







The following is the established format for referencing this article:
Silva, L. F., and E. Guilherme. 2023. Understory bird assemblage in a riparian environment dominated by Cecropia membranacea (Urticaceae) in southwestern Amazonia. Journal of Field Ornithology 94(1):11.ABSTRACT
Studies on bird assemblage structure in pioneer environments allow us to understand the use of space, distribution patterns, ecosystem services, and population dynamics of these ephemeral habitats. However, studies on the bird community related to these types of habitats in the southwest of the Brazilian Amazon are rare. We compared the bird assemblage structure of an area dominated by embaúba-da-várzea (Cecropia membranacea Trécul) with the adjacent alluvial rainforest along the Chandless River, a right-margin tributary of the Purus River, in the southwest Brazilian Amazon. Birds were captured with mist nets and banded (with a metal ring) along trails in both habitats. The following environmental variables were measured: temperature, humidity, and canopy openness. Understory bird assemblage was similar in the two habitats, with no influence from the environmental variables sampled. Average similarity of understory species composition between the two habitats was 77.6%, Pipra fasciicauda being the species that most contributed to this similarity (10.76%). Despite their almost homogeneous canopy and inconspicuous understory composed of short life cycle plants, due to the impact of river dynamics, areas dominated by C. membranacea have an understory bird assemblage composition similar to that of the adjacent alluvial rainforest. Understory species move horizontally and easily between these two habitats. Habitats dominated by C. membranacea on the lower Chandless River, contrary to expected, harbor a rich bird fauna. Given its importance to biodiversity maintenance, the protection of this habitat is recommended, as well as the undertaking of further biological studies on this unique Amazonian ecosystem.
RESUMEN
Los estudios sobre la estructura del ensamble de aves en ambientes pioneros nos permiten comprender el uso del espacio, los patrones de distribución, los servicios ecosistémicos y la dinámica poblacional de estos hábitats efímeros. Sin embargo, son raros los estudios sobre la comunidad de aves relacionada con este tipo de hábitats en el suroeste de la Amazonia brasileña. Comparamos la estructura del ensamble de aves de un área dominada por embaúba-da-várzea (Cecropia membranacea Trécul) con la selva aluvial adyacente a lo largo del río Chandless, un afluente del margen derecho del río Purus, en el suroeste de la Amazonia brasileña. Las aves fueron capturadas con redes de niebla y anilladas (con un anillo metálico) a lo largo de senderos en ambos hábitats. Se midieron las siguientes variables ambientales: temperatura, humedad y apertura del dosel. El ensamble de aves del sotobosque fue similar en los dos hábitats, sin influencia de las variables ambientales muestreadas. La similitud media de la composición de especies del sotobosque entre ambos hábitats fue del 77,6%, siendo Pipra fasciicauda la especie que más contribuyó a esta similitud (10,76%). A pesar de tener un dosel casi homogéneo y un sotobosque poco visible compuesto por plantas de ciclo vital corto, debido al impacto de la dinámica fluvial, las áreas dominadas por C. membranacea tienen una composición de aves del sotobosque similar a la de la selva aluvial adyacente. Las especies del sotobosque se mueven en sentido horizontal y fácilmente entre estos dos hábitats. Los hábitats dominados por C. membranacea en la parte baja del río Chandless, al contrario de lo esperado, albergan una rica avifauna. Dada su importancia para el mantenimiento de la biodiversidad, se recomienda la protección de este hábitat, así como la realización de nuevos estudios biológicos sobre este ecosistema amazónico único.
INTRODUCTION
In southwestern Amazonia, all rivers are meandering, highly sinuous, and dynamic, yielding valuable information on the formation of ecosystems that harbor and maintain biodiversity and population dynamics of ephemeral habitats formed by the influence of annual flood pulses (Junk et al. 2012). In these rivers, during the dry season, a beach is formed on the convex side of the channel and a cliff is formed on the concave side (Novaes 1958). Each bend in the river, on its convex side, is composed of a sandbar where a vegetation mosaic called “pioneer formation of fluvial influence” (PFFI; SEMA 2010) appears. In these rivers’ PFFI areas, a pioneer vegetation grows in such a way that with its roots fixed to the soil, acts as a kind of barrier reducing river velocity, giving way to a more complex forest because the distance from the margin increases (da Cunha et al. 2015). As succession stages progress, a type of pioneer forest, called open alluvial rainforest, with palm trees and/or bamboo appears and it may extend for several kilometers from the river margin (IBGE 2005).
The force of water, a type of natural disturbance, increases river bend curvature, altering local distribution of several biological groups including birds, amphibians, reptiles, and mammals. This is the result of homogenization and reduction in resources availability, especially for the bird fauna (Silva 2000, Parolin 2002, Wittmann et al. 2004, Chettri et al. 2005, Christianini and Martins 2015). The upper Purus River basin, in southwestern Amazonia, is home to many rare and endemic species of plants and animals (Silveira and Daly 2000, Guilherme and Dantas 2011). Because of the river dynamics, the soil along the margins is unstable and constantly reworked (Toivonen et al. 2007, Pereira et al. 2016). For this reason, short life-cycle plants establish themselves in its meanders (Rosenberg 1990, SEMA 2010, Laranjeiras et al. 2019), forming pioneer habitats of river influence. These habitats are classified by Junk et al. (2012) as early secondary tree communities. Such vegetation assemblages establish themselves on soils exposed during the low-water periods, which are highly fertile and viable for pioneer plants. These recently formed habitats harbor vegetation dominated by Urticaceae species of the genus, Cecropia Loefl, with two dominating species: Cecropia latiloba and Cecropia membranacea (Berg 1978).
Several environmental factors alter the composition and distribution of many bird species along Amazonian rivers, such as: canopy and understory temperature (Almeida 2018), light incidence, assessed by canopy openness (Novaes 1958), and pioneer environments dominated by Cecropia trees (Rosenberg 1990). When considering temperature, it is important to note that birds are homeothermic, i.e., organisms that maintain a constant internal body temperature, which leads them to select their habitat according to their adaptation capability because in many cases they cannot withstand extreme temperatures (Abreu and Abreu 2011). Some species of forest birds adapt to the inner forest microhabitat, with specific temperature and humidity, becoming habitat specialists. In these cases, species become adapted to living in the terra firme forest dark understory, which prevents them from coexisting with species that live in habitats with a different microclimate, such as those formed by rivers. Generalist species constitute an exception (Remsen and Parker 1983).
The habitats formed by rivers, especially those dominated by pioneer plants, harbor significant bird diversity, as suggested by several studies (Novaes 1958, Marcondes-Machado and Oliveira 1988, Rosenberg 1990, Terborgh et al. 1990, Terborgh and Petren 1991, Parolin 2002, Laranjeiras et al. 2019, Del-Rio et al. 2021) but remain fully uninvestigated in some regions of southwestern Amazonia, such as the upper Purus River region. The Chandless River is one of the Purus’ right-margin tributaries with little anthropic influence (Pereira et al. 2016). The sediment deposition zones of its meanders harbor a PFFI dominated by C. membranacea whose understory bird assemblage structure has never been studied.
Thus, we aim to compare the structure of bird assemblages in two types of PFFI along an Amazonian River, within Chandless State Park. Because human population density in the park’s area is insignificant, habitats are highly preserved (Pereira et al. 2016). Our main objectives are: (1) to describe understory bird assemblage structure (composition, richness, and abundance) in a PFFI dominated by C. membranacea comparing it to the adjacent alluvial rainforest, and (2) to test whether the understory bird assemblages in these two habitats are being influenced by environmental factors such as temperature, air humidity, and canopy cover.
METHODS
Study area
We carried out our study at Chandless State Park (PEC), 9°21′31.26″ S, 69°55′34.55″ W, headquarters, a conservation unit created by decree 10.670 dated September 2, 2004. Chandless State Park occupies an area of 695,303 ha, corresponding to 4.23% of Acre State territory and lies within Manoel Urbano, Santa Rosa do Purus and Sena Madureira municipalities reaching the border with Peru (SEMA 2010).
The Chandless State Park is crossed by the Chandless River, a right-margin tributary of the Purus River (SEMA 2010). The park’s vegetation is characterized by a mosaic, dominated by open rainforest with bamboo and/or palm trees, in addition to dense rainforest, alluvial rainforest, and high terraces of open floodplain forest (SEMA 2010). The soil on the margins of the Chandless River is hydromorphic gleyed, i.e., made of pure sand and more clayey soil on terra firme (Amaral 2000, SEMA 2010).
Average temperatures in the region range between 24ºC and 25ºC and may reach a minimum of 10ºC depending on the time of the year. Altitude ranges from 180 to 370 m (SEMA 2010). Total annual rainfall varies from 1600 mm to 2750 mm, and the severe dry period is from June to August. Relative humidity varies from 80 to 90% (Silveira and Daly 2000, SEMA 2010).
On the margins of the Chandless River’s bends there are various spots of PFFI (areas where succession is immediately above the sandbar formed by the river bend) in which the habitats dominated by C. membranacea and the adjacent alluvial rainforest are found and where we set up the trails for data collection (Fig. 1).
Bird capture and identification
Captures with mist nets occurred from July 2020, during the low-water period and when C. membranacea is at the final stage of fruiting (Parolin 2002), until December 2020. We suspended our field work during the wet period and the intense river floods, resuming the captures in May 2021. We used five 14 x 3 m mist nets in each habitat. To optimize captures on sampling days, we opened the nets at dawn (around 05h45) and closed them around 16h45.
For sampling, we randomly selected seven bends along the Chandless River (Fig. 1). The selected bends were at least 1 km from each other and at least 100 m in length and 70 m in width, for both habitats studied. In each sampling locality, two 250 m-long parallel trails were set up, 100 m from each other, where we installed 5 mist nets in each habitat (Fig. 2). We sampled 28 trails, 14 in the C. membranacea dominated habitat (EM) environment and 14 in the adjacent alluvial rainforest (AF) environment, with 2 trails for each of the 7 curves. To avoid collecting data in a transitional environment, sampling in the alluvial rainforest started at least 100 m from the contact between the forest and the EM. Sampling effort for each trail was 2 days/net (105.6 hours/net), repeated twice during the sampling period. After all sites were sampled, we performed a new round of sampling.
We identified all captured birds to the species level, with the help of illustrated guides (Schulenberg et al. 2010, Guilherme 2016). After banding each bird with numbered metal bands, we released them on the same site where they were captured. Recaptures were recorded and were not included in abundance analyses. Aluminum bands were provided by Centro Nacional de Pesquisa e Conservação de Aves Silvestres (CEMAVE/IBAMA) as part of project 4521, under the direction of Edson Guilherme (Registry Number 324654). Banding followed CEMAVE/IBAMA protocols (IBAMA 1994). Nomenclature followed the Brazilian Ornithological Records Committee (CBRO; Pacheco et al. 2021).
Environmental data sampling
We used an Incoterm 7666.02.0.00 thermal hygrometer to record humidity and temperature data. During the capture period, we set up the hygrometer between trails and recorded the data at the opening of the nets at noon and at the closing of the nets in both habitats.
Canopy cover data were recorded with a vertical sighting tube, both in the EM and AF environments (Robards et al. 2000). Looking through the instrument, we recorded 1 for the presence of any plant part (leaves, branches, or trunk) and 0 when plant cover was absent and only the sky was visible. On a transect, we scored 70 points for both habitats. We sum all presence points (1) of each transect, divide by 70 and multiply by 100 to obtain the percentage of canopy cover per transect (%). To obtain the canopy cover by a curve in each habitat, we add the two percentages (%) and divide by two. These data were recorded only once, at the beginning of sampling for all sampled curves.
Data analysis
We constructed a rarefaction and extrapolation curve to compare bird community richness data in the C. membranacea and in the adjacent alluvial rainforest habitats. We then used a paired t-test to check whether the captured species abundance means differed between both habitats sampled along the seven river bends. The statistical significance level used was p < 0.05.
The similarity among captured species composition between sampled habitats was calculated through a multilevel comparison of the data with permutational multivariate analysis of variance (PERMANOVA). We constructed a distance matrix between these areas and ran a principal coordinates analysis (PCoA) using the Vegan software package and the Jaccard index.
We ran similarity percentage (SIMPER) using the Bray-Curtis similarity index to check which taxa most contributed to the similarity in species composition between the C. membranacea dominated habitat and the adjacent alluvial rainforest (Clarke 1993).
A redundance analysis test, with the Bray-Curtis index, was used to verify whether environmental factors (temperature, humidity, and canopy openness) influenced bird species composition in the two habitats studied. For this test, the “rda” function of the Vegan package was used, considering 999 permutations. The RDA result was checked by a variance analysis with the “anova” function (Oksanen et al. 2019).
To check if the environmental variables individually influenced species richness and abundance in the two habitats (EM and AF), we ran a multiple linear regression using the “lm” function available in the R program. After the model was generated, we used the “summary” function for a general reading of the data. Finally, we used the “step” function, with the stepwise method, to check which variables really influenced richness and abundance for each habitat studied.
All analyses were made with resources available in the R 4.1.1, Microsoft Excel 2016 and past 4.1 softwares (Hammer et al. 2001, Oksanen et al. 2019). We used each transect within each habitat as a unit of replication to compare community composition of birds, i.e., is it summarized over river bends within each habitat (n = 2 x 7 = 14)?
RESULTS
Bird fauna in Cecropia membranacea dominated understory in relation to adjacent alluvial rainforest
After 5799 hours of capture effort, a total of 471 individuals were banded (228 were captured in EM and 243 in AF). We also had 26 recaptures. The banded birds belonged to 87 species, 29 families, and 11 orders (Appendix 2). In EM, 65 species were caught (74.7% of the species total) representing 23 families and 10 orders, whereas in AF, 61 species were caught (70.1%), representing 24 families and 8 orders.
The rarefaction and extrapolation curve in both habitats did not show signs of stabilization (asymptote). See Fig. 3.
The families with greatest abundance of individuals in the two habitats sampled were Thamnophilidae (EM = 64.9%, AF = 52.6%), followed by Pipridae (EM = 28%, AF = 32.3%), and Dendrocolaptidae (EM = 21.4%, AF = 21%). The three families with greatest species richness were Thamnophilidae (EM = 24.6%, AF = 19.7%), Dendrocolaptidae (EM = 12.3%, AF = 14.7%), and Tyrannidae (EM = 9.2%, AF = 8.2%). See Appendix 1.
In EM, Pipra fasciicauda was the most abundant species (n = 29), followed by Akletos goeldii (n = 18), and Taraba major (n = 17). In AF, P. fasciicauda was also the most abundant species (n = 38), followed by Phlegopsis nigromaculata (n = 16), and Cacicus cela (n = 12). See Appendix 1.
The paired t-test yielded t = -0.520, df = 6, p = 0.621, non-significant. The SIMPER indicated that species composition in the habitats studied has an average similarity of 77.6%. This test showed that P. fasciicauda (10.76%) was the species that most contributed to the similarity between habitats EM and AF (Appendix 2).
Principal coordinate analysis, based on species composition of the birds caught (Fig. 4), showed that the understory bird assemblage structure in EM did not differ significantly from that in AF (R² = 0.04, adjusted p = 0.166).
Effect of environmental factors on abundance and richness of understory birds
At EM, average temperature was around 25.03ºC ± 1.31, average humidity was 78.13% ± 10.69, and average canopy cover was 84.99% ± 6.92. In AF, average temperature was around 23.83ºC ± 1.32, average humidity was 80.88% ± 10.70, and average canopy cover was 88.94% ± 3.81.
Redundancy analysis showed that environmental factors are not good predictors of species distribution for habitats EM and AF (R² = 0.122). In EM areas, environmental data only explained 14.54% of the variation in bird composition, and in AF areas, environmental data only explained 18.02% of the variation. The ANOVA using the global test on the RDA result proved non-significant in EM (p = 0.61, R² = 0.22) and in AF (p = 0.18, R² = 0.25), i.e., no difference in species distribution between habitats can be explained by the variables analyzed (Fig. 5).
Multiple linear regression indicated no relation between any of the 3 environmental variables and species richness (p = 0.09, R² = 0.45, F = 2.73, df = 10) or abundance (p = 0.12, R² = 0.25, F = 2.44, df = 10) for the habitats studied.
DISCUSSION
Understory species richness in the PFFIs under influence of the Chandless River
Many bird species are associated with habitats created by rivers, and although they are not entirely defined as specialists, the presence of several of them is associated with specific forest succession stages (Rosenberg 1990, Cohn-Haft et al. 2007, Smith et al. 2014, Laranjeiras et al. 2019, Naka et al. 2020). Although these habitats do not differ much from the adjacent alluvial rainforest in species richness, the taxa specialized in exploiting environments linked to water dynamics contribute to alpha and beta diversity in the Amazon biome (Rosenberg 1990, Laranjeiras et al. 2019).
Species richness recorded in the habitats dominated by C. membranacea and in the adjacent alluvial rainforests (87 species) was 35.4% greater than the 57 species reported by Novaes (1958) in the varzea forest of the higher Juruá River, Acre State. However, it was 62.34% and 75.29% lower, respectively, than the species richness reported by Rosenberg (1990) from floodplain areas and fluvial islands in northern Peru and southern Colombia, and by Laranjeiras et al. (2019) from the Negro River basin. Recently, Del-Rio et al. (2021) recorded 128 species in varzea forests and along river margins on a tract of the middle Juruá River, Amazonas State, a result 32% greater than the one found in the Chandless River tract herein studied.
The presence of the austral migrants Elaenia spectabilis and Lathrotriccus euleri and of the Nearctic migrants Catharus swainsoni and Empidonax alnorum was recorded in the study areas. These birds arrive from northern South America to use Amazon habitats as wintering grounds for food, rest, and shelter during migration (Beja et al. 2010, Guilherme 2016). According to Rosenberg (1990), habitats formed by rivers in western Amazonia are used as migratory pathways for Nearctic species from August to April, which coincides with the period when both C. swainsoni and E. alnorum were mist netted in this study.
Two species associated with riparian habitats in western Amazonia and very rarely sampled, Eubucco tucinkae and Crypturellus bartletti, were captured during the present study. In addition, 13 species endemic to Acre State were found in the habitats studied, representing 37% of the 35 bird species endemic to the state (Guilherme 2016), reinforcing the idea that these areas are important for maintaining species of interest for conservation and require attention in terms of management and preservation policies (DeLuca 2012).
Understory bird community structure and environmental variables
The structure of the bird community in the low understory (0-3 m) does not differ significantly between the two habitats studied along the Chandless River, and it seems not to be affected by environmental variables (temperature, humidity, canopy cover). Bird species show no preference for either, as foraging areas. According to Jahn (2011), heterogeneous vegetation formations significantly affected bird communities of terra firme habitats, a result quite different from what we found in the PFFIs, i.e., in this pioneers environment bird community, dynamics seem to follow a different pattern from that observed in mature terra firme forests.
Because both habitats studied are adjacent, a constant seed rain occurs stemming from AF and transported by a number of dispersal agents to EM, favoring the presence of pioneer plants like the Vismia and Miconia species. Also, the incoming waters bring more seeds to EM during the annual floods. In the understory of the C. membranacea dominated areas, this pioneer formation certainly attracts some bird species that live in the adjacent AF understory, such as Phaethornis hispidus, Dendrocincla fuliginosa, and Xiphorhynchus obsoletus, commonly associated to varzeas (Beja et al. 2010) and which we recorded in both habitats, without showing any specific preference.
Four species, Pipra fasciicauda, Phlegopsis nigromaculata, Taraba major, and Akletos goeldii, accounted for 25.18% of the similarity between environments (Appendix 3). Although it cannot be said that these species are intimately associated with the low EM understory because there was no significative difference in captures between the two habitats studied, it is likely that they use this habitat regularly and find adequate conditions for foraging and for other biological activities therein. In other words, these species possess some type of specialization for living in the understory of habitats dominated by Cecropia species in Amazonia (Novaes 1958, Remsen and Parker 1983, Rosenberg 1990, Borges and Carvalhaes 2000, Smith et al. 2014, Junk et al. 2020).
CONCLUSION
The small tracts of pioneer formation of fluvial influence dominated by C. membranacea found in narrow-margin rivers of southwestern Amazonia, such as the Chandless River, harbor an understory bird assemblage structure not distinct from that found in the adjacent alluvial rainforest. Only now are these Amazon habitats becoming the subject of ecological studies, and it seems advisable to expand the scope to include wider rivers, such as Purus, Juruá, and Madeira, which have much more extensive pioneer formations along their meanders than the Chandless River. The aim is to check whether or not the patterns found herein are the same, or whether in more extensive areas covered by C. membranacea, the bird assemblage structure differs from the one found in the adjacent alluvial rainforest because of the specialization of some species for exploiting this environment.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
This study was carried out with support from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES/Brazil), financing code 001, and the Secretaria do Meio Ambiente e das Políticas Indígenas (SEMAPI/AC). We thank CEMAVE/ICMBio for providing metallic bands. We thank the members of the UFAC Ornithology Laboratory - Ornitolab - for their help and companionship and the ornithologist, Victor Castanho, for his help in the field. We especially thank the Chandless State Park manager, Ricardo Antônio de Andrade Plácido, and his entire team for their logistical support during the field data collection.
DATA AVAILABILITY
The data/code that support the findings of this study are openly available in Zenodo at https://doi.org/10.5281/zenodo.7141620. Ethical approval for this research study was granted by SISBIO and Centro Nacional de Pesquisa e Conservação de Aves Silvestres (CEMAVE/IBAMA) as part of project 4521, under the direction of Professor Edson Guilherme (registry number 324654).
LITERATURE CITED
Abreu, V. M. N., and P. G. de Abreu. 2011. Os desafios da ambiência sobre os sistemas de aves no Brasil. Revista Brasileira de Zootecnia 4:1-14. https://www.alice.cnptia.embrapa.br/bitstream/doc/901939/1/osdesafiosdaambienciasobreossistemas.pdf
Almeida, S. M. 2018. Efeitos ecológicos e evolutivos nos padrões de diversidade de aves na Amazônia. Thesis. Universidade Federal do Pará, Belém, Pará, Brazil.
Amaral, E. F. 2000. Caracterização pedológica das unidades regionais do Estado do Acre. Embrapa Acre, Circular Técnica, Rio Branco, Acre, Brazil. https://www.embrapa.br/busca-de-publicacoes/-/publicacao/498482/caracterizacao-pedologica-das-unidades-regionais-do-estado-do-acre
Beja, P., C. D. Santos, J. Santana, M. J. Pereira, J. T. Marques, H. L. Queiroz, and J. M. Palmeirim. 2010. Seasonal patterns of spatial variation in understory bird assemblages across a mosaic of flooded and unflooded Amazonian forests. Biodiversity and Conservation 19(1):129-152. https://doi.org/10.1007/s10531-009-9711-6
Berg, C. C. 1978. Espécies de Cecropia da Amazônia Brasileira. Acta Amazonica 8(2):149-182. https://doi.org/10.1590/1809-43921978082149
Borges, S. H., and A. Carvalhaes. 2000. Bird species of black water inundation forests in the Jaú National Park (Amazonas State, Brazil): their contribution to regional species richness. Biodiversity and Conservation 9(2):201-214. https://doi.org/10.1023/A:1008902306499
Chettri, N., D. C. Deb, E. Sharma, and R. Jackson. 2005. The relationship between bird communities and habitat. Mountain Research and Development 25(3):235-243. https://doi.org/10.1659/0276-4741(2005)025[0235:TRBBCA]2.0.CO;2
Christianini, A. V., and M. M. Martins. 2015. Ecologia reprodutiva e produção de sementes - Frugivoria e dispersão de sementes. Pages 83-101 in F. Piña-Rodrigues, M. Figliolia, A. da Silva, editors. Sementes florestais tropicais: da ecologia à produção. Associação Brasileira de Tecnologia de Sementes (ABRATES), Londrina, Paraná, Brazil.
Clarke, K. R. 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18(1):117-143. https://doi.org/10.1111/j.1442-9993.1993.tb00438.x
Cohn-Haft, M., L. N. Naka, and A. M. Fernandes. 2007. Padrões de distribuição da avifauna da várzea dos rios solimões e amazonas. Pages 287-323 in A. L. Albernaz, editor. Conservação da várzea: identificação e caracterização de regiões biogeográficas. Provárzea, Manaus, Amazonas, Brazil.
da Cunha, C. N., M. T. F. Piedade, and W. J. Junk. 2015. Classificação e delineamento das Áreas Úmidas Brasileiras e de seus acrohabitats. Edufmt, Cuiabá, Mato Grosso, Brazil. http://cppantanal.org.br/wp-content/uploads/2017/04/E-book-Classificacao-e-Delineamento-das-AUs.pdf
Del-Rio, G., M. J. Mutchler, B. Costa, A. E. Hiller, G. Lima, B. Matinata, J. F. Salter, L. F. Silveira, M. A. Rego, and D. C. Schmitt. 2021. Birds of the Juruá River: extensive várzea forest as a barrier to terra firme birds. Journal of Ornithology 162(2):565-577. https://doi.org/10.1007/s10336-020-01850-0
DeLuca, J. J. 2012. Birds of conservation concern in eastern Acre, Brazil: distributional records, occupancy estimates, human-caused mortality, and opportunities for ecotourism. Tropical Conservation Science 5(3):301-319. https://doi.org/10.1177/194008291200500306
Guilherme, E. 2016. Aves do Acre. Edufac, Rio Branco, Acre, Brazil. file:///C:/Users/lesli/Downloads/Aves%20do%20Acre.pdf
Guilherme, E., and S. M. Dantas. 2011. Avifauna of the upper Purus River, State of Acre, Brazil. Revista Brasileira de Ornitologia 19(2):185-199. http://www.revbrasilornitol.com.br/BJO/article/view/4306/pdf_684
Hammer, Ø., D. A. T. Harper, and P. D. Ryan. 2001. Past: paleontological statistics software package for education and data analysis. Paleontologia Eletronica 4(1):1-9. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Instituto Brasileiro de Geografia e Estatística (IBGE). 2005. Potencial florestal do Estado do Acre: relatório técnico, resultados preliminares. Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro, Brazil.
Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA). 1994. Manual de anilhamento de aves silvestres. Editora Ibama, Brasília, Brazil. https://www.gov.br/icmbio/pt-br/centrais-de-conteudo/publicacoes/publicacoes-diversas/fauna-e-flora/manual-de-anilhamento-de-aves-silvestres/manual_de_anilhamento_de_aves_silvestres.pdf
Jahn, O. 2011. Bird communities of the Ecuadorian Choco: a case study in conservation. Bonner Zoologische Monographien, Bonn, Germany.
Junk, W. J., M. T. F. Piedade, J. Schöngart, and F. Wittmann. 2012. A classification of major natural habitats of Amazonian white-water river floodplains (várzeas). Wetlands Ecology and Management 20:461-475. https://doi.org/10.1007/s11273-012-9268-0
Junk, W. J., M. T. F. Piedade, F. Wittmann, and J. Schöngart. 2020. Várzeas Amazônicas: desafios para um manejo sustentável. Editora INPA, Manaus, Amazonas, Brazil.
Laranjeiras, T. O., L. N. Naka, and M. Cohn-Haft. 2019. Using river color to predict Amazonian floodplain forest avifaunas in the world’s largest blackwater river basin. Biotropica 51(3):330-341. https://doi.org/10.1111/btp.12650
Marcondes-Machado, L. O., and M. M. A. De Oliveira. 1988. Comportamento alimentar de aves em Cecropia (Moraceae), em mata Atlântica no Estado de São Paulo. Revista Brasileira de Zoologia 4(4):331-339. https://doi.org/10.1590/S0101-81751987000400005
Naka, L. N., T. O. Laranjeiras, G. R. Lima, A. Plaskievicz, F. Pinto, and T. Gonçalves-Souza. 2020. Climate as a major driver of avian diversity in riparian Amazonian habitats along an environmental gradient. Journal of Biogeography 47:2328-2340. https://doi.org/10.1111/jbi.13957
Novaes, F. C. 1958. As aves e as comunidades bióticas no Alto Rio Juruá, Território do Acre. Boletim do Museu Paraense Emilio Goeldi, Nova Série. Zoologia 14:1-13. https://repositorio.museu-goeldi.br/bitstream/mgoeldi/724/1/B%20MPEG%20Zoo%20n%2014%201958%20NOVAES.pdf
Oksanen, J., F. G. Blanchet, M. Friendly, R. Kindt, P. Legendre, D. McGlinn, P. R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens, E. Szoecs, and H. Wagner. 2019. Vegan: community ecology package. R package. Version 2.5.6. Vienna, Austria. https://cran.r-project.org/web/packages/vegan/index.html
Pacheco, J. F., L. F. Silveira, A. Aleixo, C. E. Agne, G. A. Bencke, G. A. Bravo, G. R. R. Brito, M. Cohn-Haft, G. N. Maurício, L. N. Naka, F. Olmos, S. R. Posso, A. C. Lees, L. F. A. Figueiredo, E. Carrano, R. C. Guedes, E. Cesari, I. Franz, F. Schunck, and V. de Q. Piacentini. 2021. Annotated checklist of the birds of Brazil by the Brazilian Ornithological Records Committee. Second edition. Ornithology Research 29:94-105. https://doi.org/10.1007/s43388-021-00058-x
Parolin, P. 2002. Life history and environment of Cecropia latiloba in Amazonian floodplains. Revista de Biologia Tropical 50(2):531-545. https://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S0034-77442002000200013
Pereira, D. M., C. F. Szlafsztein, and F. A. Araújo. 2016. Avaliação de risco de desastres na bacia hidrográfica do Rio Purus (Brasil) com base em índices compostos. Revista da Anpege 12(17):167-190. https://doi.org/10.5418/RA2016.1217.0008
Remsen, J. V., and T. A. Parker. 1983. Contribution of river-created habitats to bird species richness in Amazonia. Biotropica 15(3):223-231. https://doi.org/10.2307/2387833
Robards, T. A., M. W. Berbach, P. H. Cafferata, and B. E. Valentine. 2000. A comparison of techniques for measuring canopy in watercourse and lake protection zones. California Forestry Note 115:1-15.
Rosenberg, G. H. 1990. Habitat specialization and foraging behavior by birds of Amazonian River islands in northeastern Peru. Condor 92(2):427-443. https://doi.org/10.2307/1368240
Schulenberg, T. S., D. F. Stotz, D. F. Lane, J. P. O’neill, and T. A. Parker. 2010. Birds of Peru. Revised and updated edition. Princeton University Press, Princeton, New Jersey, USA.
Secretaria do Meio Ambiente (SEMA). 2010. Plano de manejo do parque Estadual Chandless I, II, III. Diário oficial da União, Rio Branco, Acre, Brazil. http://semapi.acre.gov.br/wp-content/uploads/sites/20/2020/11/Resumo_Executivo_PEC_jan_2010.pdf
Silva, H. G. V. 2000. Mosaicos de vegetação em Áreas de atuação dos Rios Acre e Purus, Amazônia Sul-Ocidental, Brasil. Dissertation. Universidade Federal do Acre, Rio Branco, Acre, Brazil.
Silveira, M., and D. Daly. 2000. Zoneamento ecológico-econômico do Acre: relatório analítico. Secretaria de Ciência, Tecnologia e Meio Ambiente, Rio Branco, Acre, Brazil.
Smith, B. T., J. E. Mccormack, A. M. Cuervo, M. J. Hickerson, A. Aleixo, C. D. Cadena, J. Pérez-Emán, C. W. Burney, X. Xie, M. G. Harvey, B. C. Faircloth, T. C. Glenn, E. P. Derryberry, J. Prejean, S. Fields, and R. T. Brumfield. 2014. The drivers of tropical speciation. Nature 515(7527):406-409. https://doi.org/10.1038/nature13687
Terborgh, J., and K. Petren. 1991. Development of habitat structure through succession in an Amazonian floodplain forest. Pages 28-46 in S. S. Bell, E. D. McCoy, and H. R. Mushinsky, editors. Habitat structure: the physical arrangement of objects in space. Springer, Dordrecht, The Netherlands.
Terborgh, J., S. K. Robinson, T. A. Parker, C. A. Munn, and N. Piperpont. 1990. Structure and organization of an Amazonian forest bird community. Ecological Monographs 60(2):213-238. https://doi.org/10.2307/1943045
Toivonen, T., S. Mäki, and R. Kalliola. 2007. The riverscape of Western Amazonia - a quantitative approach to the fluvial biogeography of the region. Journal of Biogeography 34(8):1374-1387. https://doi.org/10.1111/j.1365-2699.2007.01741.x
Wittmann, F., W. J. Junk, and M. T. F. Piedade. 2004. The várzea forests in Amazonia: flooding and the highly dynamic geomorphology interact with natural forest succession. Forest Ecology and Management 196(2-3):199-212. https://doi.org/10.1016/j.foreco.2004.02.060
Fig. 1
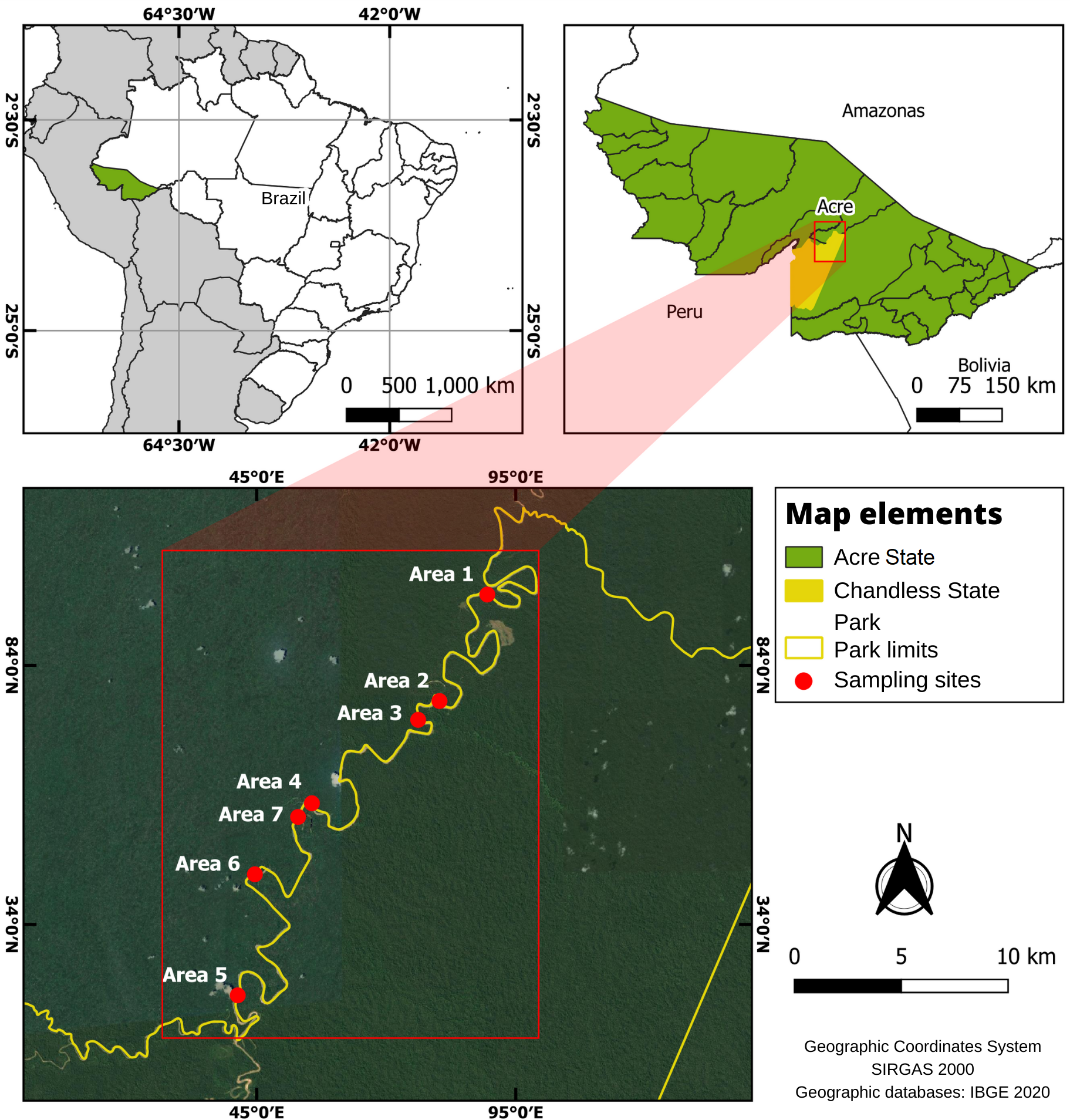
Fig. 1. Localization of Chandless State Park (above) and details of the sites where trails for data collection were set up along the Chandless River (below). Illustration: W. Lima and M. Oliveira.
Fig. 2
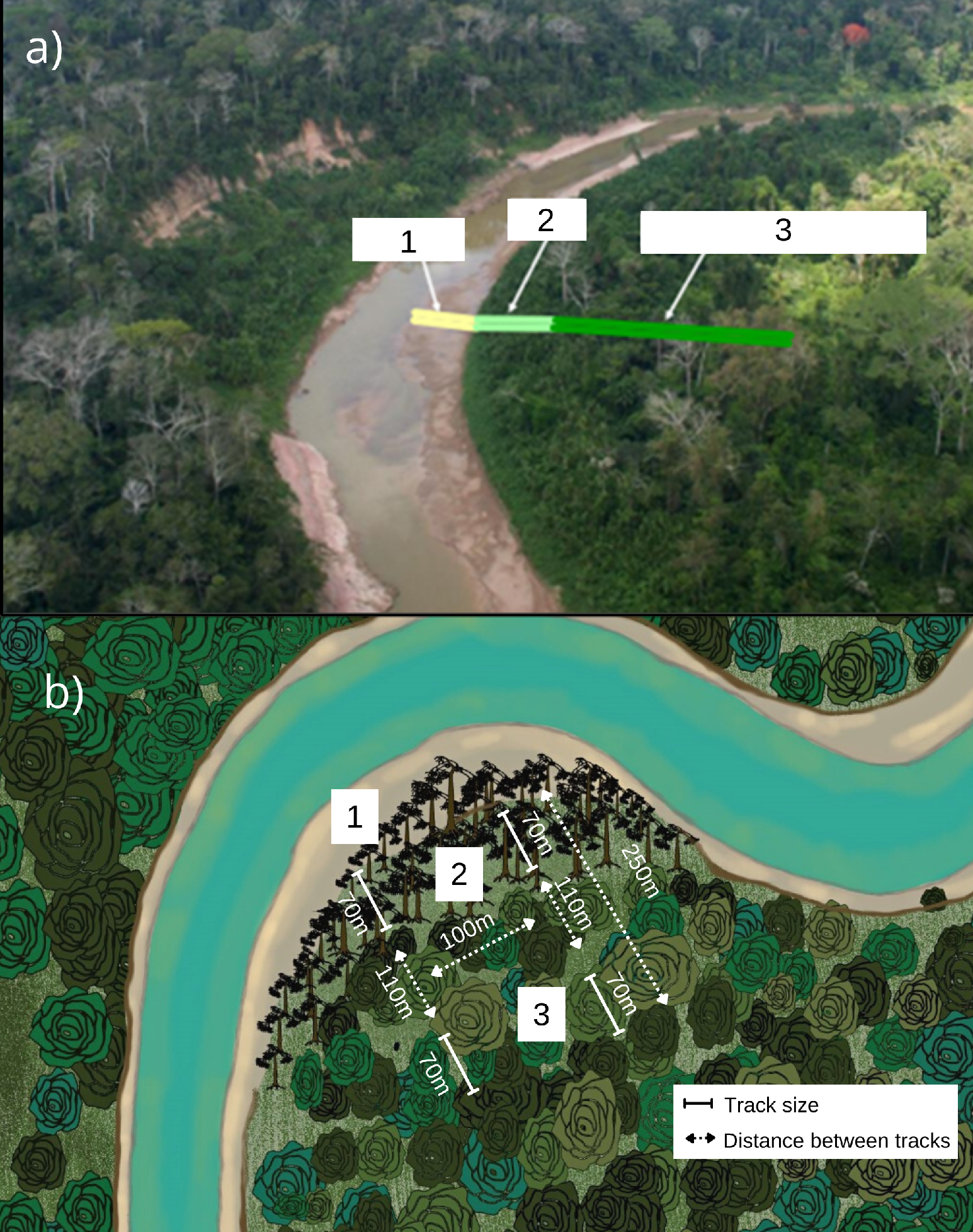
Fig. 2. Schematic representation of (a) study area that comprises (1) sandbar, (2) area dominated by Cecropia, and (3) alluvial open ombrophilous forest, and (b) arrangement of mist nets on trails installed in the Chandless State Park, Manoel Urbano-AC. Adapted from SEMA 2010 and illustration by G. Rodrigues.
Fig. 3
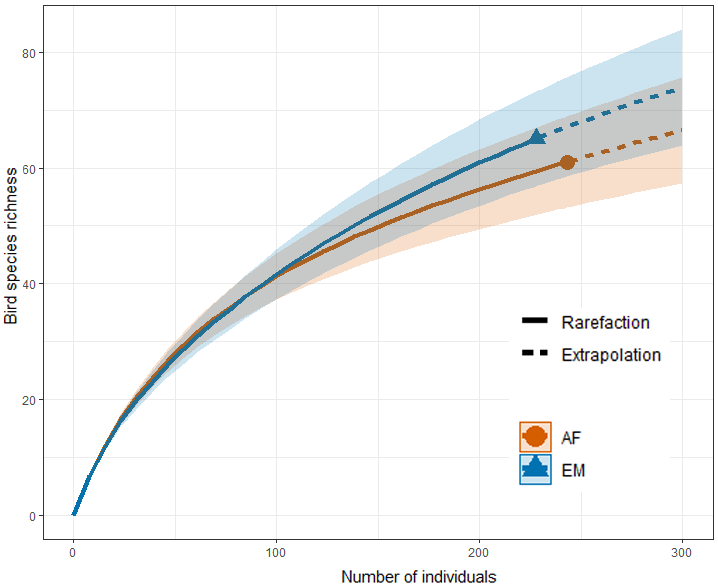
Fig. 3. Rarefaction curves and data extrapolation for birds captured in the Cecropia membranacea dominated habitat (EM) and in the adjacent alluvial rainforest (AF) on 14 trails set on the margins of the Chandless River, Manoel Urbano municipality, Acre State, Brazil.
Fig. 4
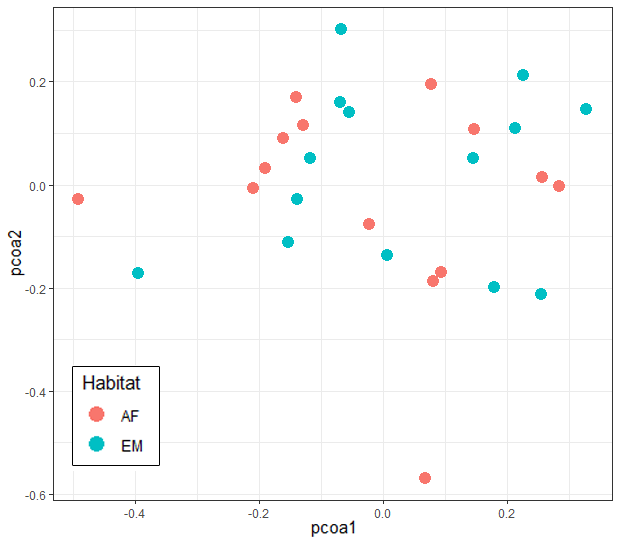
Fig. 4. Principal coordinate analysis (PCoA) using the Jaccard index, done with bird species composition in Cecropia membranacea dominated habitat (EM) and in the adjacent alluvial rainforest (AF) along the Chandless River, Manoel Urbano municipality, Acre State, Brazil.
Fig. 5
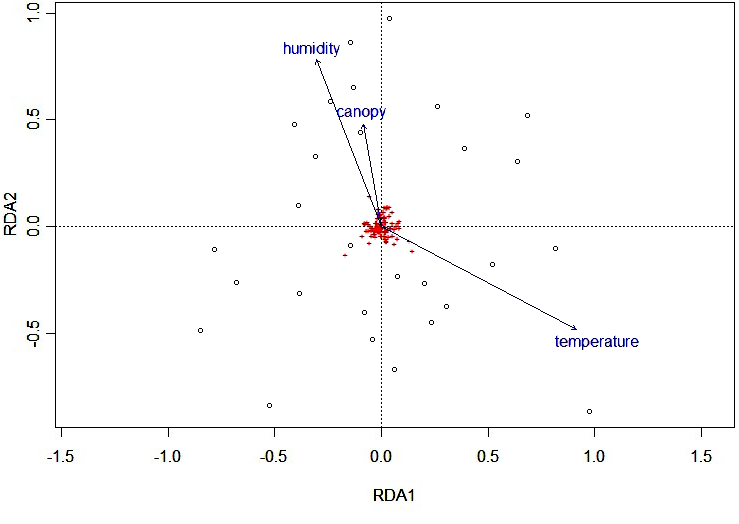
Fig. 5. Redundancy analysis (RDA) in Cecropia membranacea dominated habitat and adjacent alluvial rainforest at the Chandless River, Manoel Urbano municipality, Acre State, Brazil. Note: capture sites (○), captured bird species (+), and environmental factors humidity, canopy, and temperature (→).