







The following is the established format for referencing this article:
Brooks, W. E., J. Boersma, N. Paprocki, P. Wimberger, and S. Hotaling. 2023. Community science for enigmatic ecosystems: using eBird to assess avian biodiversity on glaciers and snowfields. Journal of Field Ornithology 94(1):6.ABSTRACT
Mountain glaciers and snowfields are rapidly receding because of climate warming. Species living in these habitats remain poorly studied, likely because of the remoteness and ruggedness of their terrain. We leveraged community science data from eBird—an online database of bird observations from around the world—to characterize bird use of mountain glaciers and snowfields. We estimated total bird biodiversity and preference for glaciers and snowfields over nearby, ice-adjacent habitats. We used field notes from eBird users and breeding codes to extend our data set to include insight into habitat usage and behavior. Finally, we compared our community-science approach to previous studies that used traditional (i.e., professional) field survey methods. We identified considerable avian biodiversity in glacier and snowfield habitat (46 species) with four specialists that appeared to prefer glaciers and snowfields over nearby, ice-adjacent habitats. Birds appeared to primarily use the glacier and snowfield habitats for foraging. When community science data was compared to traditional methods, results were similar, but community science resulted in a higher species diversity estimate. Our findings suggest glacier and snowfield retreat threatens specialist species via loss of nutrient resources. Additionally, community science data appears valuable for characterizing difficult to access areas, but traditional surveys are still useful for more rigorous quantification of avian biodiversity.
RESUMEN
Glaciales de montaña y campos de nieve están retrocediendo rápidamente por causa del calentamiento climático. Las especies que viven en estos hábitats aún son poco estudiadas, probablemente por ser lugares remotos y tener terreno rugoso. Aprovechamos los datos de la ciencia comunitaria de eBird- una base de datos en línea de las observaciones de aves alrededor del mundo- para caracterizar el uso de los glaciares de montaña y campos de nieve por aves. Estimamos la biodiversidad total de aves y la preferencia por glaciares y campos de nieve cercanos sobre los hábitats adyacentes al hielo. Utilizamos las notas de campo de usuarios de eBird y códigos de reproducción para extender nuestro conjunto de datos e incluir conocimiento sobre el uso del hábitat y comportamiento. Finalmente comparamos nuestra aproximación basada en la ciencia comunitaria con estudios previos que usaron métodos de estudio de campo tradicionales (i.e. profesionales). Identificamos una biodiversidad de aves considerable en hábitats glaciales y campos de nieve (46 especies) con cuatro especies especialistas que aparentemente prefirieron glaciales y campos de nieve sobre hábitats adyacentes al hielo. Las aves aparentemente utilizan hábitats glaciales y campos de nieve principalmente para forrajear. Cuando comparamos los datos de la ciencia comunitaria con los de los métodos tradicionales, los resultados fueron similares, pero la ciencia comunitaria resulto en una diversidad más alta de especies estimadas. Nuestros resultados sugieren que el retroceso de los glaciales y los campos de nieve amenaza especies especialistas por medio de la perdida de recursos nutriticionales. Adicionalmente, los datos de la ciencia comunitaria son valiosos para caracterizar áreas difíciles de acceder, pero métodos de estudio tradicionales son útiles todavía para una cuantificación más rigurosa de la biodiversidad de aves.
INTRODUCTION
Contemporary climate change is driving global glacier recession reflected in mass loss and increased meltwater runoff (Moore et al. 2009, Bolch et al. 2012, Bliss et al. 2014). Recession of the mountain cryosphere, primarily glaciers and perennial snowfields, will negatively impact the considerable biodiversity these ecosystems host (Cauvy-Fraunié and Dangles 2019, Stibal et al. 2020, Hotaling et al. 2021). Glacier biodiversity is dominated by microbial communities thriving within, below, and downstream of glacial ice (Hotaling et al. 2017). A range of macroinvertebrates have also been described from glacier ecosystems, commonly acting as resource subsidies for larger taxa (e.g., birds; Hotaling et al. 2020). Vertebrates also use glaciers for a host of activities including thermoregulation, food caching, and travel (Rosvold 2016), but the extent of vertebrate links to glacier habitats remains poorly characterized. This knowledge gap is likely due to the rarity of vertebrates relative to smaller organisms paired with the remote, difficult-to-access terrain of glacier ecosystems, and the ease of sampling invertebrates relative to vertebrate field surveys, highlighting the need for alternative survey methods. Given the ongoing recession of the mountain cryosphere, more information about the biodiversity present in these habitats is urgently needed to quantify the impacts of glacier loss.
Among vertebrates, glaciers and perennial snowfields may represent particularly important habitats for birds. However, mountain birds are less studied than lowland species, despite widespread evidence for climate change impacting their reproduction, survival, and distributions (Scridel et al. 2018). Although no systematic surveys of birds on glaciers and snowfields have been conducted, observations from literature reviews and a recent camera trap study have recorded just 22 bird species associated with glaciers worldwide (Goodman 1971, Rosvold 2016; S. Hotaling et al., personal communication 2022). Most birds appear to use cryospheric habitats for foraging on wind-blown arthropods, pollen, and seeds (Crawford and Edwards 1986, Antor 1995, Shain et al. 2001, Camfield et al. 2010, Hotaling et al. 2020). Several birds also nest near (Johnson 1965, Rosvold 2016) or on glaciers (Hardy and Hardy 2008), likely because of food sources on the glacier surface and reduced predation risks.
Community science (also referred to as citizen science) is a rapidly growing source of data to fill gaps in ecological research (Silvertown 2009, Kosmala et al. 2016). Community science uses volunteers to gather data, benefiting professional researchers through high-volume, low-cost data, and the community scientists through gained knowledge and exposure to research (Silvertown 2009, Peter et al. 2021). Community science data have been successfully applied in biodiversity monitoring on large spatio-temporal scales (Chandler et al. 2017, Freeman et al. 2022), and to inform policy and decision making (Conrad and Hilchey 2011, de Sherbinin et al. 2021). The abundance of community science data presents opportunities to study rare species and habitats (Robinson et al. 2018), but these studies remain rare. For avian community science, eBird has emerged as a global leader; birdwatchers submit eBird “checklists,” i.e., lists of their field observations, containing bird abundances, qualitative observations, and sampling effort (Sullivan et al. 2009). These checklists can be aggregated to calculate trends and distributions of birds in locations or habitats of interest. In general, eBird-based estimates of abundances align well with traditional surveys (Callaghan and Gawlik 2015, Callaghan et al. 2017).
In this study, we used eBird data to clarify avian biodiversity in an understudied, climate-change-threatened ecosystem: mountain glaciers and perennial snowfields. Through this objective, we present a framework for using a public, community science-derived database (eBird) to characterize links between a taxonomic group of interest (birds) and difficult-to-study habitats (glaciers and perennial snowfields in mountain ecosystems). Specifically, we compared avian communities and biodiversity in glaciers and snowfields and nearby, ice-adjacent habitats, and described bird behavior in these habitats from observer notes. We show that eBird data can be used to rigorously quantify avian biodiversity and document habitat and behavior, highlighting the power of community science projects for studying uncommon and understudied ecosystems.
METHODS
Data acquisition
We acquired eBird data (Sullivan et al. 2009) from alpine areas in southern British Columbia (BC), Washington (WA), and Oregon (OR) to document avian biodiversity on mountain glaciers and snowfields, as well as in nearby, ice-adjacent habitats. We chose this study region because it contains many mountain glaciers and is relatively highly populated, meaning it receives many eBird submissions compared to other alpine areas. We identified 129 focal alpine bird species (full list is included in Appendix 1) based primarily on two criteria: (1) they were detected in mountain bird surveys by Boyle and Martin (2015) or (2) have been reported in four frequently used, high-altitude (2149–3157-m elevation) eBird hotspots representative of our study region: Panorama Point (Mount Rainier National Park, WA, 46.804407, -121.721005), Blackcomb Mountain (BC, 50.092500, -122.887778), Mt. Hood (Hood River Co., OR, 45.372360, -121.672211), South Sister summit (Lane Co., OR, 44.102726, -121.769070). eBird hotspots are public eBird locations created by eBird users that generally reflect popular birding locations. Species reported at each hotspot are aggregated and summarized on eBird.org. Additionally, based on known high-altitude migration in Baird’s Sandpiper (Calidris bairdii; Shewey and Blount 2017) and ice worm foraging by Semipalmated Plovers (Charadrius semipalmatus; Goodman 1971), 12 common inland shorebird species were included to reflect the possible use of glaciers and snowfields during migration.
Collectively, we analyzed three separate data types reported by eBird users: observations, field notes, and breeding codes. Observation data are counts of each species reported by users, field notes are entered by users in a free text field for each observation, and breeding codes are optionally entered by users to document specific breeding behaviors. To acquire the data, we downloaded the “basic” eBird data set containing all eBird checklists submitted worldwide prior to June 2021. We filtered the data set in R version 4.1.0 using the auk_filter command of the auk package (Strimas-Mackey et al. 2018, RStudio Team 2020) to only include checklists submitted in our focal region (WA, OR, and BC) that were marked as “complete.” Users mark their checklists as complete if they have reported all of the species they observed at a location. These complete checklists are ideal for measuring species diversity and abundance because they reduce overreporting of species considered more interesting to birdwatchers (e.g., rare species) and they can be used to infer the absence of species (those not reported). We then extracted complete checklists that included any of our 129 focal species.
Spatial filtering
We spatially filtered our data set to bin checklists by those close to or including glaciers and snowfields versus nearby, ice-adjacent habitats. We used QGIS (QGIS Development Team 2022) to create polygon layers defining our two habitat types. We considered both glaciers and snowfields together following Rosvold (2016). We used a permanent ice polygon layer to outline glaciers and snowfields (Natural Resources Canada et al. 2004), and added a 100-m buffer around each polygon because it seemed likely that eBird users would observe birds from the edge of a glacier or snowfield. We included ice-adjacent habitat as a biodiversity comparison and to determine if any species had preference for glaciers and snowfields. We created “ice-adjacent” habitat polygons with a 2-km buffer around all glaciers and snowfields, which resulted in a layer roughly equal in area to the glacier and snowfield layer. We used the select by location tool to identify eBird checklists intersecting either of our habitat categories and extracted elevation data for each checklist from a 1-km resolution elevation raster layer using the point sampling tool (U.S. Geological Survey and Natural Resources Canada 2007).
After binning checklists into “glaciers and snowfield” or “ice-adjacent” habitat categories, we zero-filled and cleaned the final dataset. We used the auk_zerofill command to add zeros where a species was not observed, allowing each checklist to represent both presence and absence. We elected to remove checklists from northern British Columbia (latitude > 54.7° N) as glaciers transition to sea level there, thus possibly supporting a different bird community than the montane glaciers and snowfields that we were focused on. Because stringent filtering, particularly by sampling effort, generally improves accuracy in community science analyses (Steen et al. 2019, Johnston et al. 2021, Van Eupen et al. 2021), we further filtered our zero-filled observation data to include only checklists that were stationary or with a travel distance under 1 km (85% reduction in total checklists). We chose a 1 km travel distance because it is half of the minimum width of most of our habitat polygons, making it less likely that users traveled out of the desired habitat.
Calculating biodiversity metrics and comparing habitat types
To assess biodiversity in our focal habitats, we calculated species richness, or the total number of bird species observed, and Shannon-Weiner diversity index (H’). Before calculating Shannon-Wiener diversity index, we removed 171 non-numeric observation (4% of all observations), which likely reflect community scientists marking species as present with an X, but not providing an abundance estimate. We then calculated Shannon-Wiener diversity index with the equation:
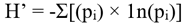 |
(1) |
“p” is the proportion of the total population made up of species “i.” To identify the most common species, we calculated the observation frequency of each species as the proportion of all checklists where a given species is present.
We compared biodiversity and community composition in glaciers and snowfields to nearby, ice-adjacent habitat. After filtering, our data set consisted of 70 glacier and snowfield checklists and 887 ice-adjacent habitat checklists (Fig. 1). Because this 10:1 ratio ice-adjacent to glacier and snowfield checklists could bias our estimates of species richness, we resampled with replacement checklists for ice-adjacent habitats 10,000 times in sets of 70 and calculated species richness for each resampled set. Next, glacier and snowfield richness was compared to the 95% confidence interval (95th percentile) of the bootstrapped data from ice-adjacent habitat. We compared species diversity in glaciers and snowfields and ice-adjacent habitat for Shannon-Wiener indices with a Hutcheson t-test (Hutcheson 1970), using an online calculator (https://www.dataanalytics.org.uk/comparing-diversity/).
To assess community similarity between glaciers and snowfields and ice-adjacent habitat, we compared species observation frequency in each habitat with linear regression. Normality of species frequencies was tested and corrected in ice-adjacent habitats with a cube root transformation. We also compared communities with Sorensen’s Coefficient (CC; Sorensen 1948) using the formula:
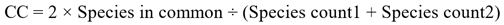 |
(2) |
“Species count1” is the species richness of glaciers and snowfields and “Species count2” is the species richness of ice-adjacent habitats. Finally, we examined habitat preference by subtracting the observation frequency of each species in nearby, ice-adjacent habitat from glaciers and snowfields; positive values represented species more common in glaciers and snowfields. We performed a series of Pearson’s Chi square tests to test for significant habitat associations.
Using observer notes to understand habitat usage and behavior
We used observer field notes as an additional method for identifying glacier and snowfield habitat use by birds. eBird users report a wide array of field notes that represent a largely untapped source of descriptive data (Slager 2020). As a separate count of habitat use, we identified field notes from the full checklist set (not filtered by habitat polygons) that included the keywords “glacier” and “snowfield” as well as variations of each (e.g., “snowfield” or “snow field”). We manually sorted field notes to remove false positives, thus only counting descriptions of birds in the habitat, not near it. We summed observations with notes for each bird species.
We assessed bird behavior on glaciers and snowfields using two previously defined data sets for glaciers and snowfields, checklists identified in our (1) spatial analysis and (2) field notes survey. We extracted descriptions of behavior using the same methods as habitat; we used keywords to identify relevant notes and manually reviewed them. We examined birds flying over glaciers and snowfields because they may be associated with the habitat despite not contacting snow or ice. We summed birds flying over (keyword: “flyover”) or using the flyover (F) breeding code. Similarly, because unseen birds heard by observers may be vocalizing from a glacier and snowfield, or nearby, ice-adjacent habitat we summed heard-only species in field notes (keywords: “heard only”). Finally, to examine how glaciers and snowfields function as a nutrient source, we identified foraging behavior (keywords: “feed, eat, forage”) and food choice in field notes (keywords: “spider, bug, insect, seed, pollen, worm,” and “ice worm”), and used the carrying food (CF) breeding code. We specifically sought descriptions of ice worm foraging because this is a well-established but understudied resource for glacier-associated birds (Hotaling et al. 2020).
Comparing community science data to traditional surveys
To compare diversity estimates generated by community science with traditional methods (e.g., transects) we performed two analyses. (1) We compared glacier- and snowfield-associated species identified in eBird field notes to those identified by Rosvold (2016) and a camera trap study of glacial wildlife (Hotaling et al., 2022, personal communication). We chose these two studies because Rosvold (2016) was globally focused and Hotaling et al. (2022) was published later and focused on a glacier within our study area (Paradise Glacier, Mount Rainier, WA, USA). (2) We quantified the similarity of community composition and observation frequencies for our eBird data and traditional survey study—Boyle and Martin (2015)—for the ice-adjacent habitat. We were unable to do a similar analysis for snow/ice species because there are no traditional surveys of avian diversity for that habitat. We used Boyle and Martin (2015) as our comparison data set because it shared similar habitat, region, and total survey time [eBird “ice-adjacent”: ~723 observation hours; Boyle and Martin (2015): 717 observation hours]. We compared the two data sets as described above using CC, observation frequencies, and linear regression.
RESULTS
Our initial data download included 1,984,111 eBird checklists. After spatial filtering, we identified 957 mountain checklists, comprising ~845 observation hours of survey time. Mountain checklists were not evenly distributed in space or time: dates spanned 1999–2021 but were biased to summer months in the past 10 years. Geographically, most locations were in Washington (54%) and Oregon (40%) with few in British Columbia (6%; Fig. 1). We identified 70 checklists from glaciers and snowfields (~122 observation hours) and 887 checklists from ice-adjacent habitats (~723 observation hours). We extracted 867,989 species field notes from the total data set for text analysis, 52 of which contained keywords pertaining to habitat, behavior, or diet. We extracted just one observation with a breeding code relevant to our study.
Avian biodiversity and community composition
Glacier and snowfield checklists contained 46 species (Fig. 2), and a Shannon-Weiner diversity index (H’) of 2.77. The most common species were Common Raven (Corvus corax; 47% of checklists), Gray-crowned Rosy-finch (Leucosticte tephrocotis; 46%), American Pipit (Anthus rubescens, 29%), Pine Siskin (Spinus pinus, 11%), Clark’s Nutcracker (Nucifraga columbiana, 10%), and Dark-eyed Junco (Junco hyemalis, 9%). All other species were reported in ≤ 6% of checklists. Overall, glacier and snowfield habitat exhibited lower biodiversity than nearby, ice-adjacent habitat. We identified 106 species in the ice-adjacent data set. However, after correcting for sample size, ice-adjacent habitats were still more biodiverse but the difference was much smaller at ~68 ice-adjacent species (bootstrapped 95% CI: 61–76; results in Appendix 2). Glaciers and snowfields also had lower Shannon-Weiner diversity (H’ = 2.77) than ice-adjacent habitats (H’ = 3.66; Hutcheson’s t-test: t = 14.69, p < 0.001).
Community composition was similar in glaciers and snowfields versus ice-adjacent habitats: roughly 59% of species were present in both habitats (CC = 0.59) and observation frequencies were significantly correlated (R² = 0.17, F1,127 = 26.59, p < 0.001). Four species were significantly more common in glaciers and snowfields than ice-adjacent habitats (Fig. 3): Gray-crowned Rosy-finch (Pearson’s Chi Square test, X² = 161, p < 0.001), American Pipit (X² = 10.6, p = 0.001), Rock Wren (Salpinctes obsoletus; X² = 5.61, p = 0.018), and American Kestrel (Falco sparverius; X² = 5.09, p = 0.024). All were evenly distributed across our study region except Rock Wren, which was primarily reported in Oregon.
Observer-reported habitat and behavior
We identified field notes describing 12 species occupying mountain glaciers and snowfields (Table 1). Travel by flying over glaciers and snowfields was noted for 13 species, and ground travel was only observed for White-tailed Ptarmigan (Lagopus leucura). Common Ravens were observed sliding down snowfields on their breasts. Feeding on glaciers and snowfields was noted for five species with arthropods, conifer seeds, and ice worms noted as food sources (Table 1). Baird’s Sandpipers were the only species observed feeding on ice worms. Observations of predators feeding in glaciers and snowfields were rare. A Golden Eagle (Aquila chrysaetos) was observed feeding on something on a snowfield, and a Prairie Falcon (Falco mexicanus) was observed chasing American Pipits near a glacier. American Pipits (3 obs.) and a Yellow-rumped Warbler (Setophaga coronata, 1 obs.) carrying food were the only observations marked with a breeding code. eBird users described hearing, but not seeing, five species: Clark’s Nutcracker (2 obs.), Cassin’s finch (Haemorhous cassinii, 3 obs.), Dark-eyed Junco (1 obs.), Pine Grosbeak (Pinicola enucleator, 1 obs.), and Yellow-rumped Warbler (1 obs.).
Community-science versus traditional field surveys of avian biodiversity
Field notes documented species occupying glaciers and snowfields that matched previous observations: 7 of 19 species compiled in a global review (Rosvold 2016) and all 7 species documented by camera trapping at Mount Rainier, Washington (Hotaling et al., 2022, personal communication). We identified four new glacier- or snowfield-associated species using community-science data: Baird’s Sandpiper, White-crowned Sparrow (Zonotrichia leucophrys), Mountain Chickadee (Poecile gambeli), and Townsend’s Solitaire (Myadestes townsendi). Our community science data set was also comparable to a traditional transect-based survey approach for ice-adjacent habitat. Our data set shared 80% of species (CC = 0.81) with Boyle and Martin (2015). Moreover, eBird data outperformed traditional survey methods by capturing 11 more species. Observation frequency of each species was moderately correlated in both data sources (Fig. 4; Linear regression: R² = 0.35, F1,130 = 71.02, p < 0.001).
DISCUSSION
Mountain glaciers and snowfields support unique, specialized biodiversity but are critically threatened by climate change worldwide (Hotaling et al. 2020, Stibal et al. 2020). To date, vertebrate biodiversity on glaciers and snowfields has been understudied (Rosvold 2016), likely because of difficulty of access. In this study, we used community science to characterize avian diversity on glaciers and snowfields, highlighting the power of this type of data for improving understanding of hard-to-study ecosystems. Through community-science data, we identified 46 bird species associated with glaciers and snowfields including four species that had not been associated with these habitats previously. Given the limited geographic scope of our study, we expect that new cryosphere-associated species remain to be discovered via community science approaches.
Specialized avian biodiversity on mountain glaciers and snowfields
Our results demonstrate strong preference in Gray-crowned Rosy-finches and American Pipits for glacier and snowfield habitats, which aligns with prior studies (e.g., Rosvold 2016), including observations of Gray-crowned Rosy-finches nesting near glaciers (Johnson 1965). Other species more common in these habitats were more surprising, notably American Kestrels and Rock Wrens. American Kestrels have also been observed via camera trapping (Hotaling et al., 2022, personal communication), and are known to occupy a broad range of open habitats, including alpine tundra. Rock Wrens have not been documented using glaciers or snowfields but are known to be high-elevation tolerant (Benedict et al. 2021). It is possible that these unexpected species represent recent changes in habitat use. Collectively, our results suggest that a small, overlooked community of ice-specializing birds may exist, likely with region-specific taxa (e.g., White-winged Diuca Finches nesting in crevasses of Andean glaciers, Hardy and Hardy 2008).
Perennial snow and ice as a resource subsidy for mountain birds
Glaciers and snowfields capture drifting material like arthropods and seeds, and in some regions, hold in situ nutrient sources like ice worms (Hotaling et al. 2017), thereby providing easily obtainable food sources for alpine birds (Edwards and Banko 1976, Mann et al. 1980, Crawford and Edwards 1986, Antor 1995, Rosvold 2016). Glaciers and snowfields appear to serve as a nutrient supplement during two energetically demanding life-cycle periods: breeding and migration (Hotaling et al. 2020). Most of the species observed foraging by eBird users are alpine breeders, so glaciers and snowfields likely hold important nutrient subsidies early in the breeding season (June) when snow cover reaches its peak (Hotaling et al. 2020). One species, Baird’s Sandpiper, was only recorded as a migrant in July and August. We captured the first observation of Baird’s Sandpipers feeding on ice worms, adding to six bird species previously recorded (Hotaling et al. 2020). The use of glaciers and snowfields by higher trophic level species remains poorly understood (Rosvold 2016), but we identified some evidence of hunting by birds of prey. Hunting habitat choice may explain apparent preference by American Kestrels for glaciers and snowfields as the surface may provide less escape cover for prey species, instead providing a white background on which they are easier to target.
North American avian biodiversity imperiled by glacier melt
Across species and space, a general rule of the biodiversity impacts of glacier recession on ice-linked species emerges: as glaciers decline, the abundance of habitat generalists increase and specialists decline (Cauvy-Fraunié and Dangles 2019). American Pipits showed the second strongest preference for glaciers and snowfields in our study, often using the ice and snow for foraging. In a well-studied population in Wyoming, up to 18% of foraging trips were made to snowfields during the breeding season (Hendricks 1987, Hendricks and Verbeek 2020). However, American Pipits also seem to prefer foraging on south-facing slopes, which are typically more snow free (Norvell and Creighton 1990). It is possible that glacier melt will negatively impact American Pipit populations by reducing food availability during breeding, but their use of diverse feeding habitat suggests they may be able to supplement their diet in adjacent habitats.
Among North American species, Gray-crowned Rosy-finches, however, may be most at risk from glacier melt. In addition to a strong preference for foraging on glaciers and snowfields, Gray-crowned Rosy-finch are the highest-nesting songbird in North America (MacDougall-Shackleton et al. 2020). Species distribution modeling indicates that they are extremely vulnerable to projected climate change in the next half century because of their narrow, high-altitude breeding niche (Conrad 2015). It seems likely that loss of foraging habitat will be a major factor, particularly during breeding, contributing to population declines. Our evidence and previous observations (Rosvold 2016, Hotaling et al. 2020) suggest Gray-crowned Rosy-finches frequently feed on arthropods and ice worms in glaciers and snowfields, and that invertebrate feeding increases during breeding (MacDougall-Shackleton et al. 2020). Whether Gray-crowned Rosy-finches can adjust their diet to compensate for changing food resources is unclear.
Leveraging community science to study enigmatic ecosystems
Although potential for sampling bias is ever present in eBird data (Sullivan et al. 2009, Boyle and Martin 2015), we were able to extract a large sample size to characterize avian communities in difficult-to-study mountain habitats. We found species that matched previous observations from glaciers and snowfields, and our data set was comparable in terms of community composition and species richness to traditional approaches. However, to generate comparable data sets and mitigate sampling bias, it is important to normalize data, perhaps by using resampling methods like the bootstrapping approach we applied. In a similar study focused on a rare species, Tricolored Blackbird (Agelaius tricolor), undersampling of absence data was used to compensate for sampling bias and improve species distribution model performance (Robinson et al. 2018). In our view, community science data represent a highly valuable resource for studying uncommon species and habitats as long as appropriate measures are taken to address sampling bias.
We used eBird field notes to extend our data set to include behavioral insight. eBird users more frequently described behavior in field notes rather than breeding codes. Considering the vast eBird framework, a more automated approach to processing field notes could provide equally powerful insights for behavior as the existing observation-focused framework. Such automation could be achieved by applying natural language processing (NLP; Hirschberg and Manning 2015), which uses computational methods to interpret written language. NLP has been successfully applied in other fields (e.g., health outcomes from medical records; Velupillai et al. 2018), and pre-trained models have emerged recently, which simplify the application of NLP methods (Qiu et al. 2020).
The most difficult challenge to using community science data for our research goals was overcoming spatial uncertainty in eBird data. eBird checklists with a shorter travel distance produce more precise location data, and thus higher spatial certainty (Steen et al. 2019). Therefore, we faced a common trade-off between spatial resolution and data abundance for community science data when rare species or understudied habitats are considered: more stringent filtering by travel distance meant a smaller data set (Van Eupen et al. 2021). Additionally, user error when submitting eBird checklist locations and/or inclusion of species heard or seen in other areas may falsely inflate records. At present, eBird data is a valuable starting point for studying difficult to access areas or assessing large-scale (e.g., global) patterns, but traditional surveys are still required when fine-scale quantification is needed. Perhaps an ideal approach integrates the two data sources, leveraging the advantages of each (e.g., Howell et al. 2022).
Beyond birds, similar community science analyses of rare and under-studied ecosystems can likely be conducted for other taxa. Many platforms aggregate data from non-avian taxa, but some have structural features that limit data quality and value. For example, iNaturalist is a popular online platform that accepts reports of any organism (or abiotic evidence of organisms, e.g., tracks). The broad scope of iNaturalist allows a remarkable range of creative studies, like testing for increased use of urban habitat by mammals during the COVID-19 pandemic lockdown (Vardi et al. 2021). However, projects using iNaturalist data are limited by presence-only data. Because eBird checklists have the option to be marked as complete, absence data and abundance estimates can be inferred, thereby leading to conclusions that are more robust and resistant to sampling bias (Robinson et al. 2018). Additionally, iNaturalist uses a community reviewing system, with observations deemed “research grade” when enough users have confirmed the identification. The community reviewing system includes rewards for number of reviews, creating a potential incentive for maximizing quantity over quality when reviewing identifications. For instance, research grade observations have been shown to not have higher taxonomic accuracy (Hochmair et al. 2020), suggesting a community review system is not sufficient for data validation. Adoption of a system similar to eBird’s—where rare species are captured by an automatic filter and reviewed by pre-established experts—could improve this issue.
CONCLUSION
Community science data clearly provide a valuable resource for quantifying biodiversity and engaging non-professional scientists in research. In this study, we described a framework for using eBird to assess avian biodiversity in understudied mountain cryosphere habitats that are imperiled by climate change. While supporting lower avian diversity than nearby, ice-adjacent habitats, glaciers and snowfields appear to provide key foraging grounds for many birds, including a few specialized species. As climate change proceeds, we expect that a loss of foraging grounds may negatively impact species like the Gray-crowned Rosy-finch. As community science platforms grow and mature, we expect their research value will continue to improve and eventually allow for robust temporal assessments of biodiversity change as year-over-year records accumulate. Collectively, our study highlights the power of community science data for modern ecological research, including the additive power that user reported field notes can add to observational data. We hope the professional research community continues to embrace community science data as we seek to realize their full potential for monitoring global biodiversity amidst an array of anthropogenic threats.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
AUTHOR CONTRIBUTIONS
W.B. and P.W. conceived of the study. W.B., J.B., N.P., P.W., and S.H. gave input on study design and data analysis. W.B. collected and analyzed the data. W.B. wrote the initial manuscript draft and all authors made considerable contributions to its development.
ACKNOWLEDGMENTS
We thank eBird developers, users, and reviewers for their contributions to the eBird project, and those who publish publicly available coding help. We also thank an anonymous reviewer and Editor-in-Chief Reed Bowman for their valuable comments on our manuscript.
DATA AVAILABILITY
Data is available for download on eBird.org. All code used for this study is publicly available on GitHub (https://github.com/willbrooks0/eBird-glacier.git).
LITERATURE CITED
Antor, R. J. 1995. The importance of arthropod fallout on snow patches for the foraging of high-alpine birds. Journal of Avian Biology 26:81-85. https://doi.org/10.2307/3677216
Benedict, L., N. B. Warning, N. A. Najar, S. G. Pitt, P. E. Lowther, D. E. Kroodsma, and G. H. Farley. 2021. Rock Wren (Salpinctes obsoletus), version 2.0. In P. G. Rodewald and B. K. Keeney, editors. Birds of the World, Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.rocwre.02
Bliss, A., R. Hock, and V. Radić. 2014. Global response of glacier runoff to twenty-first century climate change. Journal of Geophysical Research: Earth Surface 119:717-730. https://doi.org/10.1002/2013JF002931
Bolch, T., A. Kulkarni, A. Kääb, C. Huggel, F. Paul, J. G. Cogley, H. Frey, J. S. Kargel, K. Fujita, M. Scheel, S. Bajracharya, and M. Stoffel. 2012. The state and fate of Himalayan glaciers. Science 336(6079):310-314. https://doi.org/10.1126/science.1215828
Boyle, W. A., and K. Martin. 2015. The conservation value of high elevation habitats to North American migrant birds. Biological Conservation 192:461-476. https://doi.org/10.1016/j.biocon.2015.10.008
Callaghan, C. T., and D. E. Gawlik. 2015. Efficacy of eBird data as an aid in conservation planning and monitoring. Journal of Field Ornithology 86:298-304. https://doi.org/10.1111/jofo.12121
Callaghan, C. T., M. Lyons, J. M. Martin, R. E. Major, and R. T. Kingsford. 2017. Assessing the reliability of avian biodiversity measures of urban greenspaces using eBird citizen science data. Avian Conservation and Ecology 12(2):12. https://doi.org/10.5751/ACE-01104-120212
Camfield, A. F., S. F. Pearson, and K. Martin. 2010. Life history variation between high and low elevation subspecies of Horned Larks Eremophila spp. Journal of Avian Biology 41:273-281. https://doi.org/10.1111/j.1600-048X.2009.04816.x
Cauvy-Fraunié, S., and O. Dangles. 2019. A global synthesis of biodiversity responses to glacier retreat. Nature Ecology and Evolution 3:1675-1685. https://doi.org/10.1038/s41559-019-1042-8
Chandler, M., L. See, K. Copas, A. M. Z. Bonde, B. C. López, F. Danielsen, J. K. Legind, S. Masinde, A. J. Miller-Rushing, G. Newman, A. Rosemartin, and E. Turak. 2017. Contribution of citizen science towards international biodiversity monitoring. Biological Conservation 213:280-294. https://doi.org/10.1016/j.biocon.2016.09.004
Conrad, C. C., and K. G. Hilchey. 2011. A review of citizen science and community-based environmental monitoring: issues and opportunities. Environmental Monitoring and Assessment 176:273-291. https://doi.org/10.1007/s10661-010-1582-5
Conrad, E. C. 2015. Using species distribution models to quantify climate change impacts on the Rosy-Finch superspecies: an alpine obligate. Thesis. The University of Utah, Salt Lake City, Utah, USA. https://www.proquest.com/docview/1758252469/abstract/331A0638176E46C1PQ/1
Crawford, R. L., and J. S. Edwards. 1986. Ballooning spiders as a component of arthropod fallout on snowfields of Mount Rainier, Washington, U.S.A. Arctic and Alpine Research 18:429-437. https://doi.org/10.2307/1551092
de Sherbinin, A., A. Bowser, T. R. Chuang, C. Cooper, F. Danielsen, R. Edmunds, P. Elias, E. Faustman, C. Hultquist, R. Mondardini, I. Popescu, A. Shonowo, and K. Sivakumar. 2021. The critical importance of citizen science data. Frontiers in Climate 3. https://www.frontiersin.org/article/10.3389/fclim.2021.650760
Edwards, J. S., and P. C. Banko. 1976. Arthropod fallout and nutrient transport: a quantitative study of Alaskan snowpatches. Arctic and Alpine Research 8:237-245. https://doi.org/10.2307/1550470
Freeman, B. G., M. Strimas-Mackey, and E. T. Miller. 2022. Interspecific competition limits bird species’ ranges in tropical mountains. Science 377(6604):416-420. https://doi.org/10.1126/science.abl7242
Goodman, D. 1971. Ecological investigations of ice worms on casement glacier, southeastern Alaska (No. 39; Institute of Polar Studies). Ohio State University Research Foundation, Columbus, Ohio, USA. https://doi.org/10.2172/4569201
Hardy, D. R., and S. P. Hardy. 2008. White-winged Diuca Finch (Diuca speculifera) nesting on Quelccaya Ice Cap, Perú. Wilson Journal of Ornithology 120:613-617. https://doi.org/10.1676/06-165.1
Hendricks, P. 1987. Foraging patterns of Water Pipits (Anthus spinoletta) with nestlings. Canadian Journal of Zoology 65:1522-1529. https://doi.org/10.1139/z87-235
Hendricks, P., and N. A. Verbeek. 2020. American Pipit (Anthus rubescens), version 1.0. In S. M. Billerman, editor. Birds of the World, Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.amepip.01
Hirschberg, J., and C. D. Manning. 2015. Advances in natural language processing. Science 349:261-266. https://doi.org/10.1126/science.aaa8685
Hochmair, H. H., R. H. Scheffrahn, M. Basille, and M. Boone. 2020. Evaluating the data quality of iNaturalist termite records. PLoS ONE 15(5):e0226534. https://doi.org/10.1371/journal.pone.0226534
Hotaling, S., E. Hood, and T. L. Hamilton. 2017. Microbial ecology of mountain glacier ecosystems: biodiversity, ecological connections and implications of a warming climate. Environmental Microbiology 19:2935-2948. https://doi.org/10.1111/1462-2920.13766
Hotaling, S., S. Lutz, R. J. Dial, A. M. Anesio, L. G. Benning, A. G. Fountain, J. L. Kelley, J. McCutcheon, S. M. Skiles, N. Takeuchi, and T. L. Hamilton. 2021. Biological albedo reduction on ice sheets, glaciers, and snowfields. Earth-Science Reviews 220:103728. https://doi.org/10.1016/j.earscirev.2021.103728
Hotaling, S., P. H. Wimberger, J. L. Kelley, and H. E. Watts. 2020. Macroinvertebrates on glaciers: a key resource for terrestrial food webs? Ecology 101(4):e02947. https://doi.org/10.1002/ecy.2947
Howell, P. E., P. K. Devers, O. J. Robinson, and J. A. Royle. 2022. Leveraging community science data for population assessments during a pandemic. Ecological Applications 32:e2529. https://doi.org/10.1002/eap.2529
Hutcheson, K. 1970. A test for comparing diversities based on the Shannon formula. Journal of Theoretical Biology 29:151-154. https://doi.org/10.1016/0022-5193(70)90124-4
Johnson, R. E. 1965. Reproductive activities of Rosy Finches, with special reference to Montana. Auk 82(2):190-205. https://doi.org/10.2307/4082932
Johnston, A., W. M. Hochachka, M. E. Strimas-Mackey, V. Ruiz Gutierrez, O. J. Robinson, E. T. Miller, T. Auer, S. T. Kelling, and D. Fink. 2021. Analytical guidelines to increase the value of community science data: an example using eBird data to estimate species distributions. Diversity and Distributions 27:1265-1277. https://doi.org/10.1111/ddi.13271
Kosmala, M., A. Wiggins, A. Swanson, and B. Simmons. 2016. Assessing data quality in citizen science. Frontiers in Ecology and the Environment 14:551-560. https://doi.org/10.1002/fee.1436
MacDougall-Shackleton, S. A., R. E. Johnson, and T. P. Hahn. 2020. Gray-crowned Rosy-finch (Leucosticte tephrocotis), version 1.0. In A. F. Poole and F. B. Gill, editors. Birds of the World, Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bow.gcrfin.01
Mann, D. H., J. S. Edwards, and R. I. Gara. 1980. Diel activity patterns in snowfield foraging invertebrates on Mount Rainier, Washington. Arctic and Alpine Research 12:359-368. https://doi.org/10.2307/1550722
Moore, R. D., S. W. Fleming, B. Menounos, R. Wheate, A. Fountain, K. Stahl, K. Holm, and M. Jakob. 2009. Glacier change in western North America: influences on hydrology, geomorphic hazards and water quality. Hydrological Processes 23:42-61. https://doi.org/10.1002/hyp.7162
Natural Resources Canada, Instituto Nacional de Estadística y Geografía, U.S. Geological Survey, and North American Commission for Environmental Cooperation. 2004. North America Glaciers and Sea Ice. https://www.sciencebase.gov/catalog/item/4fb69683e4b03ad19d64b454
Norvell, J. R., and P. D. Creighton. 1990. Foraging of Horned Larks and Water Pipits in alpine communities. Journal of Field Ornithology 61:434-440.
Peter, M., T. Diekötter, T. Höffler, and K. Kremer. 2021. Biodiversity citizen science: outcomes for the participating citizens. People and Nature 3:294-311. https://doi.org/10.1002/pan3.10193
QGIS Development Team. 2022. QGIS Geographic Information System. QGIS Association.
Qiu, X., T. Sun, Y. Xu, Y. Shao, N. Dai, and X. Huang. 2020. Pre-trained models for natural language processing: a survey. Science China Technological Sciences 63:1872-1897. https://doi.org/10.1007/s11431-020-1647-3
Robinson, O. J., V. Ruiz-Gutierrez, and D. Fink. 2018. Correcting for bias in distribution modelling for rare species using citizen science data. Diversity and Distributions 24:460-472. https://doi.org/10.1111/ddi.12698
Rosvold, J. 2016. Perennial ice and snow-covered land as important ecosystems for birds and mammals. Journal of Biogeography 43:3-12. https://doi.org/10.1111/jbi.12609
RStudio Team. 2020. RStudio: Integrated Development for R. RStudio.
Scridel, D., M. Brambilla, K. Martin, A. Lehikoinen, A. Iemma, A. Matteo, S. Jähnig, E. Caprio, G. Bogliani, P. Pedrini, A. Rolando, R. Arlettaz, and D. Chamberlain. 2018. A review and meta-analysis of the effects of climate change on Holarctic mountain and upland bird populations. Ibis 160:489-515. https://doi.org/10.1111/ibi.12585
Shain, D. H., T. A. Mason, A. H. Farrell, and L. A. Michalewicz. 2001. Distribution and behavior of ice worms (Mesenchytraeus solifugus) in south-central Alaska. Canadian Journal of Zoology 79:1813-1821. https://doi.org/10.1139/z01-143
Shewey, J., and T. Blount. 2017. Birds of the Pacific Northwest. Timber, Portland, Oregon, USA.
Silvertown, J. 2009. A new dawn for citizen science. Trends in Ecology & Evolution 24:467-471. https://doi.org/10.1016/j.tree.2009.03.017
Slager, D. L. 2020. Seasonal and directional dispersal behavior in an ongoing dove invasion. Journal of Avian Biology 51(2):e02332. https://doi.org/10.1111/jav.02332
Sorensen, T. A. 1948. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biologiske Skrifter 5:1-34.
Steen, V. A., C. S. Elphick, and M. W. Tingley. 2019. An evaluation of stringent filtering to improve species distribution models from citizen science data. Diversity and Distributions 25:1857-1869. https://doi.org/10.1111/ddi.12985
Stibal, M., J. A. Bradley, A. Edwards, S. Hotaling, K. Zawierucha, J. Rosvold, S. Lutz, K. A. Cameron, J. A. Mikucki, T. J. Kohler, M. Šabacká, and A. M. Anesio. 2020. Glacial ecosystems are essential to understanding biodiversity responses to glacier retreat. Nature Ecology & Evolution 4:686-687. https://doi.org/10.1038/s41559-020-1163-0
Strimas-Mackey, M., E. Miller, and W. Hochachka. 2018. auk: EBird Data Extraction and Processing with AWK. R Package Version.
Sullivan, B. L., C. L. Wood, M. J. Iliff, R. E. Bonney, D. Fink, and S. Kelling. 2009. eBird: A citizen-based bird observation network in the biological sciences. Biological Conservation 142:2282-2292. https://doi.org/10.1016/j.biocon.2009.05.006
U.S. Geological Survey, and Natural Resources Canada. 2007. North America Elevation 1-Kilometer Resolution GRID. https://www.sciencebase.gov/catalog/item/4fb5495ee4b04cb937751d6d
Van Eupen, C., D. Maes, M. Herremans, K. R. R. Swinnen, B. Somers, and S. Luca. 2021. The impact of data quality filtering of opportunistic citizen science data on species distribution model performance. Ecological Modelling 444:109453. https://doi.org/10.1016/j.ecolmodel.2021.109453
Vardi, R., O. Berger-Tal, and U. Roll. 2021. iNaturalist insights illuminate COVID-19 effects on large mammals in urban centers. Biological Conservation 254:108953. https://doi.org/10.1016/j.biocon.2021.108953
Velupillai, S., H. Suominen, M. Liakata, A. Roberts, A. D. Shah, K. Morley, D. Osborn, J. Hayes, R. Stewart, J. Downs, W. Chapman, and R. Dutta. 2018. Using clinical natural language processing for health outcomes research: overview and actionable suggestions for future advances. Journal of Biomedical Informatics 88:11-19. https://doi.org/10.1016/j.jbi.2018.10.005
Fig. 1
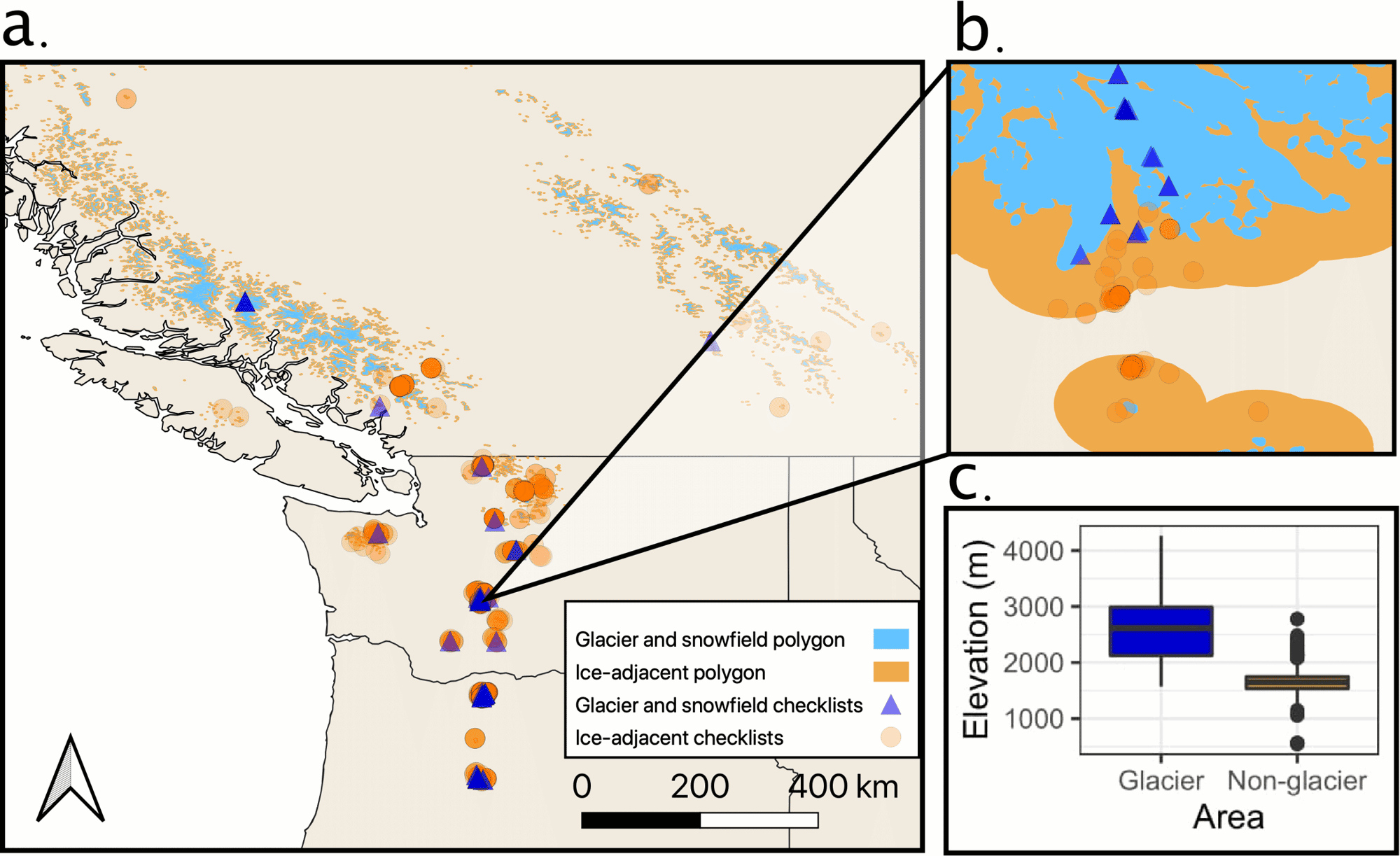
Fig. 1. (a) Locations of eBird checklists on glaciers and snowfield (blue triangles) versus ice-adjacent habitats (orange circles). (b) Inset of the area around one of our focal hotspots—Panorama Point in Mount Rainier National Park, WA—showing how checklists were identified with habitat polygons. (c) Elevation of eBird checklists in our data set by habitat type.
Fig. 2
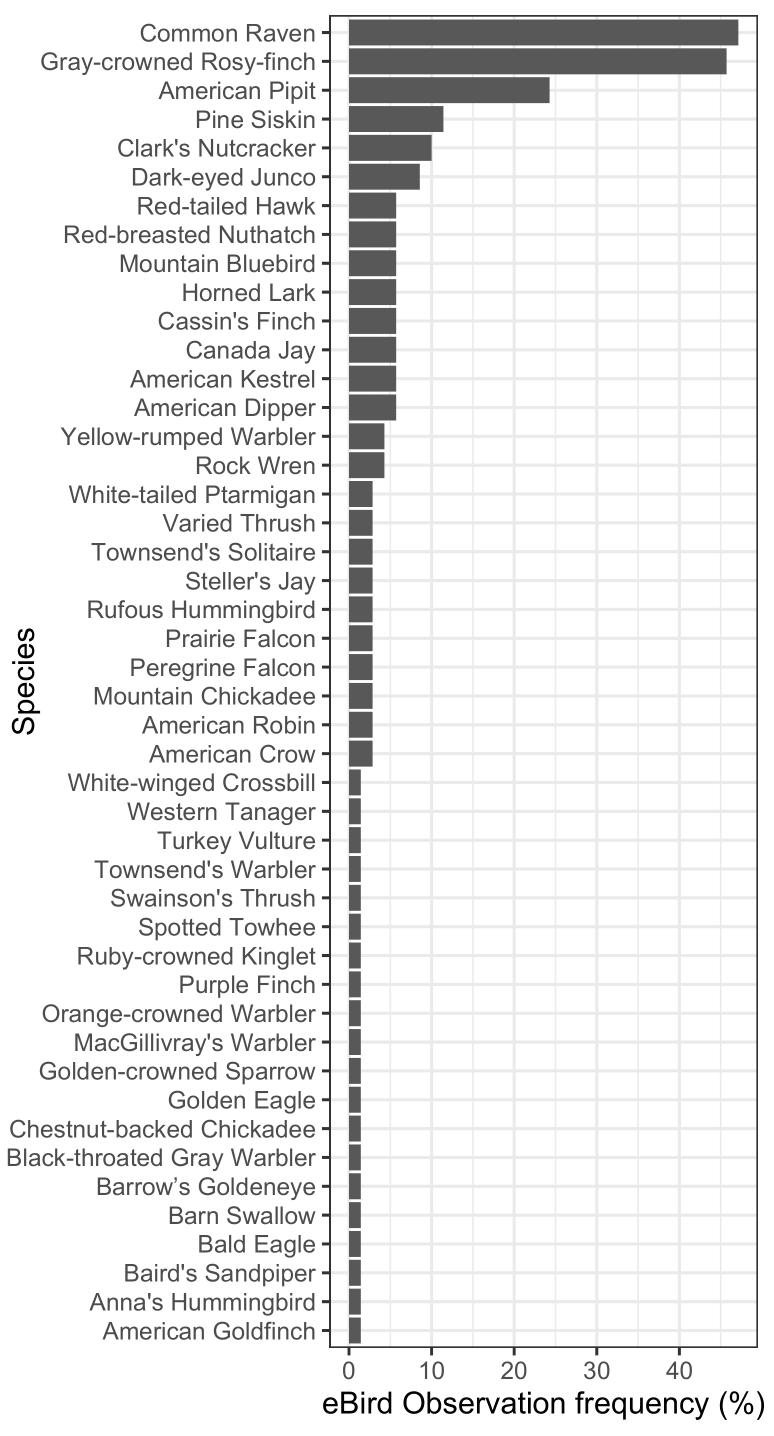
Fig. 2. Observation frequency for all birds included in our data set for glacier and snowfield habitat.
Fig. 3
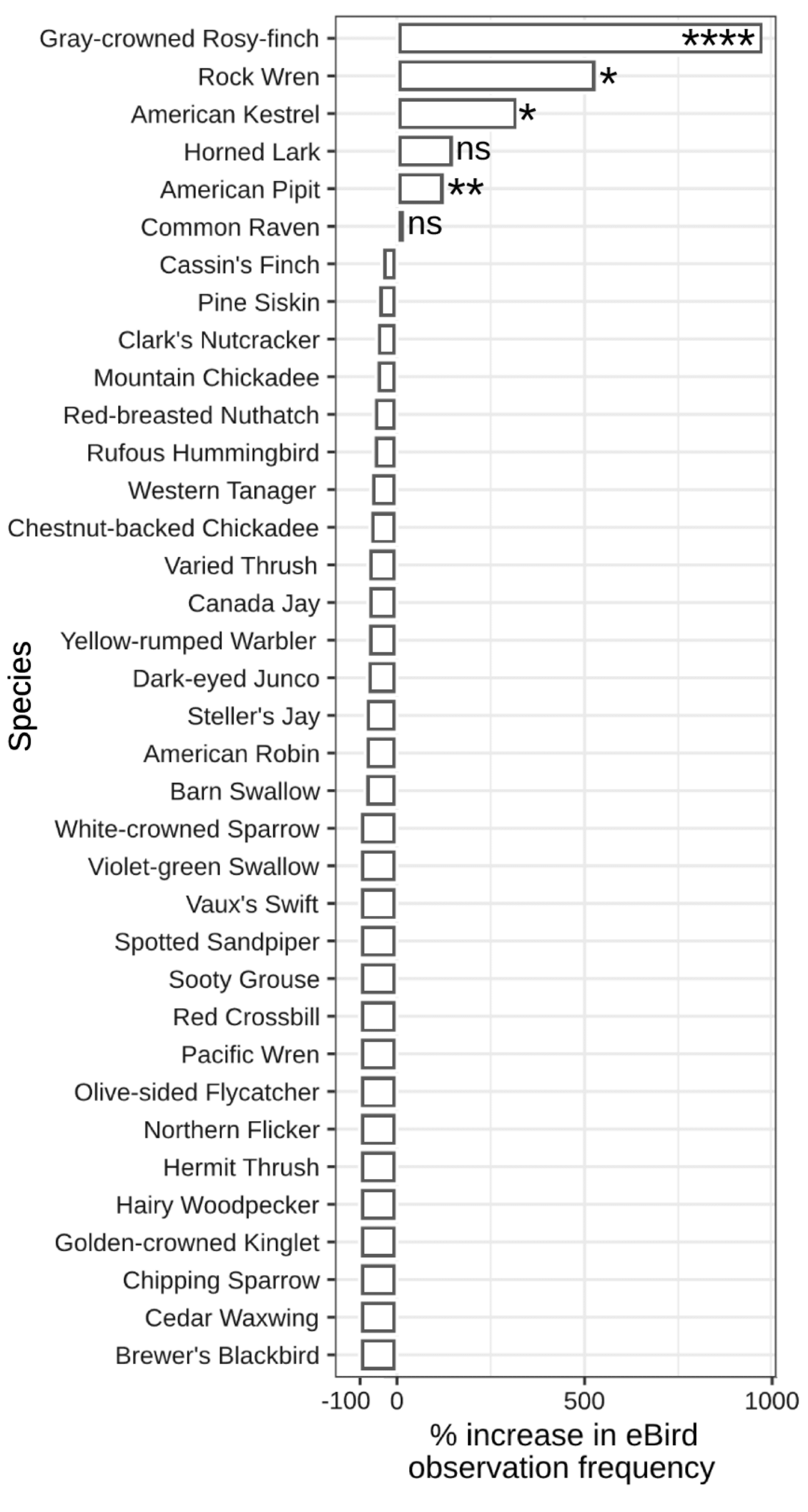
Fig. 3. The difference in eBird observation frequency (%) for each species in glaciers and snowfield habitat relative to ice-adjacent habitat. Statistical significance from chi-square tests indicate species that are more common in glaciers and snowfields at p < 0.0001 (****), p < 0.001 (***), p < 0.01 (**), and p < 0.05 (*). Non-significant values are indicated with “ns.” Only observation frequency differences ≥ 0.03 are shown.
Fig. 4
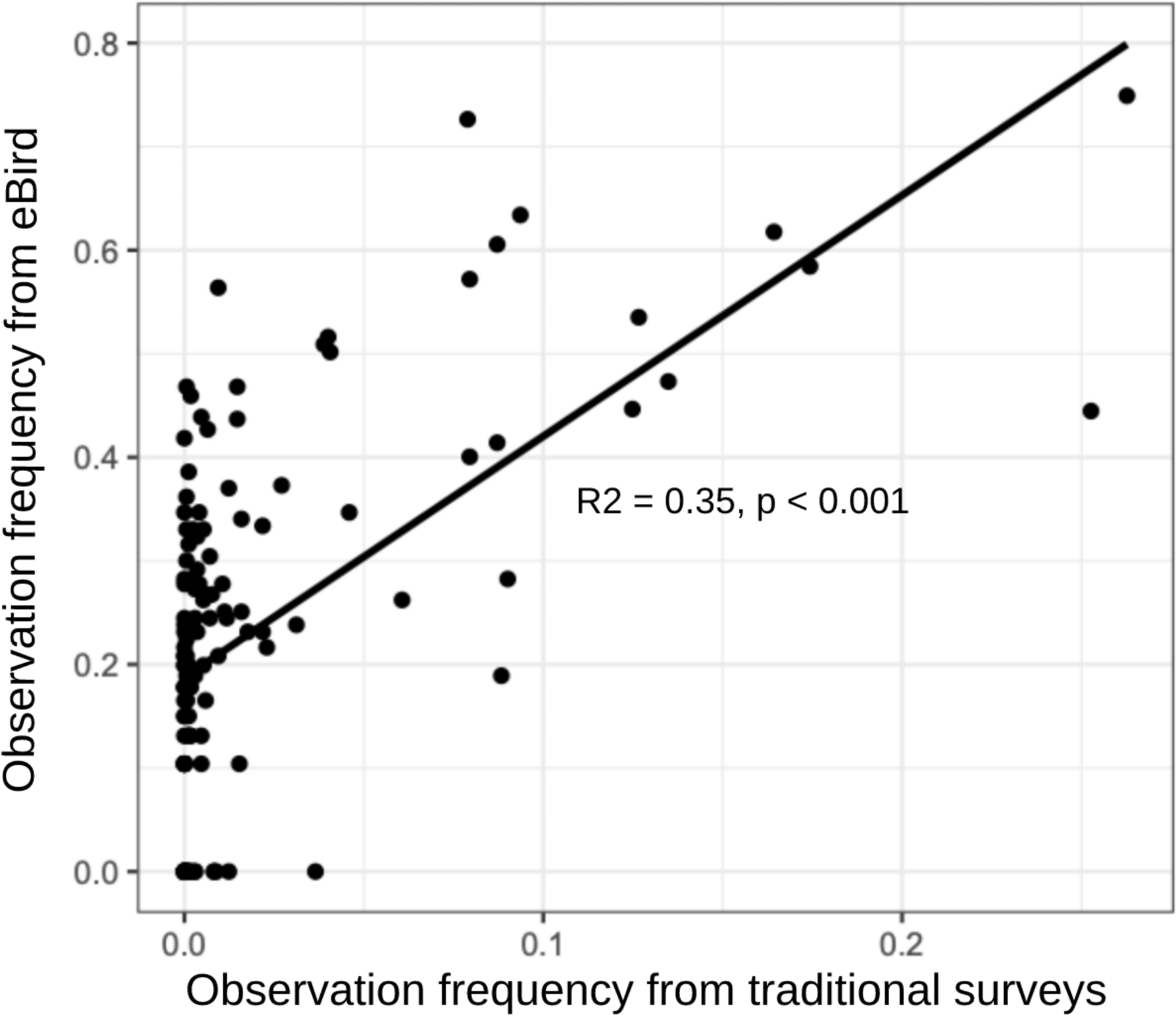
Fig. 4. Species observation frequencies for community science data (eBird) versus traditional surveys (Boyle and Martin 2015). Each point represents a single species.
Table 1
Table 1. Habitat and behavior descriptions extracted from eBird field notes. Birds flying over glaciers and snowfields were counted separately from birds standing in either habitat. The number of observations for each food source is shown in parentheses.
| Species | Habitat | Flying | Foraging | Food source | ||
| Glacier | Snowfield | Total | ||||
| Gray-crowned Rosy-finch (Leucosticte tephrocotis) |
6 | 28 | 34 | 12 | Arthropods (5), conifer seeds (1) | |
| American Pipit (Anthus rubescens) |
4 | 7 | 11 | 1 | 5 | Arthropods (2) |
| Horned Lark (Eremophila alpestris) |
1 | 7 | 8 | 3 | Arthropods (2) | |
| Baird’s Sandpiper (Calidris bairdii) |
5 | 5 | 1 | 5 | Arthropods (1), ice worms (1) | |
| Common Raven (Corvus corax) |
4 | 4 | ||||
| White-tailed Ptarmigan (Lagopus leucura) |
3 | 3 | ||||
| White-crowned Sparrow (Zonotrichia leucophrys) |
1 | 1 | ||||
| Townsend’s Solitaire (Myadestes townsendi) |
1 | 1 | ||||
| Mountain Chickadee (Poecile gambeli) |
1 | 1 | ||||
| Mountain Bluebird (Sialia currucoides) |
1 | 1 | ||||
| Golden Eagle (Aquila chrysaetos) |
1 | 1 | 2 | 1 | ||
| American Robin (Turdus migratorius) |
1 | 1 | 1 | |||
| Prairie Falcon (Falco mexicanus) |
2 | |||||
| Vaux’s Swift (Chaetura vauxi) |
2 | |||||
| Pine Siskin (Spinus pinus) |
2 | |||||
| Hermit Warbler (Setophaga occidentalis) |
1 | |||||
| Red-tailed Hawk (Buteo jamaicensis) |
1 | |||||
| Long-billed Dowitcher (Limnodromus scolopaceus) |
1 | |||||
| Pine Grosbeak (Pinicola enucleator) |
1 | |||||
| Calliope Hummingbird (Selasphorus calliope) |
1 | |||||
| Cooper’s Hawk (Accipiter cooperii) |
1 | |||||
| Bald Eagle (Haliaeetus leucocephalus) |
1 | |||||