M., T. E. Martin, and D. Sol. 2020. Daily nest predation decreases with body size in passerine birds. American Naturalist 196:743-754. https://doi.org/10.1086/711413
Wada, H., T. P. Hahn, and C. W. Breuner. 2007. Development of stress reactivity in White-crowned Sparrow nestlings: total corticosterone response increases with age, while free corticosterone response remains low. General and Comparative Endocrinology 150:405-413. https://doi.org/10.1016/j.ygcen.2006.10.002
Weidinger, K. 2002. Interactive effects of concealment, parental behavior and predators on the survival of open passerine nests. Journal of Animal Ecology 71:424-437. https://doi.org/10.1046/j.1365-2656.2002.00611.x
White, D. 2020. Avian egg timers: female cowbirds judge past, present, and future when making nest parasitism decisions. Frontiers in Ecology and Evolution 8:203. https://doi.org/10.3389/fevo.2020.00203
White, G. C., and K. P. Burnham. 1999. Program MARK: survival estimation from populations of marked animals. Bird Study 46:S120-139. https://doi.org/10.1080/00063659909477239
Wiebe, K. L., and K. Martin. 1998. Costs and benefits of nest cover for ptarmigan: changes within and between years. Animal Behaviour 56:1137-1144. https://doi.org/10.1006/anbe.1998.0862
Willson, M. F., and S. M. Gende. 2000. Nesting success of forest birds in southeast Alaska and adjacent Canada. Condor 100:314-325. https://doi.org/10.1093/condor/102.2.314
Wilson, R. R., and R. J. Cooper. 1998. Acadian Flycatcher nest placement: does placement influence reproductive success? Condor 100:673-679. https://doi.org/10.2307/1369748
Wingfield J. C., P. Deviche, S. Sharbaugh, L. B. Astheimer, R. Holberton, R. Suydam, and K. Hunt. 1994. Seasonal changes of the adrenocortical responses to stress in Redpolls, Acanthis flammea, in Alaska. Journal of Experimental Zoology 270(4):372-380. https://doi.org/10.1002/jez.1402700406
Fig. 1
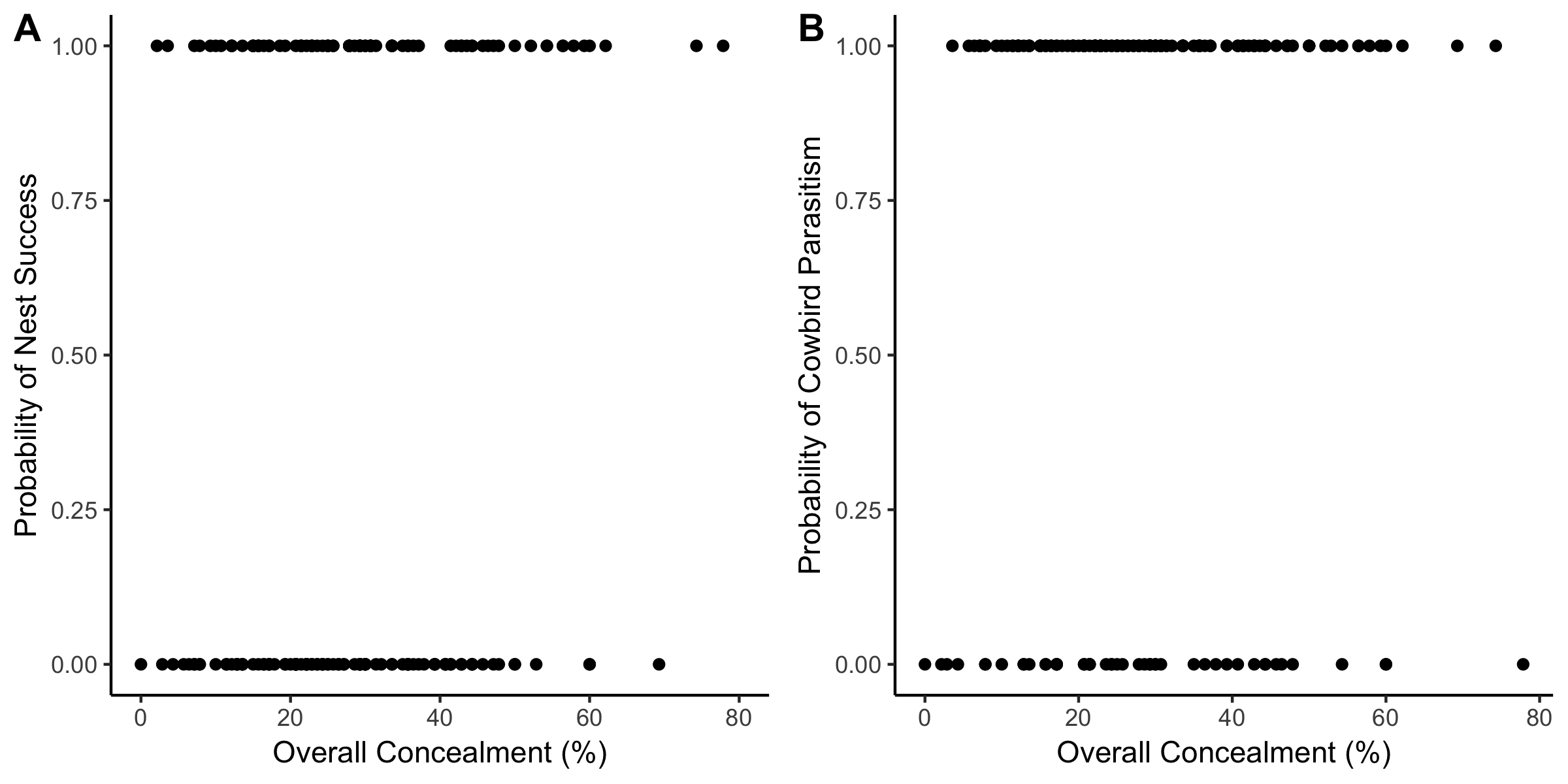
Fig. 1. Logistic regression plots showing the probability of (A) nest success and (B) cowbird parasitism in Wood Thrush (Hylocichla mustelina) nests (n = 186) in relation to overall nest concealment (%). Overall nest concealment was not a good predicator of daily nest survival or cowbird parasitism (see Table 1).
Fig. 2
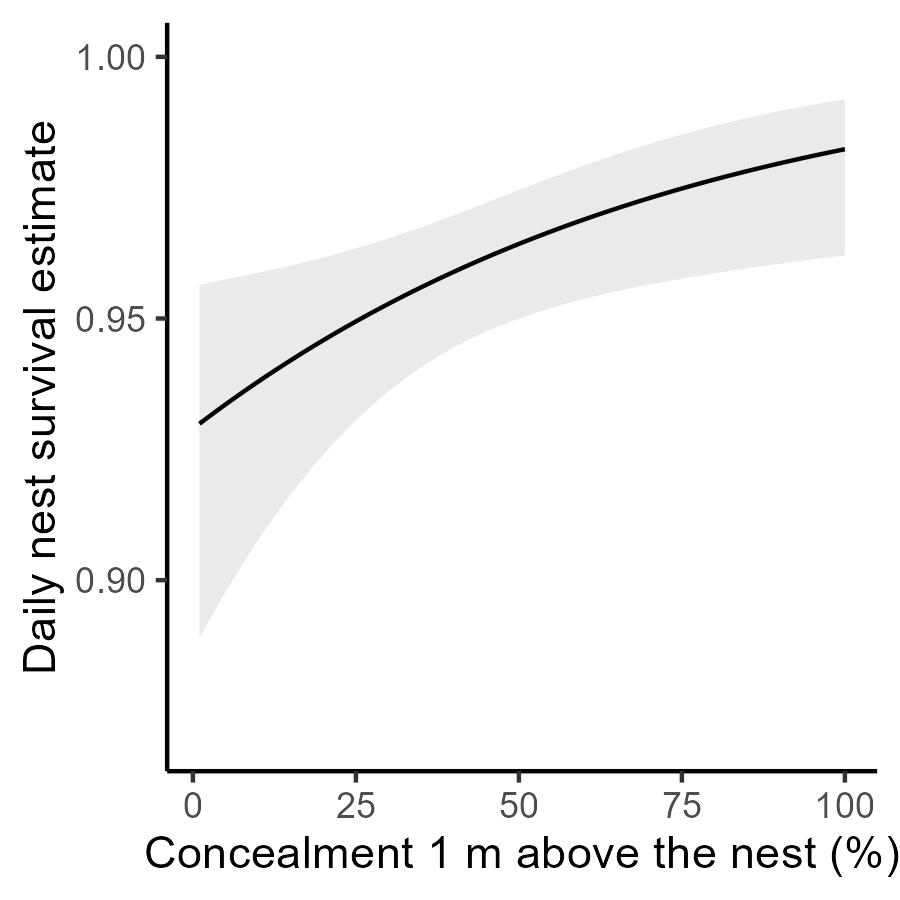
Fig. 2. Estimated relationship (±SE) between daily nest survival rate (S) and nest concealment from 1m above the nest for Wood Thrush nests (n = 141) using beta parameters from the best-fit model (Table 1). Shaded area is 95% confidence interval. Analysis performed in RMARK.
Fig. 3
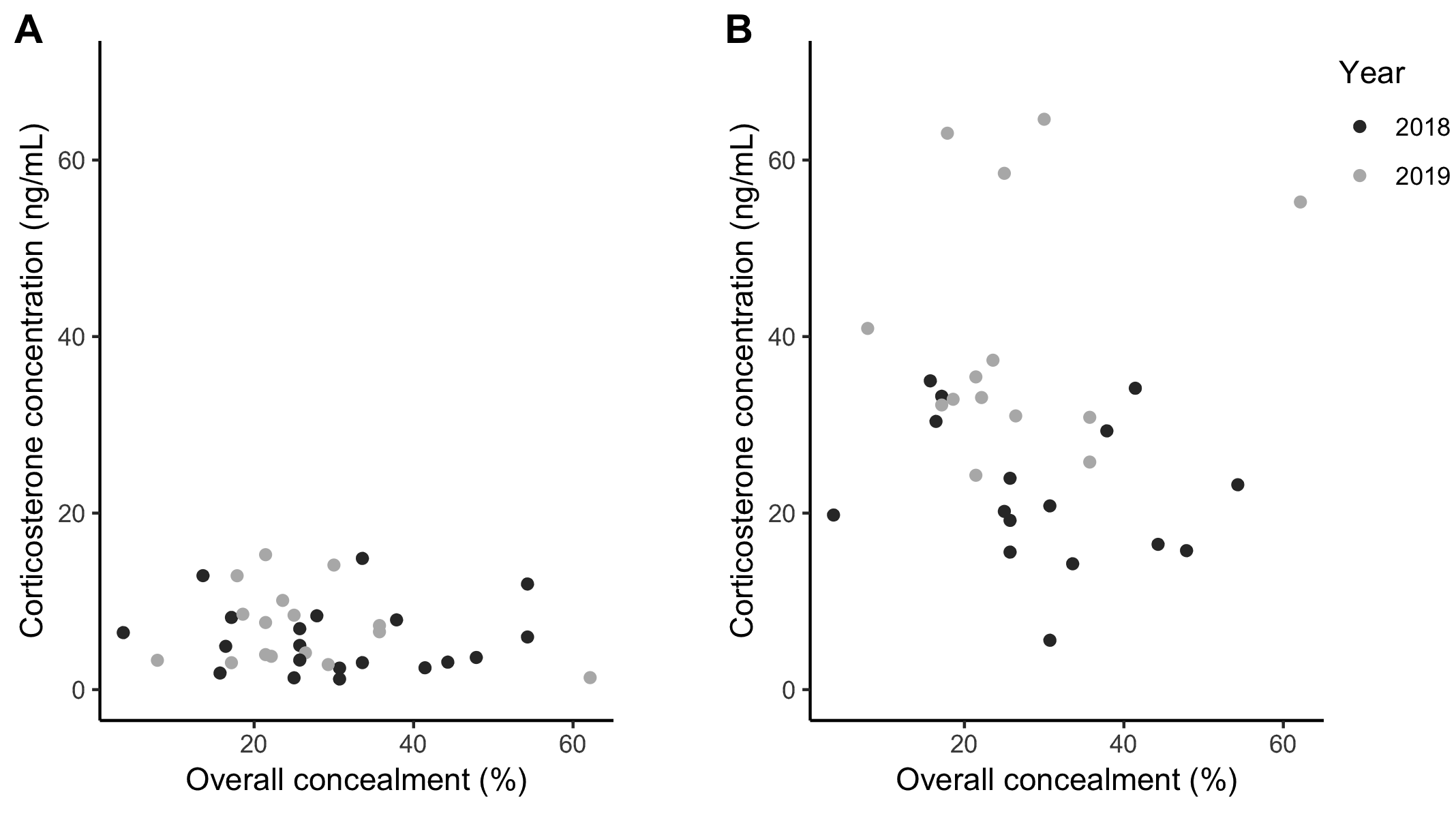
Fig. 3. The relationship between corticosterone concentration (ng/mL) and overall nest concealment for (A) baseline (n = 36) and (B) 30-minute CORT samples (n = 30). Year of sampling (2018 versus 2019) indicated with grey versus black symbols. Overall nest concealment was not a strong predictor of baseline or 30-min CORT (see Table 4).
Fig. 4
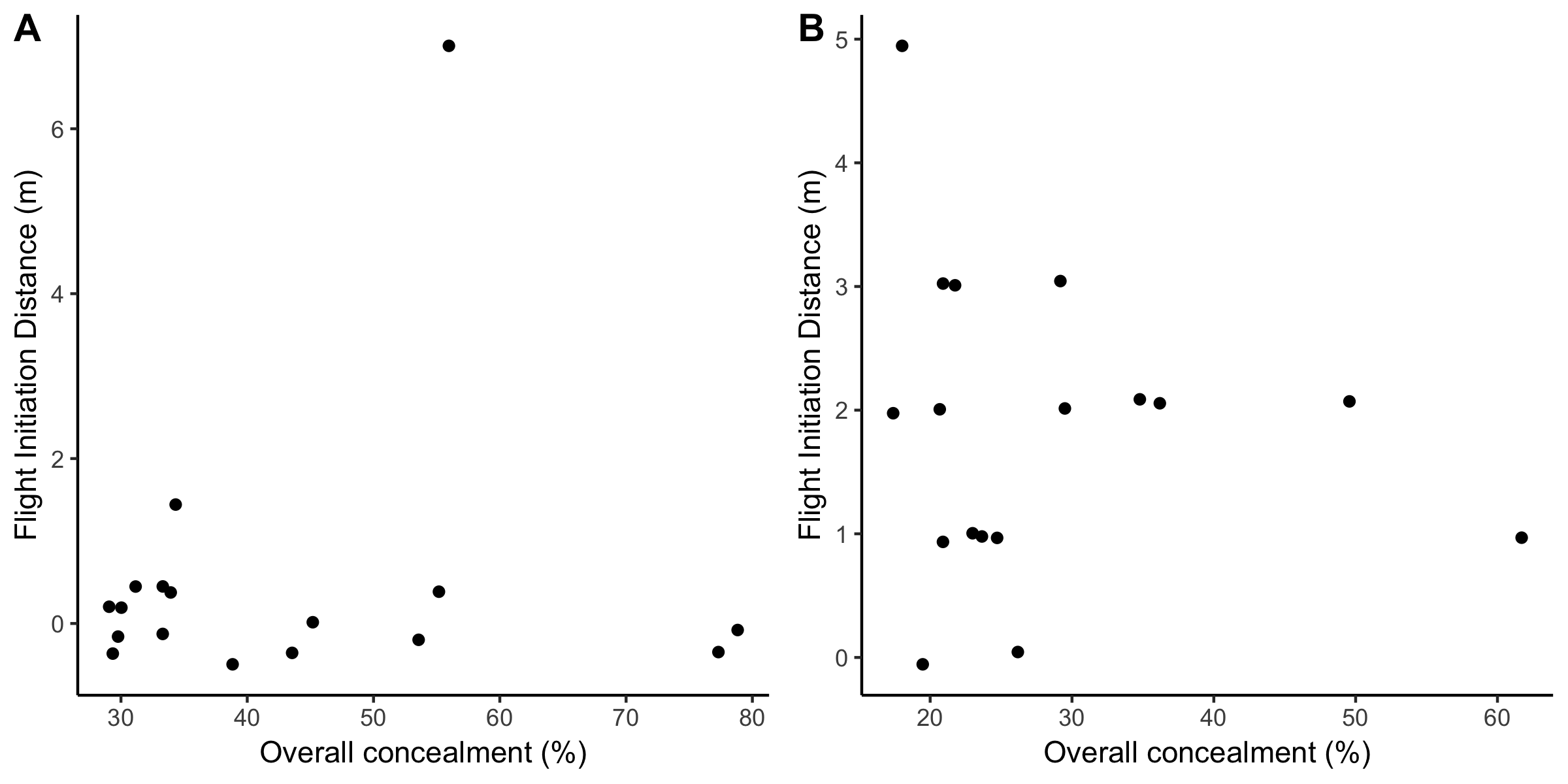
Fig. 4. The relationship between FID and overall nest concealment for 2018 (A; n = 30) and 2019 (B; n = 17). Many points were overlapping at 0m FID in 2018, and so these have been jittered for visualization. Nest concealment was not a strong predictor of FID (see Table 5).
Fig. 5
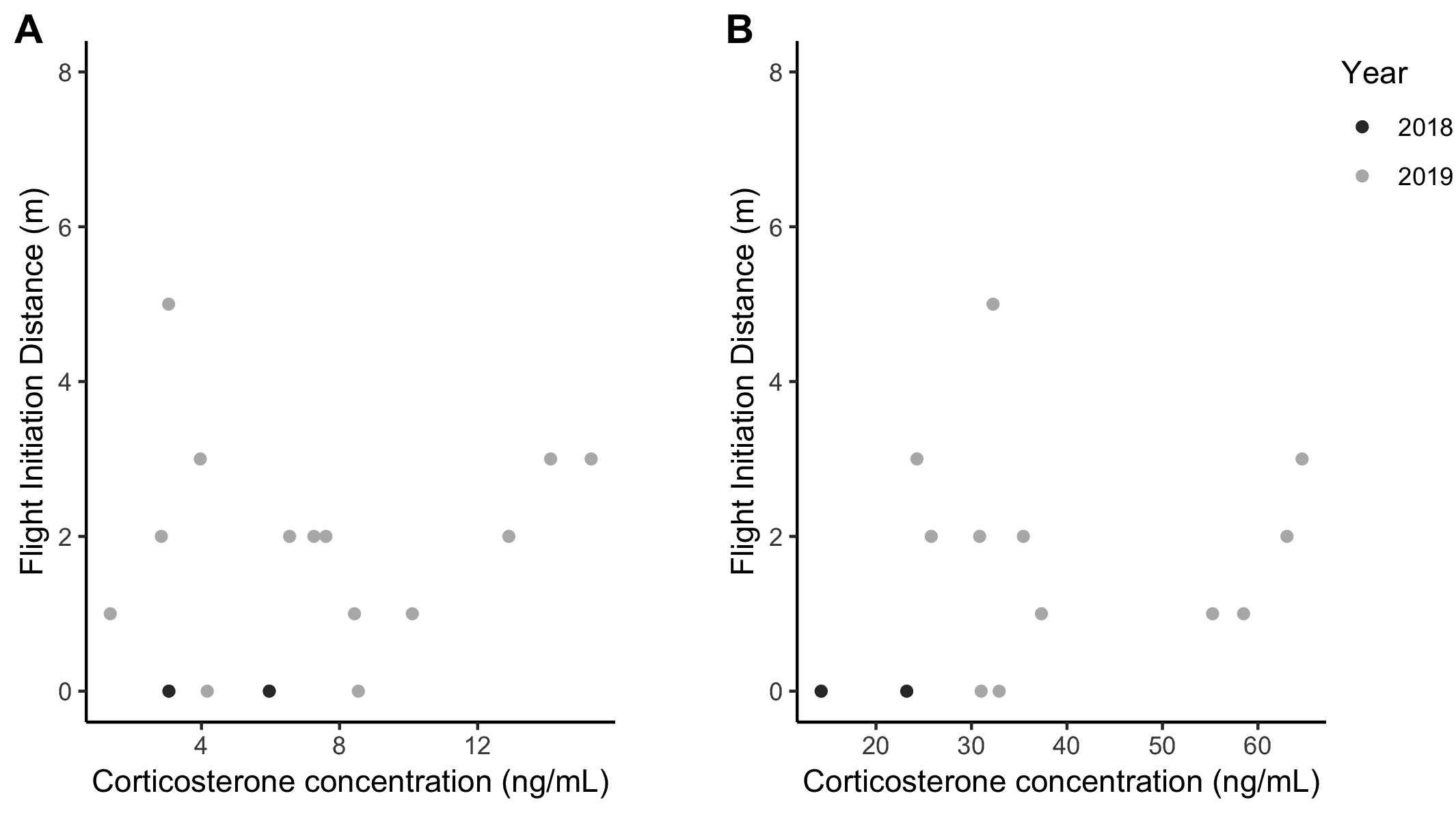
Fig. 5. The relationship between flight initiation distance and corrected corticosterone concentration from (A) baseline (n = 16) and (B) 30-minute CORT samples (n = 14), for 2018 and 2019. Neither of the CORT measures were strong predictors of FID (see Table 5).
Table 1
Table 1. Summary of model selection results for survival of Wood Thrush (Hylocichla mustelina) nests (n = 141) generated using the nest survival model in the program MARK 6.0. The list begins with the best-fitting model and is sorted by the ΔAIC value. The Wi value is the weight of each model in the model set, and sums to 1. All models except the null model S(1) included NestAge (age of nest since first egg) but different measures of nest concealment (overall, from 1 m above, from ground level), and effects of year (2017, 2018, 2019), nest height (m), and a categorical measure of fragment size (small: 11-69 ha; large: 162-500 ha).
| Model | No. of parameters | AIC | ΔAIC | Wi | Deviance |
| S(NestAge + ConcealAbove) | 3 | 331.58 | 0.00 | 0.744 | 325.57 |
| S(NestAge) | 2 | 336.31 | 4.74 | 0.070 | 332.31 |
| S(NestAge + NestHeight) | 3 | 337.54 | 5.96 | 0.038 | 331.53 |
| S(NestAge + ConcealGround) | 3 | 337.78 | 6.20 | 0.034 | 331.76 |
| S(NestAge + ConcealOverall) | 3 | 337.85 | 6.27 | 0.032 | 331.84 |
| S(NestAge + Cowbird) | 3 | 338.08 | 6.5 | 0.029 | 332.07 |
| S(NestAge + ConcealOverall + NestHeight) | 4 | 338.56 | 6.99 | 0.026 | 330.55 |
| S(NestAge + ConcealOverall + FragSize) | 4 | 339.48 | 7.90 | 0.014 | 331.47 |
| S(NestAge * ConcealOverall) | 4 | 339.83 | 8.25 | 0.012 | 331.81 |
| S(NestAge + ConcealOverall + Year) | 5 | 341.66 | 10.08 | 0.005 | 331.63 |
| S(1) | 1 | 354.28 | 22.70 | 0.000 | 352.28 |
Table 2
Table 2. Summary of beta estimates for the best model (NestAge + ConcealAbove) for survival of Wood Thrush nests (n = 141) obtained using the nest survival model in the program RMARK (see Table 1).
| Estimate | SE | Lower 95%; CI | Upper 95% CI | |
| S: (Intercept) | 4.3041 | 0.3776 | 3.5640 | 5.0440 |
| S: NestAge | -0.1020 | 0.0233 | -0.1476 | -0.0563 |
| S: ConcealAbove | 0.0145 | 0.0058 | 0.0032 | 0.0258 |
Table 3
Table 3. The z-values and p-values for a binomial generalized linear model (GLM) predicting cowbird parasitism (n = 186 nests) using multiple predictors (overall nest concealment, concealment 1m above, nest height, fragment size category, and year).
| Predictor | Coefficient estimate | Standard error | z-value | p-value |
| Overall concealment | 0.0088 | 0.0183 | 0.485 | 0.628 |
| Concealment 1 m above | -0.0105 | 0.0110 | -0.961 | 0.337 |
| Nest height | -0.3988 | 0.2786 | -1.431 | 0.152 |
| Fragment size | 0.7310 | 0.3679 | 1.987 | 0.047 |
| Year | 0.2080 | 0.2491 | 0.835 | 0.404 |
Table 4
Table 4. The t-values and p-values for a gamma generalized linear model (GLM) predicting initial corticosterone (CORT within 3 min of capture; n = 36) and 30-min CORT levels (n = 30) of incubating female Wood Thrush using multiple predictors (overall nest concealment, concealment 1m above, nest height, fragment size (small vs large), and year).
| Initial CORT | 30-minute CORT | |||
| Predictor | t-value | p-value | t-value | p-value |
| Overall concealment | 0.309 | 0.759 | -0.840 | 0.409 |
| Concealment 1 m above | -0.654 | 0.518 | 0.945 | 0.354 |
| Nest height | -1.520 | 0.139 | -0.585 | 0.564 |
| Fragment size | 1.708 | 0.098 | 0.525 | 0.605 |
| Year | 1.057 | 0.299 | 4.359 | 0.001 |
Table 5
Table 5. The z-values and p-values for a Poisson generalized linear model (GLM) predicting FID (n = 14) using multiple different predictors (nest concealment, concealment 1m above, nest height, fragment size, baseline CORT, 30-min CORT, and year).
| Predictor | Coefficient estimate | Standard error | z-value | p-value |
| Overall concealment | 0.0393 | 0.0430 | 0.917 | 0.350 |
| Concealment 1 m above | -0.0543 | 0.0356 | -1.528 | 0.126 |
| Nest height | -1.5323 | 0.0705 | -2.174 | 0.030 |
| Fragment size | 0.8762 | 0.8584 | 1.021 | 0.307 |
| Initial CORT | -0.1070 | 0.1107 | -0.996 | 0.334 |
| 30-min CORT | 0.0220 | 0.0317 | 0.692 | 0.489 |
| Year | 20.340 | 5931.8 | 0.003 | 0.997 |