







The following is the established format for referencing this article:
Robineau-Charette, G., D. M. Whitaker, and I. G. Warkentin. 2023. Change in altitudinal distribution of Newfoundland Gray-cheeked Thrush (Catharus minimus minimus) revealed through historical stop-level breeding bird survey data. Journal of Field Ornithology 94(1):4.ABSTRACT
Breeding bird survey (BBS) data are typically analyzed at the route level in which the average or combined total number of individuals across the 50 stops is treated as a single observation. However, analysis at the stop level may provide information on finer-scale habitat affiliations of species within the broader landscape, as well as changes in species distribution over time. Though once abundant, the Newfoundland Gray-cheeked Thrush (Catharus minimus minimus) underwent a dramatic population collapse across most of the island during the 1980s and 1990s and today is most common in montane forests. The reason for this collapse is not well understood but introduced North American red squirrels (Tamiasciurus hudsonicus), which colonized most forested areas on the island below ~400 m from the late 1970s through the mid-1990s, have been identified as a possible cause. To improve our understanding of changes in the elevational distribution of Gray-cheeked Thrushes, we compared stop-level BBS data from two periods. Our historic period from 1974-1984 corresponds with the initial expansion phase of introduced red squirrels, whereas that from 2000-2015 reflects a period during which the colonization of suitable habitat by squirrels was complete. We used 17 routes (850 stops) where the thrushes had been observed and there were at least 3 surveys completed during each period. Surveys during the 1970s and 1980s indicated that Gray-cheeked Thrushes were abundant at all elevations sampled by the BBS (0-450 m), with higher incidence below 100 m and above 300 m; in contrast, recent surveys indicate that the species is now essentially absent below 350 m. This finding highlights the value of historical stop-level BBS data for reconstructing changes in species distributions and is consistent with the hypothesis that red squirrels have adversely affected Newfoundland Gray-cheeked Thrushes.
RESUMEN
Los datos de los muestreos de aves reproductoras (BBS, por sus siglas en inglés) suelen analizarse a nivel de ruta, en el que el número total medio o combinado de individuos a lo largo de las 50 paradas se trata como una única observación. Sin embargo, el análisis a nivel de parada puede proporcionar información sobre asociaciones de hábitat de las especies, a escala más fina dentro de un paisaje más amplio, así como cambios en la distribución de las especies a lo largo del tiempo. Aunque abundante en el pasado, el Zorzal de Cara Gris de Terranova (Catharus minimus minimus) sufrió un dramático colapso poblacional en la mayor parte de la isla durante las décadas de 1980 y 1990 y hoy es más común en los bosques montanos. La razón de este colapso no se conoce bien, pero la introducción de las ardillas rojas norteamericanas (Tamiasciurus hudsonicus), que colonizaron la mayoría de las zonas boscosas de la isla por debajo de ~400 m desde finales de los 70 hasta mediados de los 90, ha sido identificada como una posible causa. Para mejorar nuestra comprensión de los cambios en la distribución altitudinal de los Zorzales de Cara Gris, comparamos los datos del BBS a nivel de parada de dos periodos. Nuestro periodo histórico de 1974-1984 se corresponde con la fase inicial de expansión de las ardillas rojas introducidas, mientras que el de 2000-2015 refleja un periodo durante el cual la colonización del hábitat adecuado por las ardillas fue completa. Se utilizaron 17 rutas (850 paradas) en las que se habían observado los zorzales y se realizaron al menos 3 muestreos durante cada periodo. Las encuestas realizadas durante los años 70 y 80 indicaron que los Zorzales de Cara Gris eran abundantes en todas las elevaciones muestreadas por el BBS (0-450 m), con una mayor incidencia por debajo de 100 m y por encima de 300 m; por el contrario, las encuestas recientes indican que la especie está ahora esencialmente ausente por debajo de 350 m. Este hallazgo pone de relieve el valor de los datos históricos del BBS a nivel de parada para reconstruir los cambios en la distribución de las especies y es consistente con la hipótesis de que las ardillas rojas han afectado negativamente a los Zorzales de Cara Gris de Terranova.
INTRODUCTION
The breeding bird survey (BBS) was established in 1966 to generate continental-scale population monitoring data for a broad range of bird species but with a particular focus on songbirds (Downes et al. 2016, Hudson et al. 2017). Thousands of BBS routes are now surveyed along secondary roads across North America each year, with each route consisting of 50 stops evenly spaced at fixed locations along a 39.2 km (24.5 mile) route. During a survey, the observer records all birds seen and heard during a three-minute silent point count at each stop. Breeding bird survey data have traditionally been analyzed based on observations from the entire route considering the combined total or average number of individual species observed across all 50 stops (Betts et al. 2022). However, BBS data include finer-scale information at the individual stop level that may be of value for research and conservation because it reflects the responses of individual birds to local variation in the environment that ultimately affect the distribution or occurrence of species (Veech and Crist 2007, Niemuth et al. 2017). Concerns were raised about the relative value of digitizing historical stop-level BBS data given the amount of effort required and the fact that the high resolution of stop-level data is poorly matched to the large-scale monitoring focus of the BBS (O’Connor et al. 2000). Nevertheless, the advent of technological advances including large-scale satellite imagery, digital maps of landform and landcover, global positioning systems, and increased computing power have increased the potential to harness the finer-scale information in BBS data (Downes et al. 2016) and BBS data collected since 1997 have been digitized at the stop level (Hudson et al. 2017). Although the recent stop-level BBS data are useful for assessing contemporary species distributions (Stralberg et al. 2015), historical BBS data may contain novel information on the distribution of species that have declined or disappeared from an area, as well as on changes in species distributions as a result of environmental stressors (McElhone et al. 2011, Betts et al. 2022). Consequently, efforts are underway to digitize historical stop-level BBS data from the original field data sheets (Downes et al. 2016, Hudson et al. 2017). Analyses of these historical stop-level data may offer important retrospective information for species of conservation concern that cannot be obtained from any other datasets today and may elucidate patterns of change or even possible reasons for such change.
Gray-cheeked Thrushes (Catharus minimus) breed across the northern boreal forest from Alaska to Newfoundland and Labrador, typically occupying dense conifer and broadleaf thickets (Whitaker et al. 2020). Remoteness of most breeding areas and the species’ furtive behavior have hindered study, and as a result, it remains one of the least understood boreal forest songbirds (Whitaker et al. 2020). Two subspecies of Gray-cheeked Thrushes have been recognized (Fitzgerald et al. 2017, Whitaker et al. 2020); the Newfoundland subspecies (C. m. minimus), which occurs on the island of Newfoundland and along the north shore of the Gulf of St. Lawrence, and the Northern subspecies (C. m. aliciae), which is found from central Labrador westward through Alaska. Although populations of the northern subspecies are relatively stable (Smith et al. 2020), the Newfoundland subspecies has experienced a dramatic decline and in 2015 was listed as threatened under the provincial Endangered Species Act based primarily on route-level BBS data analyses suggesting a ~95% decline (Whitaker 2010, Smith et al. 2020). During the 1970s and early 1980s, the Gray-cheeked Thrush was an abundant and widespread species on the island. It was detected during 75% of BBS surveys (mean 6.11 thrushes per survey; 115 surveys of 25 routes; Smith et al. 2020) and nearly one-third of surveys observed 10 or more individuals, with a maximum of 38 during 1 survey. By contrast, since 1988, no BBS survey has recorded more than 3 Gray-cheeked Thrushes on Newfoundland, and from 2010-2019 this species was only detected during 9% of BBS surveys (mean 0.13 thrushes per survey; 197 surveys of 27 routes; Smith et al. 2020).
The cause(s) of the sudden decline of Gray-cheeked Thrushes on Newfoundland are still being assessed. Habitat loss or degradation through timber harvesting have the potential to impact populations of forest birds (Thompson et al. 1999, Lamarre and Tremblay 2021, Betts et al. 2022), but the conifer scrub and montane forest favored by Gray-cheeked Thrushes on the island is seldom cut, and research also suggests that these thrushes are frequently associated with regenerating clearcuts (Whitaker et al. 2015, McDermott 2021). Likewise, protected areas on the island, like national parks, have shown sharp reductions in numbers of Gray-cheeked Thrushes (Jacques Whitford Environment 1993). Introduced species have also been tied to species extinctions across a wide range of taxa (Bellard et al. 2016), and avian species have been particularly vulnerable to becoming imperiled or extirpated when mammalian predators have been introduced to islands (Doherty et al. 2016). North American red squirrels (Tamiasciurus hudsonicus), known to cause high nest failure rates in the closely related Bicknell’s Thrush (Catharus bicknelli, Wallace 1939, McFarland et al. 2008) were introduced to Newfoundland in 1963 and 1964 and subsequent translocations led them to become widespread and abundant across most of the island by the mid-1990s (Minty 1976, Payne 1976, Goudie 1978, Whitaker 2015). This apparent synchrony between the rapid spread of squirrels across Newfoundland and the sudden decline of the Gray-cheeked Thrush population, as well as the known impact of squirrels on the closely related Bicknell’s Thrush (McFarland et al. 2008), has led to the hypothesis that the two were linked (Whitaker et al. 2015). Though they have received limited study on Newfoundland, recent research indicated that red squirrels are most abundant at lower elevations, becoming rare above ~300 m and absent above ~500 m (McDermott et al. 2020). Conversely, Gray-cheeked Thrushes have remained relatively abundant in montane forest ecosystems (Whitaker et al. 2015, McDermott 2021). Finally, degradation or loss of winter habitat in South America could also be a stressor affecting Gray-cheeked Thrushes that breed on Newfoundland (Whitaker et al. 2018). However, if impacts during the non-breeding period have driven the decline of the breeding population on Newfoundland, this might be expected to have equally affected the species across all elevations on Newfoundland.
We sought to expand on previous route-level analyses of Gray-cheeked Thrush population status (Whitaker 2010) and localized surveys (Lamberton 1976a, b, Whitaker et al. 2015, McDermott 2021). We accomplished this by comparing stop-level BBS data from historical (1974-1984) and recent (2001-2015) periods to quantify the apparent change in the elevational distribution of Gray-cheeked Thrushes on Newfoundland. The historical data correspond with the initial expansion phase of introduced red squirrels, whereas by the recent period the colonization of suitable habitat by squirrels was complete (Whitaker 2015). Quantifying change could lead to improved understanding of the limiting factors and stressors hypothesized to have contributed to the population decline on Newfoundland. It has already been documented that this species was historically abundant at both low and high elevations but today is only common at high elevations and on some coastal islands (Lamberton 1976a, b, Whitaker 2010, Whitaker et al. 2015, McDermott 2021). However, it is not clear whether they used to be evenly distributed across all elevations or if they had some other distribution such as the bimodal pattern displayed by Bicknell’s Thrushes, which were historically common on coastal islands and in montane forests of Nova Scotia, New Brunswick, and Québec but did not occur at intermediate elevations (Townsend et al. 2020).
METHODS
Newfoundland is the largest island in the circumpolar boreal biome (> 108,000 km²) and falls within bird conservation region 8 (Boreal Softwood Shield; Environment Canada 2013). Terrain rises to > 800 meters above sea level (m.a.s.l.) in the Long Range Mountains of western Newfoundland (Fig. 1), and landcover is dominated by conifer and mixed boreal forest in a mosaic with extensive bogs and coastal and alpine barrens (Damman 1983). Coastal and montane forests are exposed to high winds, leading to extensive cover of stunted, dense conifer scrub (“krumholtz”), which is dominated by balsam fir (Abies balsamea) and black spruce (Picea mariana; Damman 1983).
The first 2 BBS routes on Newfoundland were surveyed in 1974, these plus 6 other routes were surveyed a total of 10 times during the 1970s. This was followed by an intensive survey effort from 1980-1984, when most of the BBS route network on the island was established and 23 routes were surveyed. However, BBS effort then dropped, and most routes were abandoned until a renewed effort was launched in the early 2000s. Since then, most of the original routes have been reactivated and some new routes have been established to give a new total of 25 routes on the island, and survey efforts have been more consistent (Pardieck et al. 2016, Smith et al. 2020). Because of the limited BBS coverage from 1985 to ~2000, we restricted our analyses to two periods: (1) 1974-1984, when Gray-cheeked Thrushes were still common and widespread and before squirrels had widely expanded, and (2) 2001-2015, after the thrush population had declined and colonization of suitable habitat by squirrels was essentially complete (Whitaker 2015). We further restricted our analyses to include only those BBS routes where a Gray-cheeked Thrush had been observed at least on one occasion during either period, and which had been surveyed at least three times during each period. Stop-level counts of Gray-cheeked Thrushes for each of these routes, each year in our two focal periods, along with the geographic coordinates for each stop, were obtained through a special request to Environment and Climate Change Canada. We estimated the elevation of each stop from the national CanVec digital elevation geospatial database, available under the Government of Canada’s open government license https://open.canada.ca/en. Historical land cover data were not available for our study area.
We analyzed the BBS data using generalized additive models (GAMs; Zuur et al. 2009), which allowed the fitting of smoothed, non-linear relationships between continuous response and explanatory variables. Models were fit in R using the package mgcv and included time period (a factor) and elevation (fit by time period) as explanatory variables (R Core Team 2017, Wood 2021). We also explored analyses using generalized additive mixed models (GAMMs; Zuur et al. 2009) with route included as a random effect but model fit was poor. Closer examination revealed that the range of elevations among the 50 stops along each route was limited (Fig. 2), so route was a strong predictor of elevation (F16,833 = 194, p < 0.001, r²adj = 0.784) and acted as a proxy for elevation in the mixed models. Because route and elevation were confounded and could not be included in the same model, we reverted to the simpler fixed effects model (i.e., GAM); interpreting this fixed effects model was based on the assumption that the limited sampling of high elevation routes was nonetheless representative of high elevation locations. Our first model assessed the relationship between the presence/absence of a thrush during each time period and elevation, and used a binomial error distribution for presence/absence data (0/1) and a clog-log link function, which is more effective for datasets with a disproportionate number of zeros (Zuur et al. 2009). In the second analysis, we used the actual count of thrushes at each point as a function of elevation, again with time period as a main effect but using a negative binomial error distribution with a log link function, as suggested for over-dispersed data (Zuur et al. 2009). For the smoothed term in both models (i.e., elevation) we specified a value of k = 4; k is akin to the maximum number of knots (turning points) in the curve and constrains the maximum effective degrees of freedom (EDF; i.e., complexity) for the fitted curve to k - 1 (i.e., 3 in this instance; Wood 2017).
RESULTS
Seventeen BBS routes (total 850 stops) met our criteria for inclusion (Fig. 1). Though these BBS routes are widely distributed across Newfoundland, many follow roads connecting coastal communities and thus are strongly biased toward sampling lower elevations (Fig. 2). Just 52% of the island lies below 200 m.a.s.l. but 91% of stops fell below this level; similarly the highest elevation stop was just 437 m.a.s.l. but 7% of the island (7674 km²) exceeds 450 m.a.s.l. Sampling effort varied between periods, with routes being surveyed an average of 4.9 times each from 1974-1984 and 8.4 times each from 2001-2015, creating total samples of 4200 (1974-1984) and 7150 (2001-2015) stop-level counts. The average incidence was 7.20 Gray-cheeked Thrushes/route from 1974-1984 and 0.25 thrushes/route from 2001-2015. On a per-stop basis, the mean probability of observing a thrush was 0.152 from 1974-1984 (range 0-3, present at 527 of 4200 stops) versus a mean probability of 0.004 from 2001-2015 (range 0-2, present at 27 of 7150 stops), indicating a 97% reduction in thrush occurrence rate along routes between the 2 periods.
Our model of Gray-cheeked Thrush occurrence as a function of elevation explained 21.2% of the deviance in the data and indicated a significant reduction in occurrence rate along routes between periods (z = -17.22, P < 0.001). Elevation was a highly significant predictor of occurrence in both periods (1974-1984: EDF = 2.604, χ² = 26.46, P < 0.001; 2001-2015: EDF = 2.665, χ² = 23.66, P < 0.001). However, the relationship between presence/absence of Gray-cheeked Thrushes and elevation differed between periods (Fig. 3). During the 1974-1984 period, occurrence was highest at high elevations, lowest at mid elevations, and intermediate along the coast (Fig. 3), whereas during the 2001-2015 period, thrush occurrence was near zero at all but the highest elevations. Confidence limits around these lines of best fit increased with elevation because of the smaller number of BBS stops at higher elevations.
Incorporating actual counts of Gray-cheeked Thrushes detected per BBS stop produced a stronger model that explained 30.8% of the deviance in the data. Otherwise, this second model had almost identical results to our model using thrush occurrence as the response variable. Again, we found that counts of thrushes were significantly lower during 2001-2015 than during 1974-1984 (z = -18.33, P < 0.001). Likewise, elevation was a highly significant predictor of presence in both periods (1974-1984: EDF = 2.508, χ² = 21.74, P < 0.001; 2001-2015: EDF = 2.736, χ² = 22.26, P < 0.001) and showed the same patterns of change in distribution across elevations as did our occurrence model (Fig. 3).
DISCUSSION
Analyses of BBS data at the level of individual stops yielded valuable new information about changes to the historical distribution of the population of Gray-cheeked Thrushes on Newfoundland. Although the overall decline of this population had already been documented through route-level analysis of BBS data as well as local studies (Jacques Whitford 1993, Whitaker 2010, Fitzgerald et al. 2017), our finer-scale analysis provided evidence that there has been a contraction in the distribution of this imperiled island endemic subspecies. Specifically, our analysis showed that from 1974-1984 the thrushes were common below ~100 m and abundant at elevations above ~300 m but were less common at intermediate elevations. This is consistent with historical studies and the observations of birdwatchers as well as with the species reported affinity for dense coastal scrub (Peters and Burleigh 1951, Lamberton 1976a, b, Vasallo and Rice 1981, Mactavish 1988, Marshall 2000). In stark contrast, contemporary stop-level BBS data showed unequivocally that the species is now almost extirpated below 350 m, and overall evidence suggests that the majority of the contemporary population occurs in geographically limited high-elevation refugia. Although in this analysis the latter pattern is truncated by the limited elevation range of the BBS route network and a paucity of data above 200 m, it shows a striking similarity to findings of recent localized distributional studies that show a steady increase in the abundance of Gray-cheeked Thrushes at elevations from 350 to ~600 m.a.s.l. along the eastern slope of the Long Range Mountains (Whitaker et al. 2015, McDermott 2021). Similarly, in the 1970s, this species was abundant in lowland and mountainous portions of Gros Morne National Park. However, they had disappeared at low elevations by 1992 and now are only common in the montane forests of the park (Lamberton 1976a, Jacques Whitford 1993, Rae 2014; D. M. Whitaker, unpublished data).
Our results are consistent with the hypothesis that the Newfoundland population of Gray-cheeked Thrush was adversely affected by the introduction of red squirrels. A temporal correlation between the spread of red squirrels and the disappearance of thrushes has already been documented (Whitaker 2010, Whitaker et al. 2015). In 2016 and 2017, McDermott (2021) conducted colocated surveys for thrushes and squirrels across an elevation gradient from 75 to 608 m.a.s.l. and documented strong elevational segregation between the 2 species, with squirrels primarily occurring below 300 m.a.s.l. and thrushes almost entirely above 350 m.a.s.l. There is a striking similarity between this novel contemporary distribution of Gray-cheeked Thrushes on Newfoundland and that of the Bicknell’s Thrush in its Maritime Canada and U.S. breeding range, which is also thought to be strongly impacted by red squirrels at lower elevations (McFarland et al. 2008, Hill et al. 2019). Although montane areas appear to be serving as a contemporary refuge for Newfoundland Gray-cheeked Thrushes, it is unclear how long this situation will persist in the face of climate warming and land use change (McDermott et al. 2020, McDermott 2021).
Our study demonstrates the potential of stop-level BBS data for quantifying long-term changes in the distribution of species of conservation concern, though the power of such assessments will be greater where both historical and current land cover data are available. It also highlights a limitation of the BBS route network on Newfoundland because of the strong bias toward low elevations, with just 9% of stops along the 17 routes we examined occurring above 200 m.a.s.l. Other BBS routes have been established on the island, but these are also located in coastal, low elevation areas where most secondary roads are found (Fig. 1). An unfortunate consequence of this is that even though the BBS provides critical historical data documenting the disappearance of the once abundant coastal population of the Newfoundland Gray-cheeked Thrush, it is of little value for monitoring this species now that it is largely restricted to montane forests. Further, an analysis of the distribution of 32 common bird species across the eastern slope of the Long Range Mountains by McDermott et al. (2021) found that 12 species had higher rates of occupancy at higher elevations, while another 13 species showed decreased occupancy with increasing elevations. This indicates that as in other areas (Mizel et al. 2016, Ralston and DeLuca 2020), the boreal bird community on Newfoundland is strongly structured by elevation and thus highlights a shortcoming of the BBS for monitoring several widespread species such as Blackpoll Warbler (Setophaga striata) and Fox Sparrow (Passerella iliaca). However, < 14% of Newfoundland exceeds 375 m.a.s.l., and road access at higher elevations is limited and primarily occurs in the form of unpaved, poorly maintained resource roads that are ill-suited for long-term BBS routes. Thus, the solution may not lie in expansion of the BBS route network on the island but rather in more targeted monitoring approaches, such as breeding bird atlases and Mountain Birdwatch (Davidson et al. 2015, Torrenta 2021).
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
Sincere thanks to Kate Campbell (Environment and Climate Change Canada) for preparing and sharing stop-level BBS data for Gray-cheeked Thrush on Newfoundland. We also thank the hundreds of skilled volunteers in Canada who have participated in the BBS over the years and those who have served as provincial or territorial coordinators for the BBS. Breeding bird survey spatial data were made available as a result of collaboration between the Canadian Wildlife Service and the Boreal Avian Modeling Project (www.borealbirds.ca).
DATA AVAILABILITY
Available through Environment and Climate Change Canada.
LITERATURE CITED
Bellard, C., P. Cassey, and T. M. Blackburn. 2016. Alien species as a driver of recent extinctions. Biology Letters 12(2):20150623. http://dx.doi.org/10.1098/rsbl.2015.0623
Betts, M. G., Z. Yang, A. S. Hadley, A. C. Smith, J. S. Rousseau, J. M. Northrup, J. J. Nocera, N. Gorelick, and B. D. Gerber. 2022. Forest degradation drives widespread avian habitat and population declines. Nature Ecology and Evolution 6(6):709-719. https://doi.org/10.1038/s41559-022-01737-8
Damman, A. W. H. 1983. An ecological subdivision of the island of Newfoundland. Pages 163-206 in G. R. South, editor. Biogeography and ecology of the island of Newfoundland. Dr. W. Junk Publishers, The Hague, Netherlands.
Davidson, P. J. A., R. J. Cannings, A. R. Couturier, D. Lepage, and C. M. Di Corrado. 2015. Atlas of the breeding birds of British Columbia, 2008-2012. Birds Canada, Delta, British Columbia, Canada. http://www.birdatlas.bc.ca/
Doherty, T. S., A. S. Glen, D. G. Nimmo, E. G. Ritchie, and C. R. Dickman. 2016. Invasive predators and global biodiversity loss. Proceedings of the National Academy of Sciences 113(40):11261-11265. https://doi.org/10.1073/pnas.1602480113
Downes, C. M., M.-A. R. Hudson, A. C. Smith, and C. M. Francis. 2016. The breeding bird survey at 50: scientists and birders working together for bird conservation. Avian Conservation and Ecology 11(1):8. http://dx.doi.org/10.5751/ACE-00855-110108
Environment Canada. 2013. Bird conservation strategy for bird conservation region 8 and marine biogeographic units 10 and 12 in Newfoundland and Labrador: boreal softwood shield, scotian shelf and Newfoundland-Labrador shelves, and Gulf of St. Lawrence. Canadian Wildlife Service, Environment Canada, Sackville, New Brunswick, Canada. https://publications.gc.ca/site/eng/9.696546/publication.html
FitzGerald, A. M., D. M. Whitaker, J. Ralston, J. J. Kirchman, and I. G. Warkentin. 2017. Taxonomy and distribution of the imperilled Newfoundland Gray-cheeked Thrush, Catharus minimus. Avian Conservation and Ecology 12(1):10. https://doi.org/10.5751/ACE-00976-120110
Goudie, R. I. 1978. Red squirrels, Tamiasciurus hudsonicus, in the Salmonier River Valley, Newfoundland. Canadian Field-Naturalist 92:193-194.
Hill, J. M., J. D. Lloyd, K. P. McFarland, and C. C. Rimmer. 2019. Apparent survival of a range-restricted montane forest bird species is influenced by weather throughout the annual cycle. Avian Conservation and Ecology 14(2):16. https://doi.org/10.5751/ACE-01462-140216
Hudson, M.-A. R., C. M. Francis, K. J. Campbell, C. M. Downes, A. C. Smith, and K. L. Pardieck. 2017. The role of the North American Breeding Bird Survey in conservation. Condor 119(3):526-545. https://doi.org/10.1650/CONDOR-17-62.1
Jacques Whitford Environment. 1993. Avifaunal inventory, Gros Morne National Park. Parks Canada, Rocky Harbor, Newfoundland, Canada.
Lamarre, V., and J. A. Tremblay. 2021. Occupancy of the American Three-toed Woodpecker in a heavily-managed boreal forest of eastern Canada. Diversity 13(1):35. https://doi.org/10.3390/d13010035
Lamberton, R. D. 1976a. Avifaunal survey of Gros Morne National Park. Parks Canada, Rocky Harbor, Newfoundland, Canada.
Lamberton, R. D. 1976b. An avifaunal survey of L’-Anse-aux-Meadows National Historic Park, Newfoundland. Parks Canada, Rocky Harbor, Newfoundland, Canada.
Mactavish, B. 1988. The bird report: summer June 1-July 31, 1988. Bullbird, a Newfoundland Birdwatching Journal 3(4):1-9.
Marshall, J. T. 2000. The Gray-cheeked Thrush, Catharus minimus, and its New England subspecies, Bicknell’s Thrush, Catharus minimus bicknelli. Harvard University Nuttall Ornithological, Cambridge, Massachusetts, USA.
McDermott, J. P. B. 2021. Habitat associations of red squirrels (Tamiasciurus hudsonicus) and their impact on the Gray-cheeked Thrush (Catharus minimus) in western Newfoundland. Thesis. Memorial University of Newfoundland, St. John’s, Newfoundland and Labrador, Canada. https://research.library.mun.ca/15012/1/thesis.pdf
McDermott, J. P. B., I. G. Warkentin, and D. M. Whitaker. 2021. The influence of elevation on bird assemblages in the Long Range Mountains of western Newfoundland, including consideration of potential effects of introduced red squirrels. Unpublished report. Parks Canada, Rocky Harbor, Newfoundland, Canada.
McDermott, J. P. B., D. M. Whitaker, and I. G. Warkentin. 2020. Constraints on range expansion of introduced red squirrels (Tamiasciurus hudsonicus) in an island ecosystem. Canadian Journal of Forest Research 50(10):1064-1073. https://doi.org/10.1139/cjfr-2019-0369
McElhone, P. M., P. Bohall Wood, and D. K. Dawson. 2011. Effects of stop-level habitat change on Cerulean Warbler detections along breeding bird survey routes in the central Appalachians. Wilson Journal of Ornithology 123(4):699-708. http://dx.doi.org/10.1676/09-159.1
McFarland, K. P., C. C. Rimmer, S. J. K. Frey, S. D. Faccio, and B. B. Collins. 2008. Demography, ecology and conservation of Bicknell’s Thrush in Vermont, with a special focus on the Northeast Highlands. Technical Report 08-03. Vermont Center for Ecostudies, Norwich, Vermont, USA.
Minty, D. 1976. Red squirrels in Newfoundland. Osprey 7(1):19-24.
Mizel, J. D., J. H. Schmidt, C. L. McIntyre, and C. A. Roland. 2016. Rapidly shifting elevational distributions of passerine species parallel vegetation change in the subarctic. Ecosphere 7(3):e01264. https://doi.org/10.1002/ecs2.1264
Niemuth, N. D., M. E. Estey, S. P. Fields, B. Wangler, A. A. Bishop, P. J. Moore, R. C. Grosse, and A. J. Ryba. 2017. Developing spatial models to guide conservation of grassland birds in the U.S. Northern Great Plains. Condor 119(3):506-525. https://doi.org/10.1650/CONDOR-17-14.1
O’Connor, R. J., E. Dunn, D. H. Johnson, S. L. Jones, D. Petit, K. Pollock, C. Smith, J. Trapp, and E. Welling. 2000. A programmatic review of the North American Breeding Bird Survey. Report of a peer review panel. U.S. Geological Survey (USGS), Patuxent Wildlife Research Center, Laurel, Maryland, USA. https://www.pwrc.usgs.gov/bbs/bbsreview/bbsfinal.pdf
Pardieck, K. L., D. J. Ziolkowski Jr., M.-A. R. Hudson, and K. Campbell. 2016. North American Breeding Bird Survey dataset 1966 - 2015. Version 2015.1. U.S. Geological Survey (USGS), Patuxent Wildlife Research Center, Laurel, Maryland, USA.
Payne, N. 1976. Red squirrel introduction to Newfoundland. Canadian Field-Naturalist 90:60-64.
Peters, H. S., and T. D. Burleigh. 1951. The birds of Newfoundland. Riverside, Cambridge, Massachusetts, USA.
R Core Team. 2017. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rae, L. F. 2014. Assessing the impacts of moose-induced habitat change on a forest bird community. Thesis. Memorial University of Newfoundland, St. John’s, Newfoundland and Labrador, Canada. https://research.library.mun.ca/8320/1/thesis.pdf
Ralston, J., and W. V. DeLuca. 2020. Conservation lessons from the study of North American boreal birds at their southern periphery. Diversity 12(6):257. https://doi.org/10.3390/d12060257
Smith, A. C., M-A. R. Hudson, V. I. Aponte, and C. M. Francis. 2020. North American Breeding Bird Survey - Canadian trends website. Data-version 2019. Environment and Climate Change Canada, Gatineau, Quebec, Canada. https://wildlife-species.canada.ca/breeding-bird-survey-results
Stralberg, D., S. M. Matsuoka, A. Hamann, E. M. Bayne, P. Sólymos, F. K. A. Schmiegelow, X. Wang, S. G. Cumming, and S. J. Song. 2015. Projecting boreal bird responses to climate change: the signal exceeds the noise. Ecological Applications 25(1):52-69. https://doi.org/10.1890/13-2289.1
Thompson, I. D., H. A. Hogan, and W. A. Montevecchi. 1999. Avian communities of mature balsam fir forests in Newfoundland: age dependence and implications for timber harvesting. Condor 101(2):311-323. https://doi.org/10.2307/1369994
Torrenta, R. 2021. High elevation landbird program mountain birdwatch 2.0. 2020 report. Birds Canada, Atlantic Region, Sackville, New Brunswick, Canada.
Townsend, J. M., K. P. McFarland, C. C. Rimmer, W. G. Ellison, and J. E. Goetz. 2020. Bicknell’s Thrush (Catharus bicknelli). Version 1.0. In P. G. Rodewald, editor. Birds of the World. Cornell Lab of Ornithology, Ithaca, New York, USA.
Vassallo, M. I., and J. C. Rice. 1981. Differential passerine density and diversity between Newfoundland and offshore Gull Island. Wilson Bulletin 93(3):340-349. https://sora.unm.edu/sites/default/files/journals/wilson/v093n03/p0340-p0349.pdf
Veech, J. A., and T. O. Crist. 2007. Habitat and climate heterogeneity maintain beta-diversity of birds among landscapes within ecoregions. Global Ecology and Biogeography 16(5):650-656. https://doi.org/10.1111/j.1466-8238.2007.00315.x
Wallace, G. J. 1939. Bicknell’s thrush, its taxonomy, distribution, and life history. Proceedings of the Boston Society of Natural History 41(6):211-402.
Whitaker, D. 2010. The status of Gray-cheeked Thrush (Catharus minimus) in Newfoundland and Labrador. Report No. 24. Newfoundland and Labrador Species Status Advisory Committee, St. John’s, Newfoundland and Labrador, Canada.
Whitaker, D. M. 2015. The colonization of Newfoundland by red squirrels (Tamiasciurus hudsonicus). Osprey - Newfoundland Natural History Society Quarterly 46:23-29.
Whitaker, D. M., P. D. Taylor, and I. G. Warkentin. 2015. Gray-cheeked Thrush (Catharus minimus) distribution and habitat use in a montane forest landscape of western Newfoundland, Canada. Avian Conservation and Ecology 10(2):4. https://doi.org/10.5751/ACE-00778-100204
Whitaker, D. M., I. G. Warkentin, K. A. Hobson, P. Thomas, and R. Boardman. 2018. Fall and winter movements of Newfoundland Gray-cheeked Thrushes (Catharus minimus minimus). Animal Migration 5(1):42-48. https://www.degruyter.com/document/doi/10.1515/ami-2018-0004/html
Whitaker, D. M., I. G. Warkentin, J. P. B. McDermott, P. E. Lowther, C. C. Rimmer, B. Kessel, S. L. Johnson, and W. G. Ellison. 2020. Gray-cheeked Thrush (Catharus minimus). Version 1.0. In P. G. Rodewald, editor. Birds of the World. Cornell Lab of Ornithology, Ithaca, New York, USA.
Wood, S. 2017. Generalized additive models: an introduction with R. Second edition. CRC, Boca Raton, Florida, USA.
Wood, S. 2021. Package ‘mgcv’. R package. Version 1.8-38. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/mgcv/mgcv.pdf
Zuur, A. F., E. N. Ieno, N. J. Walker, A. A. Saveliev, and G. M. Smith. 2009. Mixed effects models and extensions in ecology with R. Springer, New York, New York, USA.
Fig. 1
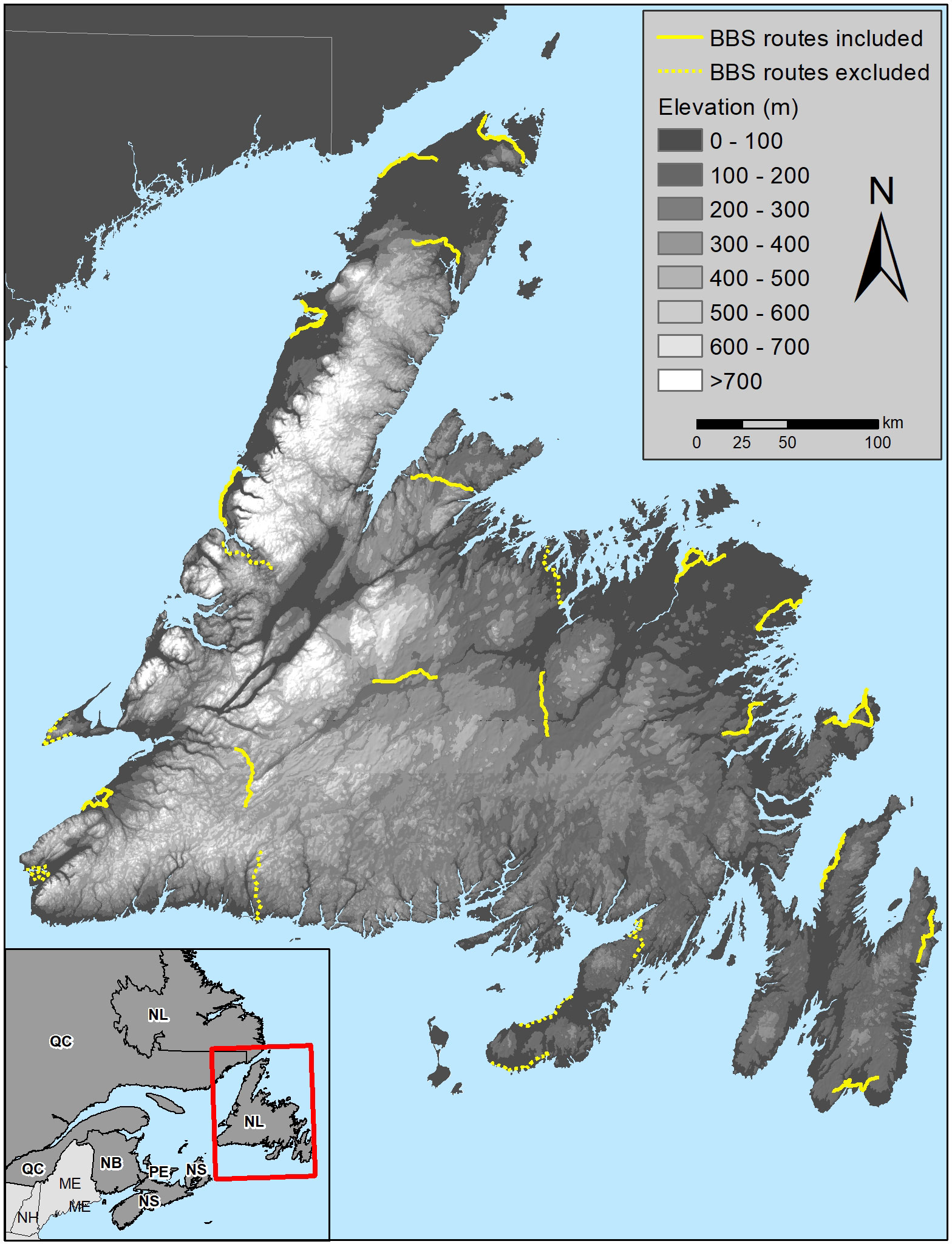
Fig. 1. Topographic map of the island of Newfoundland showing the 17 breeding bird survey routes included in our analyses (solid yellow) as well as additional routes that were not included (dotted yellow), either because they were not surveyed at least 3 times from 1974-1984 and 2001-2015, or because Gray-cheeked Thrushes (Catharus minimus) have never been observed on that route.
Fig. 2
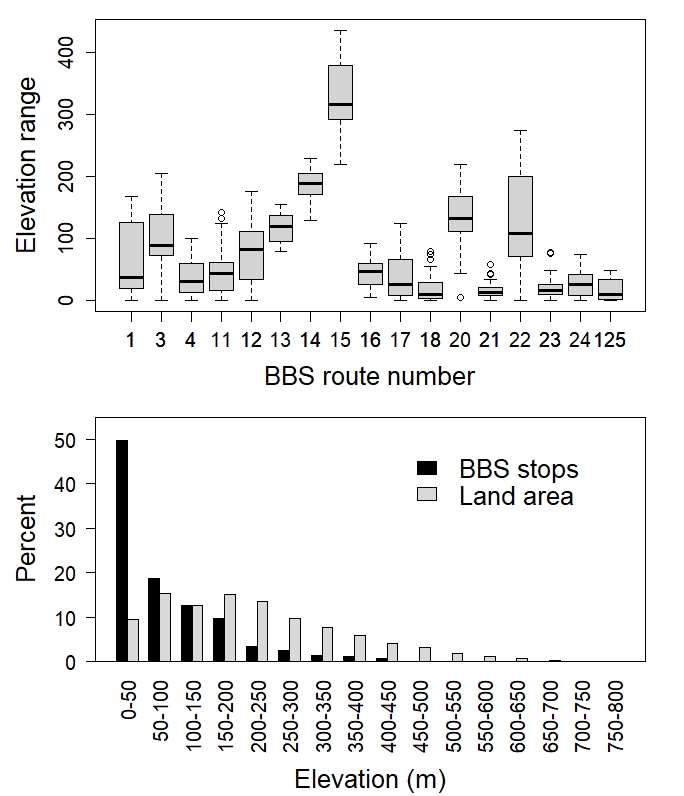
Fig. 2. Elevation range of stops along each of 17 breeding bird survey (BBS) routes on Newfoundland (top) and combined elevation distribution of all 850 stops across these 17 routes, as well as the elevation distribution of land on the island of Newfoundland (bottom). For the top figure, values presented are the median (horizontal line), first and third quartiles (boxes), ± 1.5 times the interquartile range (whiskers) with stops outside that range illustrated by open circles.
Fig. 3
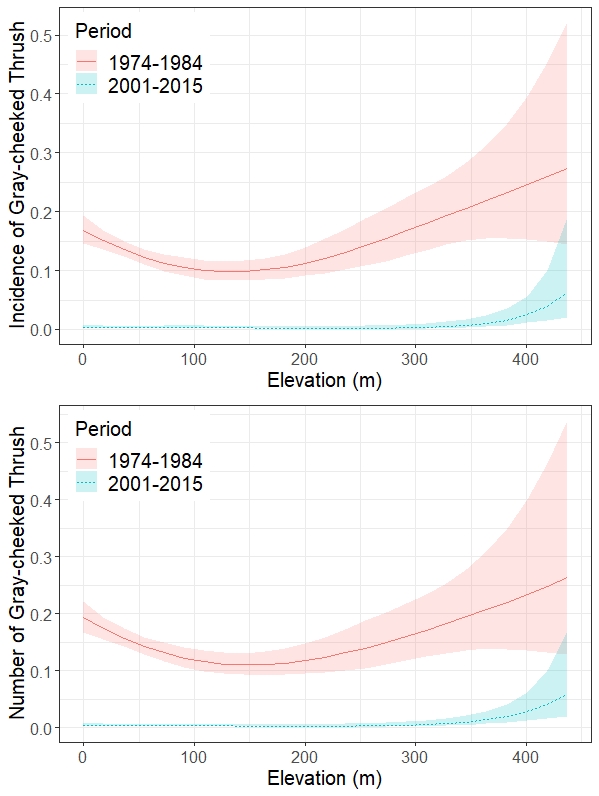
Fig. 3. Incidence (top) and mean number (bottom) of Gray-cheeked Thrushes (Catharus minimus) at breeding bird survey (BBS) stops as a function of elevation across 17 BBS routes on Newfoundland sampled during 2 time periods, 1974-1984 and 2001-2015. The model assessed the interaction between time period and elevation and was fit using a generalized additive model with either a binomial error distribution and a clog-log link function (top; incidence of thrushes) or a negative binomial error distribution and a log link function (bottom; number of thrushes).