







The following is the established format for referencing this article:
Kenow, K. P., L. J. Fara, S. C. Houdek, B. R. Gray, D. J. Heard, M. W. Meyer, T. J. Fox, R. J. Kratt, and C. L. Henderson. 2023. Dive characteristics of Common Loons wintering in the Gulf of Mexico and off the southern U.S. Atlantic coast. Journal of Field Ornithology 94(1):1.ABSTRACT
Common Loons (Gavia immer) winter primarily in marine coastal areas and utilize a forage base that is poorly defined, especially for offshore areas. Information on dive activity is needed for describing foraging strategies and for inferring prey distribution. Archival geolocator tags were used to determine the wintering locations and dive characteristics of adult Common Loons captured and marked on breeding lakes in Minnesota, Wisconsin, and the Upper Peninsula of Michigan. Among loons that completed fall migration, most wintered in the Gulf of Mexico, with smaller proportions wintering off the southern Atlantic Coast or impoundments in the southeastern United States. Adult Common Loons tended to occupy offshore areas of the Gulf of Mexico and the Atlantic Ocean and, on average, spent about 60% of daylight hours foraging. Dive depths were as deep as 50 m (Gulf of Mexico) and dive characteristics indicated that loons were primarily foraging on benthic prey. Total dive duration, time at maximum depth, and post-dive surface intervals increased with dive depths among wintering Common Loons. Our results are expected to contribute to the understanding of the wintering ecology of Common Loons and be useful in informing regional and national conservation planning efforts.
RESUMEN
Los colimbos comunes (Gavia immer) invernan principalmente en zonas costeras marinas y utilizan una base para su alimentación que está mal definida, especialmente en las zonas de altamar. La información sobre la actividad de buceo es necesaria para describir las estrategias de alimentación y para inferir la distribución de las presas. Se utilizaron marcadores geolocalizadores de archivo para determinar los lugares de invernada y las características de inmersión de los colimbos comunes adultos capturados y marcados en lagos de cría de Minnesota, Wisconsin y la Península Superior de Michigan. Entre los colimbos que completaron la migración otoñal, la mayoría invernó en el Golfo de México, y en menor proporción en la costa atlántica meridional o en embalses del sureste de Estados Unidos. Los adultos de colimbo común tienden a ocupar zonas costeras del Golfo de México y el Océano Atlántico y, en promedio, pasan alrededor del 60% de las horas de luz buscando alimento. Las profundidades de inmersión alcanzaron los 50 m (Golfo de México) y las características de la inmersión indicaron que los colimbos se alimentaban principalmente de presas bentónicas. La duración total de la inmersión, el tiempo a máxima profundidad y los intervalos en superficie tras la inmersión aumentaron con la profundidad de la inmersión entre los colimbos invernantes. Se espera que nuestros resultados contribuyan a la comprensión de la ecología de invernada de los colimbos comunes y sean útiles para informar los esfuerzos regionales y nacionales de planificación de la conservación.
INTRODUCTION
Wintering Common Loons (Gavia immer) are found along the Atlantic and Gulf of Mexico (GOM) coasts from Newfoundland and Labrador to Mexico (Paruk et al. 2021), with notable concentrations off the coasts of the Carolinas and Virginia (Haney 1990, Spitzer 1995) and along the Florida Gulf Coast (Jodice 1993, Kenow et al. 2021a). These wintering areas likely support breeding populations from across eastern Canada and the eastern United States (Paruk et al. 2021). For example, Kenow et al. (2002, 2021a) found that adult loons breeding in U.S. states in the Upper Midwest typically wintered in the GOM (79.6%), the southern Atlantic Coast (16.5%), or impoundments in the southern United States (3.9%).
Offshore habitats are likely to be important for wintering Common Loons (Kenow et al. 2021a) with most locations of radio-marked adult loons (95%) wintering in the GOM greater than 14 km offshore with maximum observed distances of >115 km for a few individuals. Along Florida GOM coastal waters, Jodice (1993) documented loons 50–65 km offshore during aerial surveys. In addition, geolocator tags affixed to adult Common Loons (Kenow et al. 2021a) collected pressure data enabling documentation of timing and depth of individual dives on wintering areas. While previous studies documented foraging activities of nearshore wintering loons (McIntyre 1978, Daub 1989, Ford and Gieg 1995), observations have not been reported for loons wintering in offshore habitats. Further, dive depths are an important component of foraging activity that have not been previously examined in wintering Common Loons. Coupled with dive shapes extracted from the data (Schreer et al. 2001), such information could provide insights into dive function and feeding strategies, as well as the types and distribution of prey available to foraging loons. For example, dive data of Common Loons using Lake Michigan during fall migration indicated that loons were primarily foraging on benthic prey (Kenow et al. 2018), which had implications for transfer of botulinum neurotoxin.
Here we describe dive characteristics of wintering loons to provide insight into foraging patterns of loons using offshore habitats in the GOM and southern U.S. coastal waters of the Atlantic Ocean, as well as loons using reservoirs in the southeastern United States. This information is expected to contribute to the understanding of the wintering ecology of Common Loons and be useful in informing regional and national conservation planning efforts. Because we also radio-marked a sample of loons with satellite transmitters, we evaluated potential negative effects of radio-marking on dive characteristics of wintering loons.
METHODS
Movements and wintering distribution of Common Loons breeding in the Upper Midwest
Breeding adult Common Loons were captured on lakes in central and northern Minnesota, central and northern Wisconsin, and the Upper Peninsula of Michigan during summers 2009–2016. Lakes with loon territories considered for inclusion in the study were selected in consultation with Minnesota Department of Natural Resources (DNR), Wisconsin DNR, and U.S. Forest Service - Hiawatha National Forest personnel. Loon territories were monitored for nesting activity and breeding success. Adult loons that successfully produced chicks, and the resulting juveniles, were targeted for capture as they are susceptible to capture using night-lighting techniques (Evers 1993). Each captured loon was marked with a metal, numbered federal leg band and a unique combination of colored plastic leg bands to aid with future field identification of individuals. Both adults of a territorial pair were fitted with archival geolocator tags (“geotagged”; Model LAT 2500; 34.6 x 8.3 mm, 4.4 g; Lotek Wireless Inc.) to collect approximate location and water depth sensor measurements; in some cases adult males were also radio-marked with satellite transmitters (“radio-marked”; Model PTT-100, Microwave Telemetry, Inc.) to collect more precise location estimates.
A combination of adhesive and plastic cable ties was used to affix the geolocator tag to a modified lock-on metal leg band (Kenow et al. 2018). Tags were programmed to record (1) light level (from which daily locations for up to two years would be estimated), (2) temperature (0.02 °C accuracy, ≤0.05 °C resolution) at 30-min intervals, and (3) pressure data (±1% dbar accuracy, 0.05% resolution) at 20-sec intervals during daylight hours providing about 138 twelve-hour days of pressure records; these data would be used to document foraging patterns (dive profiles) of study birds during fall migration (previously reported in Kenow et al. 2021a) and during the first few weeks after arriving at the wintering areas in November and December. Data stored on geolocator tags cannot be transmitted and so require that the geotagged loon be recaptured to recover the tag and download the data. The geolocator tags could store data for several years before those data needed to be downloaded.
Geotagged adult loons were recaptured during subsequent breeding seasons using diurnal lift-net trap and night-light nest-capture techniques (Kenow et al. 2009) and night-lighting during chick rearing (Evers 1993). Geolocator tag data collected over the previous year(s) were downloaded from tags using LAT Viewer Studio software (Lotek Wireless Inc.). Geolocator tag location estimates were determined by light-based geolocations using the template-fitting approach (Ekstrom 2004) in combination with tag temperature (water surface temperature) and pressure (dive depth) data. Template-fit error estimates were used to filter aberrant geolocation estimates. Sea surface temperature (derived from NASA Moderate Resolution Imaging Spectroradiometer [MODIS] imagery) across North America inland lakes, Atlantic coastal waters, and GOM, coupled with diving depth information, were used to improve or obtain location estimates and timing of migration movements when light-based geolocation estimates were unreliable. The Composite Raster and Divergence Tool (Fox 2018) was used to facilitate this process. Geotag temperature data also provided indication of sustained flights of ≥ 1.5 hrs (characterized by prolonged drop in temperature). Flight times were used to set bounds on the distances of migration events (Kenow et al. 2021a). Location estimates were used to determine gross movement patterns and generalized location (e.g., Lake Michigan, reservoirs of southeastern states, Atlantic Coast, GOM) of loons. Archival geolocator and telemetry location data are available at https://doi.org/10.5066/P9UIDEMO (Kenow et al. 2021b).
Satellite transmitters (64 g, ~1.4% of body mass of adult male loons) were implanted in a subsample of adult male Common Loons (marked during July 2010 and July 2011) that were captured on breeding lakes in Minnesota, Wisconsin, and the Upper Peninsula of Michigan. For each loon, a transmitter with percutaneous antenna was surgically implanted in the abdominal cavity following procedures developed by Korschgen et al. (1996). Surgical techniques and the handling and care of loons (Kenow et al. 2021a) were done under approval of the Animal Care and Use Committee of the U.S. Geological Survey Upper Midwest Environmental Sciences Center and complied with the Animal Welfare Act (Public Law 99-198 and 9 CFR Parts 1, 2, and 3).
The satellite transmitters were programmed to transmit on a variable schedule based on the anticipated stage of migration during the loons’ annual cycle: 72 h off during the breeding season; 8 h on - 24 h off while migrating; 6–8 h on - 96 h off on the wintering areas; 8 h on - 24 h off during spring migration; and 8 h on - 96 h off for the remaining life of the transmitter. Loon locations were estimated from the Doppler shift in the transmitter carrier frequency and provided by the Argos system (CLS America, Lanham, Maryland). Location estimates were acquired using Argos Standard Service Processing (Argos Location Classes [LC] 3, 2, 1 and 0) and Auxiliary Location Processing (LC A, B and Z). One standard deviation of nominal accuracy for location estimates with LC 3, 2, 1 and 0 are < 250, 250–500, 500–1500 and > 1500 m, respectively (Argos 2016). We utilized the most accurate locations per 8-h transmission period (based on superior Argos Location Class, number of transmissions, and best signal level) to describe the daily location of each loon.
Interpretation of dives from geotag pressure data
Attributes of adult loon dives (i.e., proportion of time underwater, depth of dive, ascent and descent rates, duration of dive, time duration within 2 m of maximum depth, and post-dive surface interval) were extracted from geolocator tag pressure data, using software code developed by Kenow (2018). When geotagged loons were in freshwater environments, depth was estimated by pressure (dbar) * 1.019716 m/dbar (Sea-Bird 2002). When geotagged loons were in marine environments, depth was estimated from pressure measurements, estimates of gravity at the approximate mean latitude of wintering radio-marked loons in this study (29° N), and depth, all at standard ocean mass salinity of 35 g/kg (Fofonoff and Millard 1983). Dives were considered as submergence below the water surface, which we defined as recorded depth of > 0.8 m (typical body length of adult Common Loons in this study). Because pressure data were collected at 20-sec intervals, we interpolated the times of both dive initiation and surfacing. Maximum daily ascent and descent rates were estimated individually for each loon and used to interpolate times of dive initiation and surfacing, where rates were based on median values derived from dives >35 m (n = 3974 dives of 10 individual loons). The descent and ascent rates used for interpolation were 1.056 m/sec and 1.605 m/sec, respectively, when loons were in freshwater (Kenow et al. 2018), and 0.992 and 1.399 m/sec, respectively, in marine environments (this study). We defined dive bouts as sequential dives that were separated by a post-dive surface interval >360 sec (Kenow et al. 2018). Dive durations of <20 sec may or may not have been detected depending on the position of the loon (surface or underwater) at the time of pressure recording.
We coupled dive profile patterns (from geolocator tag data) of radio-marked loons and bathymetric data associated with their wintering locations to provide insights into vertical foraging patterns. Spot measurements of water depth and distance to shore were determined for each radio-marked loon location. Water depth associated with estimated locations of radio-marked loons was determined using bathymetric data (National Geophysical Data Center 1999a, 1999b, 2001a, 2001b, 2001c) using ArcGIS software (ESRI 2015). Only the most precise of the Argos Location Classes were used in the analysis (i.e., LC 3).
Statistical Analyses
Using dive data collected from geolocator tags, we estimated associations between the dependent variables (dive depth, dive duration, time at maximum depth, and log-transformed post-dive surface interval) and the explanatory variables (location (Atlantic Coast and GOM), sex–radio), and their interaction. The term “sex–radio” denotes a variable with three categories, namely non-radio-marked females, radio-marked males, and non-radio-marked males. We did not treat sex–radio as an interaction term because only males were fitted with radios. The effects of individual birds (“bird effects”) on dive duration, post-dive surface interval, and time at maximum depth were treated as random, while variances of bird effects on dive durations were permitted to vary by winter location. Radio-marking status was considered in the analyses because abdominally-implanted transmitters with percutaneous antennas can potentially have negative effects on dive performance in diving waterbirds (Latty et al. 2010). Models were fitted using restricted maximum likelihood and SAS’s generalized linear mixed-modeling procedure (PROC GLIMMIX; SAS Institute 2019). Code and data are available at https://doi.org/10.5066/P9UIDEMO (Kenow et al. 2021b). Dive depths were analyzed using geotag-specific mean dive depths because dive depths were strongly left-skewed and because sample means of those depths were estimated precisely (i.e., relative standard errors did not exceed 2.4%, given an assumption of independence of dive estimates for each bird). Hence, estimates of mean dive depth from dive depth models represent means of means or, simply, “grand means.”
Reported means represent marginal means for balanced populations, whereas geometric means represent exponentiations of marginal means. Unless otherwise specified, means for post-dive surface intervals represent geometric means. Error rates for multiple comparison tests were addressed using Scheffé’s method (Scheffé 1953, Snedecor and Cochran 1989); error variances were permitted to vary among winter locations.
RESULTS
We captured, banded, and attached geolocator tags to 204 adult Common Loons during summers 2009–2016 (Table 1). Thirty-one of these loons (all male) were radio-marked. We recovered 111 geolocator tags from 134 of the geotagged adult loons that were recaptured or recovered as carcasses (i.e., geolocator tags were missing from 23 of the geotagged loons at the time of recapture). Hence, loon recapture/recovery and geolocator tag retention percentages were 66% and 83%, respectively. Premature geolocator tag failure, data corruption, or tag programming issues resulted in only 61 tags attached to adult loons that provided usable dive data while on wintering areas. Twenty-four of the 31 radio-marked adult male loons were tracked to their wintering areas; one loon died on its breeding lake, one loon’s satellite transmitter failed prior to migration, two loons died during migration, and signals were lost from three loons’ transmitters during migration (Kenow et al. 2021a).
Foraging patterns of wintering adult Common Loons
Dive analyses were based on 176,101 dives by 61 geotagged adult loons while wintering in the GOM, the Atlantic Ocean, or southeastern reservoirs (Table 2). Geolocator tags collected pressure data a mean of 20 days (median = 16; 5th and 95th quantiles: 5, 43) following arrival to wintering areas. The median winter arrival date among geotagged loons was 27 November (7 November, 9 December) and pressure data were collected through a median date of 12 December (5 December, 1 January).
The proportion of time loons were recorded as underwater during the day reflects a pattern of foraging activity during daylight (Fig. 1). The proportion of time underwater appeared to be greater among non-radio-marked loons compared to radio-marked loons. The median hourly proportion of time spent underwater among radio-marked loons during midday (0900 to 1600 h) was 0.30 (5th and 95th quantiles: 0.10, 0.54), compared to median values of 0.44 (0.20, 0.71) for non-radio-marked females, and of 0.38 (0.13, 0.63) for non-radio-marked males.
Common Loon dives were as deep as 26 m in southeastern reservoirs, 34 m in the Atlantic Ocean, and 50 m in the GOM (Fig. 2). Because dives of very few loons were documented in southeastern reservoirs due to a small number of geotagged loons that wintered on these reservoirs, we limited subsequent analyses of dive characteristics to loons wintering in the Atlantic Ocean and the GOM. Distributions of dive depths in the Atlantic Ocean and GOM were multimodal. Grand means of loon-specific mean dive depths appeared similar for all location × sex–radio combinations except for those that reflect radio-marked individuals (Tables 3, 4). Specifically, radio-marked males in the Atlantic Ocean yielded an unusually small grand mean dive depth while those in the GOM yielded a correspondingly large grand mean dive depth. The former grand mean represents the mean of quite different loon-specific means, i.e., 4.5 (0.02 SE), 12.7 (0.03), and 20.0 (0.10) m, whereas the latter reflects eight means that were relatively similar (range: 23.7 to 29.3 m) and a ninth that was not (7.7, SE 0.1). The difference in mean dive depths by location largely reflects these differences in dive depths by radio-marked males.
Mean dive duration (where minimum dive duration was >20 sec) did not clearly differ by sex or location for non-radio-marked loons (Tables 3, 4). A possible exception (and as with maximum dive depths) was that mean dive durations for radio-marked loons appeared lower in the Atlantic Ocean than in the GOM. Dive duration was strongly associated with dive depth in all wintering locations (Fig. 3). Mean durations at maximum depth did not clearly differ by wintering area or sex–radio categories (Tables 3, 4). Mean duration at maximum depth increased with depth of dive in the Atlantic Ocean and GOM (Fig. 4). Mean post-dive surface intervals among location × sex–radio categories were similar, except for that of radio-marked males in the GOM (Tables 3, 4). Mean post-dive surface intervals typically increased with depth of dive (given dives greater than approximately 10 m; Fig. 5).
Common Loon dive shapes were characterized by a relatively long duration at maximum depth (defined as within 2 m of maximum depth) relative to the duration of the entire dive. Such dive shapes are illustrated in a dive bout that occurred in the GOM consisting of 20 dives to a repeated depth of about 28 m (Fig. 6). The dive shape of most dives (93% in the Atlantic Ocean and GOM; 88% in southeastern reservoirs) in our study met the criterion of Schreer et al. (2001) of a “square” shape, where time at maximum depth was ≥50% of the dive duration. The relation between recorded dive depths and estimated water depth among LC3 telemetry location estimates of radio-marked loons in this study provided multiple examples of loons foraging at or near the benthic zone. Of 27 dive-depth records from six geolocator tags that were matched within 15 min of locations, 22 were within 2 m of indicated water depths from bathymetry data.
DISCUSSION
Dive activity results presented here, coupled with wintering distribution results published in a companion article (Kenow et al. 2021a), offer unique insights into the wintering ecology of Common Loons. Archival geolocator tags provided the opportunity to document dive activity of wintering loons without the limitations of shore- or watercraft-based direct observation. Direct observations are generally limited to relatively shallow nearshore waters, do not provide a continuous record for an individual over the entire day, and are difficult to apply to unmarked loons foraging in aggregations. However, direct observations can be more precisely linked to a specific site.
During midday, adult Common Loons monitored in this study were underwater about 40% of the time across wintering areas. Considering the post-dive surface interval as part of foraging activity, roughly 60% of the midday period was spent engaged in foraging activity. In comparison, daily foraging activity of Common Loons wintering off Assateague Island, Virginia, was reported as 55% of activities (McIntyre 1978) and foraging accounted for 23% to 38% of recorded activity among 3 years of observations of loons wintering in coastal waters of Weekapaug, Rhode Island (Daub 1989, Ford and Gieg 1995). Mean dive duration among adult Common Loons foraging in the Atlantic Ocean (104 sec) and GOM (111 sec) exceeded those reported for wintering loons foraging in shallow waters off the Virginia coast (39.5 (SD 1.25) sec; McIntyre 1978), likely due to differences in general foraging depths of loons under direct observation compared to geotagged loons.
We also note that mean dive durations of wintering loons from this study tended to be longer than dive durations observed among the same loons while staging on Lake Michigan (the latter dive durations varied from 69 to 90 sec inclusively; Kenow et al. 2018). This difference in dive duration appears to be related to longer duration at maximum depth among wintering loons and may reflect greater prey availability leading to more efficient foraging while on Lake Michigan. The observed associations of longer dive durations, increasing duration at maximum depth, and increasing post-dive surface intervals with increasing dive depths among wintering Common Loons in this study are consistent with those observations of loons staging on Lake Michigan (Kenow et al. 2018). Longer dive durations with increasing depths have also been observed with other diving bird species (Wilson and Wilson 1988, Kooyman et al. 1992, Wilson et al. 2011) whereas increasing duration at maximum depth with increased dive depth has been documented in diving seabirds (Wilson and Wilson 1988, Wilson et al. 1996). Schreer et al. (2001) described a general increase in post-dive surface interval with increasing dive duration among 12 species of pinnipeds and seabirds. Nocera and Burgess (2002) observed that post-dive surface intervals of breeding Common Loons were not associated with dive duration, concluding that loons in their study were not approaching the aerobic dive limit. However, mean loon dive durations in their study (39.7 ± 9.0 sec) were substantially shorter than dives recorded in this study.
The geolocator tags used in this study were programmed to record pressure at a sampling interval of 20 sec to allow us to monitor dive activity over about four months to ensure we covered the anticipated fall migration period (Kenow et al. 2018). Consequently, limitations on geolocator tag data storage capacity restricted our collection of wintering dive data to November and December. Future efforts to document foraging activity of Common Loons throughout the wintering period would be useful; however, we note that radio-marked adult Common Loons generally did not shift wintering core use areas as winter progressed (Kenow et al. 2021a). Therefore, early winter dive characteristics reported in this study may represent a reasonable expectation of dive characteristics throughout the wintering period, assuming prey abundance does not shift within the water column. Some dives and surfacings <20 sec in duration may have gone unrecorded, resulting in an underestimate of the number of dives. Schreer et al. (2001) indicate that sampling interval may also affect determination of dive shape, where increased sampling intervals resulted in classification of fewer square-shaped dives. Decreasing the sampling interval (i.e., decrease to 5 or 10 sec) would provide better resolution for detecting fine-scale dive patterns (Boyd 1993) but would substantially limit the observation period.
Our results indicate potential effects of radio-marking on the proportion of time underwater (decreased) and post-dive surface interval (increased) compared to non-radio-marked loons. This finding is consistent with that of decreased proportions of time spent underwater by these same loons during their migration stopover at Lake Michigan (Kenow et al. 2018). While dive depth, dive duration, and duration at maximum depth did not vary considerably with radio-marking, the duration of time on the surface between dives tended to be longer during both fall migration and wintering. As most dives among diving mammals and birds are believed to be aerobic (Butler and Jones 1997), surface recovery time is likely related to time required to refuel O2 stores with behavior apparently controlled primarily by blood gas (O2 and CO2) levels (Enstipp et al. 2001). We speculate that the dive recovery period of radio-marked loons following prolonged dives may be related to reduced air-sac volume due to satellite transmitter implants impinging upon a portion of the air sacs. Latty et al. (2010) suggested that loss of air space volume due to transmitter implants may have reduced vertical travel time, aerobic dive limits, and buoyancy in Common Eiders (Somateria mollissima). Although we have no evidence of reduced vitality of radio-marked loons in this study, researchers should be aware of this potential subtle negative effect of intracoelomic transmitter implants on dive activity when considering possible effects of the method as it relates to the objectives of their future studies.
Dive patterns of wintering adults indicate benthic feeding behavior
Recorded dive depths of adult Common Loons marked with geolocator tags, coupled with fine-resolution satellite telemetry location data, indicate that much of winter loon foraging activity occurred along the ocean bottom at depths of up to 50 m (Fig. 7). Dive profile patterns (e.g., Fig. 6) of repeated dives to consistent depth, coupled with time at maximum depth ≥50% of the dive duration (classified as “square” dive shape by Schreer et al. [2001]) are characteristic of benthic feeding (Wilson and Wilson 1988, Halsey et al. 2007, Kenow et al. 2018). Square dives indicate an animal is maximizing the percentage of time at a particular water depth where prey may reside, and descent and ascent to the target depth is rapid (Schreer et al. 2001). In addition to our finding that 93% of recorded dives were characteristic of benthic feeding, a comparison of dive depths matched to indicated water depths of radio-marked loons provided supporting evidence of loons engaged in foraging at or near the benthic zone.
Common Loon winter use areas identified by Kenow et al. (2021a) along the continental shelf of the South Atlantic and West Florida Gulf include extensive scattered areas of reef habitats (Parker et al. 1983) and associated reef fisheries (Fig. 7). Briones (2003) reported that a large proportion of the continental shelf contains soft, muddy sediments with the benthic community dominated by invertebrates and demersal fish. Little is known about the Common Loons' winter diet, especially among loons foraging offshore. However, solitary and group foraging strategies have been observed among nearshore wintering loons (Jodice 1993, Vlietstra 2000). Larger groups (≥50) of loons were associated with assemblages of seabirds and dolphins (Jodice 1993), and a foraging flock of >600 wintering loons was observed in the GOM 17 km off the coast of Mississippi (Long and Paruk 2014). Wintering Common Loons in the GOM and Atlantic Ocean have been observed eating benthic fish species, crabs, winter flounder (Pseudopleuronectes americanus), and starfish (Haney 1990, Ford and Gieg 1995), or foraging in areas where Gulf menhaden (Brevoortia patronus), tidewater silversides (Menidia peninsulae), Atlantic croaker (Micropogonias undulatus), and spot (Leiostomus xanthurus) were abundant (Spitzer 1993, Vlietstra 2000). We speculate that prey is abundant along the continental shelf and that this abundance drives the observed distribution of wintering Common Loons in the GOM.
Implications for conservation of wintering Common Loons
Our results emphasize the importance of offshore habitats for wintering Common Loons, particularly the reef habitats and associated fisheries of the GOM West Florida Shelf that are likely a critical wintering resource for loons breeding in the Upper Midwest (Kenow et al. 2002, this study). The knowledge of loon distribution and habitat use provided here and in Kenow et al. (2021a) could also support Common Loon conservation efforts and inform management of designated protection areas in marine systems (e.g., Florida Middle Grounds Habitat Area of Particular Concern fishery management area).
Documentation of loons foraging within the benthic zone provides insight into a potential mechanism for exposure/transfer of contaminants to Common Loons, may have implications for assessing oil spill injury to loons (e.g., Deepwater Horizon Natural Resource Damage Assessment Trustees 2016), and may help guide clean-up and mitigation efforts related to marine oil spill events (e.g., Open Ocean TIG 2019).
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.
ACKNOWLEDGMENTS
Funding for this study was provided by Great Lakes Restoration Initiative through an interagency agreement with the U.S. Environmental Protection Agency, with additional support from the Legislative-Citizen Commission on Minnesota Resources' Environment and Natural Resources Trust Fund, and U.S. Geological Survey Wildlife Program funds. We thank the numerous volunteers and field biologists who assisted with field logistics in support of this study. L. Naumann, R. Baker, P. Perry, and staff with the Minnesota Department of Natural Resources lined up local cooperators and provided necessary Minnesota permits. Local field support was provided by W. Brininger and L. Deede (Tamarac National Wildlife Refuge), L. Laske and S. Maanum (Big Mantrap Lake Association), M. Hintz (Sportsmen's Club of Lake Vermilion, Inc.), R. Mack (Burntside Property Owners' Association), and J. Zernov., P. Boma, L. Naumann, J. Wilson, T. Daulton, D. Killian, D. Stockwell, K. Woizeschke, K. Carlyle, and J. Kohn assisted with capture work. The surgical procedures were conducted by D. Heard (College of Veterinary Medicine, University of Florida, Gainesville, Florida. K. Carlyle assisted with transmitter implant surgery. K. Bond, L. Browning, J. Dunn, R. Eckert, M. Goodman, S. Hillen, M. Hintz, K. Jackson, L. Martinelli, D. Newstead, P. Parker, S. Schweitzer, J. Tunnel, K. Westlake, H. Wilcox, and S. Wright assisted with recoveries of loon carcasses. C. Churchill and B. Uher-Koch provided constructive reviews of an earlier draft of this report. Movement data from the radio-marked loons were served on a website established to provide project information to partners and the public (https://umesc.usgs.gov/terrestrial/migratory_birds/loons/migrations.html). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
DATA AVAILABILITY
The data/code that support the findings of this study are openly available in ScienceBase at https://doi.org/10.5066/P9UIDEMO. Ethical approval for this research study was granted by the Animal Care and Use Committee of the USGS Upper Midwest Environmental Sciences Center.
LITERATURE CITED
Argos 2016. Argosweb user manual. CLS America, Inc., Lanham, Maryland, USA. http://www.argos-system.org/wp-content/uploads/2016/09/ArgosWeb_User_Manual.pdf
Boyd, I. L. 1993. Selecting sampling frequency for measuring diving behavior. Marine Mammal Science 9:424-430. https://doi.org/10.1111/j.1748-7692.1993.tb00475.x
Briones, E. E. 2003. Current knowledge of benthic communities in the Gulf of Mexico. Pages 108-136 in K. Withers and M. Nippers, editors. Environmental Analysis of the Gulf of Mexico. Texas A&M University, Corpus Christi, Texas, USA.
Butler, P. J., and D. R. Jones. 1997. Physiology of diving of birds and mammals. Physiological Reviews 77:837-899. https://doi.org/10.1152/physrev.1997.77.3.837
Daub, B. C. 1989. Behavior of Common Loons in winter. Journal of Field Ornithology 60:305-311.
Deepwater Horizon Natural Resource Damage Assessment Trustees. 2016. Deepwater Horizon oil spill: Final programmatic damage assessment and restoration plan and final programmatic environmental impact statement. http://www.gulfspillrestoration.noaa.gov/restoration-planning/gulf-plan
Ekstrom, P. A. 2004. An advance in geolocation by light. Memoirs of National Institute of Polar Research, Special Issue 58:210-226.
Enstipp, M. R., R. D. Andrews, and D. R. Jones. 2001. The effects of depth on the cardiac and behavioural responses of double-crested cormorants (Phalacrocorax auritus) during voluntary diving. Journal of Experimental Biology 204:4081-4092. https://doi.org/10.1242/jeb.204.23.4081
ESRI. 2015. ArcGIS Desktop: Release 10.3.1 Redlands. Environmental Systems Research Institute, CA, USA.
Evers, D. C. 1993. A replicable capture method for adult and juvenile common loons on their nesting lakes. Pages 214-220 in L. S. Morse, S. Stockwell, and M. Pokras, editors. Proceedings from the Bar Harbor, ME, USA 1992 conference on the loon and its ecosystem: Status, management and environmental concerns. U.S. Fish and Wildlife Service.
Fofonoff, N. P., and R. C. Millard, Jr. 1983. Algorithms for computation of fundamental properties of seawater. Tech. Rep 44, UNESCO, Technical Papers in Marine Science.
Ford, T. B., and J. A. Gieg. 1995. Winter behavior of the Common Loon. Journal of Field Ornithology 66:22-29. https://www.jstor.org/stable/4513976
Fox, T. J. 2018. Composite raster and divergence tool. U.S. Geological Survey code release.https://doi.org/10.5066/F7ZC8239
Halsey, L. G., C. A. Bost, and Y. Handrich. 2007. A thorough and quantified method for classifying seabird diving behaviour. Polar Biology 30:991-1004. https://doi.org/10.1007/s00300-007-0257-3
Haney, J. C. 1990. Winter habitat of common loons on the continental shelf of the southeastern United States. Wilson Bulletin 102:253-263.
Jodice, P. G. R. 1993. Distribution of wintering loons in the northeastern Gulf of Mexico. Pages 172-193 in L. S. Morse, S. Stockwell, and M. Pokras, editors. Proceedings from the Bar Harbor, ME, USA 1992 conference on the loon and its ecosystem: Status, management, and environmental concerns. U.S. Fish and Wildlife Service.
Kenow, K. P. 2018. SAS Code for Extracting Dive Profile Information from Archival Geolocator Tags. U.S. Geological Survey code release, https://doi.org/10.5066/F7VT1R95
Kenow, K. P., L. J. Fara, S. C. Houdek, B. R. Gray, D. J. Heard, M. W. Meyer, T. J. Fox, R. J. Kratt, S. L. Ford, A. Gendron-Fitzpatrick, and C. L. Henderson. 2021a. Migration patterns and wintering distribution of Common Loons breeding in the Upper Midwest. Journal of Avian Biology e02609. https://doi.org/10.1111/jav.02609
Kenow, K. P., L. J. Fara, S. C. Houdek, and B. R. Gray. 2021b. Common loon winter data: telemetry locations and archival geolocator tag location and dive data. U.S. Geological Survey data release. https://doi.org/10.5066/P9UIDEMO
Kenow, K. P., S. C. Houdek, L. J. Fara, B. R. Gray, B. R. Lubinski, D. J. Heard, M. W. Meyer, T. J. Fox, and R. J. Kratt. 2018. Distribution and foraging patterns of common loons on Lake Michigan with implications for exposure to type E avian botulism. Journal of Great Lakes Research 44:497-513. https://doi.org/10.1016/j.jglr.2018.02.004
Kenow, K. P., M. W. Meyer, D. C. Evers, D. C. Douglas, and J. Hines. 2002. Use of satellite telemetry to identify common loon migration routes, staging areas and wintering range. Waterbirds 25:449-458. https://doi.org/10.1675/1524-4695(2002)025[0449:UOSTTI]2.0.CO;2
Kenow, K. P., J. M. Wilson, and M. W. Meyer. 2009. Capturing common loons during prenesting and nesting periods. Journal of Field Ornithology 80:427-432. https://doi.org/10.1111/j.1557-9263.2009.00250.x
Kooyman, G. L., Y. Cherel, Y. Le Maho, J. P. Croxall, P. H. Thorson, V. Ridoux, and C. A. Kooyman. 1992. Diving behavior and energetics during foraging cycles in King Penguins. Ecological Monographs 62:143-163. https://doi.org/10.2307/2937173
Korschgen, C. E., K. P. Kenow, A. Gendron-Fitzpatrick, W. L. Green, and F. J. Dein. 1996. Implanting intra-abdominal radiotransmitters with external whip antennas in ducks. Journal of Wildlife Management 60:132-137. https://doi.org/10.2307/3802047
Latty, C. J., T. E. Hollmen, M. R. Petersen, A. N. Powell, and R. D. Andrews. 2010. Abdominally implanted transmitters with percutaneous antennas affect dive performance of common eiders. Condor 112:314-322. https://doi.org/10.1525/cond.2010.090022
Long, D., and J. D. Paruk. 2014. Unusually large wintering flock of common loons foraging in the Gulf of Mexico. Southeastern Naturalist 13:49-51. https://doi.org/10.1656/058.013.0406
McIntyre, J. W. 1978. Wintering behavior of common loons. Auk 95:396-403.
National Geophysical Data Center. 1999a. U.S. Coastal Relief Model - Northeast Atlantic. National Geophysical Data Center, NOAA. https://doi.org/10.7289/V5MS3QNZ
National Geophysical Data Center. 1999b. U.S. Coastal Relief Model - Southeast Atlantic. National Geophysical Data Center, NOAA. https://doi.org/10.7289/V53R0QR5
National Geophysical Data Center. 2001a. U.S. Coastal Relief Model - Florida and East Gulf of Mexico. National Geophysical Data Center, NOAA. https://doi.org/10.7289/V5W66HPP
National Geophysical Data Center. 2001b. U.S. Coastal Relief Model - Central Gulf of Mexico. National Geophysical Data Center, NOAA. https://doi.org/10.7289/V54Q7RW0
National Geophysical Data Center, 2001c U.S. Coastal Relief Model - Western Gulf of Mexico. National Geophysical Data Center, NOAA. https://doi.org/10.7289/V5QJ7F79
Nocera, J. J., and N. M. Burgess. 2002. Diving schedules of Common Loons in varying environments. Canadian Journal of Zoology 80:1643-1648. https://doi.org/10.1139/z02-157
Open Ocean TIG. 2019. Open Ocean Trustee Implementation Group Final Restoration Plan 1 and Environmental Assessment: Birds and Sturgeon. Document Number: 2019-05379.
Parker, R. O. Jr., D. R. Colby, and T. D. Willis. 1983. Estimated amount of reef habitat on a portion of the U.S. South Atlantic and Gulf of Mexico Continental Shelf. Bulletin of Marine Science 33:935-940.
Paruk, J. D., D. C. Evers, J. W. McIntyre, J. F. Barr, J. Mager, and W. H. Piper. 2021. Common Loon (Gavia immer), version 2.0., in P. G. Rodewald and B. K. Keeney, editors. Birds of the World. Cornell Lab of Ornithology. https://doi.org/10.2173/bow.comloo.02
SAS Institute, Inc. 2019. SAS/STAT 15.1 user’s guide - procedures. SAS Institute Inc., Cary, NC, USA. https://support.sas.com/documentation/onlinedoc/stat/indexproc.html
Scheffé, H. 1953. A method for judging all contrasts in the analysis of variance. Biometrika 40:87-104. https://doi.org/10.2307/2333100
Schreer, J. F., K. M. Kovacs, and R. J. O'Hara Hines. 2001. Comparative diving patterns of pinnipeds and seabirds. Ecological Monographs 71:137-162. https://doi.org/10.1890/0012-9615(2001)071[0137:CDPOPA]2.0.CO;2
Sea-Bird. 2002. Conversion of pressure to depth. Application note no. 69. Sea-Bird Electronics, Inc., Bellevue, WA, USA. https://www.seabird.com/quick.search-download.search.jsa?keywords=application+notes
Snedecor, G. W., and W. G. Cochran. 1989. Statistical Methods. Iowa State University Press, Ames, IA, USA.
Spitzer, P. R. 1993. Processes that shape the distribution, abundance, and mortality of common loons during migration and winter with examples drawn from the mid-Atlantic Coastal Region (New Jersey to South Carolina). Page 171 in L. S. Morse, S. Stockwell, and M. Pokras, editors. Proceedings from the Bar Harbor, ME, USA, 1992 conference on the loon and its ecosystem: status, management and environmental concerns. U.S. Fish and Wildlife Service.
Spitzer, P. R. 1995. Common loon mortality in marine habitats. Environmental Review 3:223-229. https://doi.org/10.1139/a95-011
Vlietstra, L. S. 2000. Local variation in foraging strategies employed by wintering common loons. Pages 25-34 in J. W. McIntyre and D. C. Evers, editors. Loons: Old history and new findings. Proceedings of a Symposium from the 1997 meeting, American Ornithologists' Union. North American Loon Fund, Holderness, NH, USA.
Wilson, R. P., B. M. Culik, G. Peters, and R. Bannasch. 1996. Diving behaviour of Gentoo penguins, Pygoscelis papua; factors keeping dive profiles in shape. Marine Biology 126:153-162. https://doi.org/10.1007/BF00571387
Wilson, R. P., C. R. McMahon, F. Quintana, E. Frere, A. Scolaro, G. C. Hays, and C. J. A. Bradshaw. 2011. N-dimensional animal energetic niches clarify behavioural options in a variable marine environment. Journal of Experimental Biology 214:646-656. https://doi.org/10.1242/jeb.044859
Wilson, R. P., and M. T. Wilson. 1988. Foraging behaviour in four sympatric cormorants. Journal of Animal Ecology 57:943-955. https://doi.org/10.2307/5103
Fig. 1
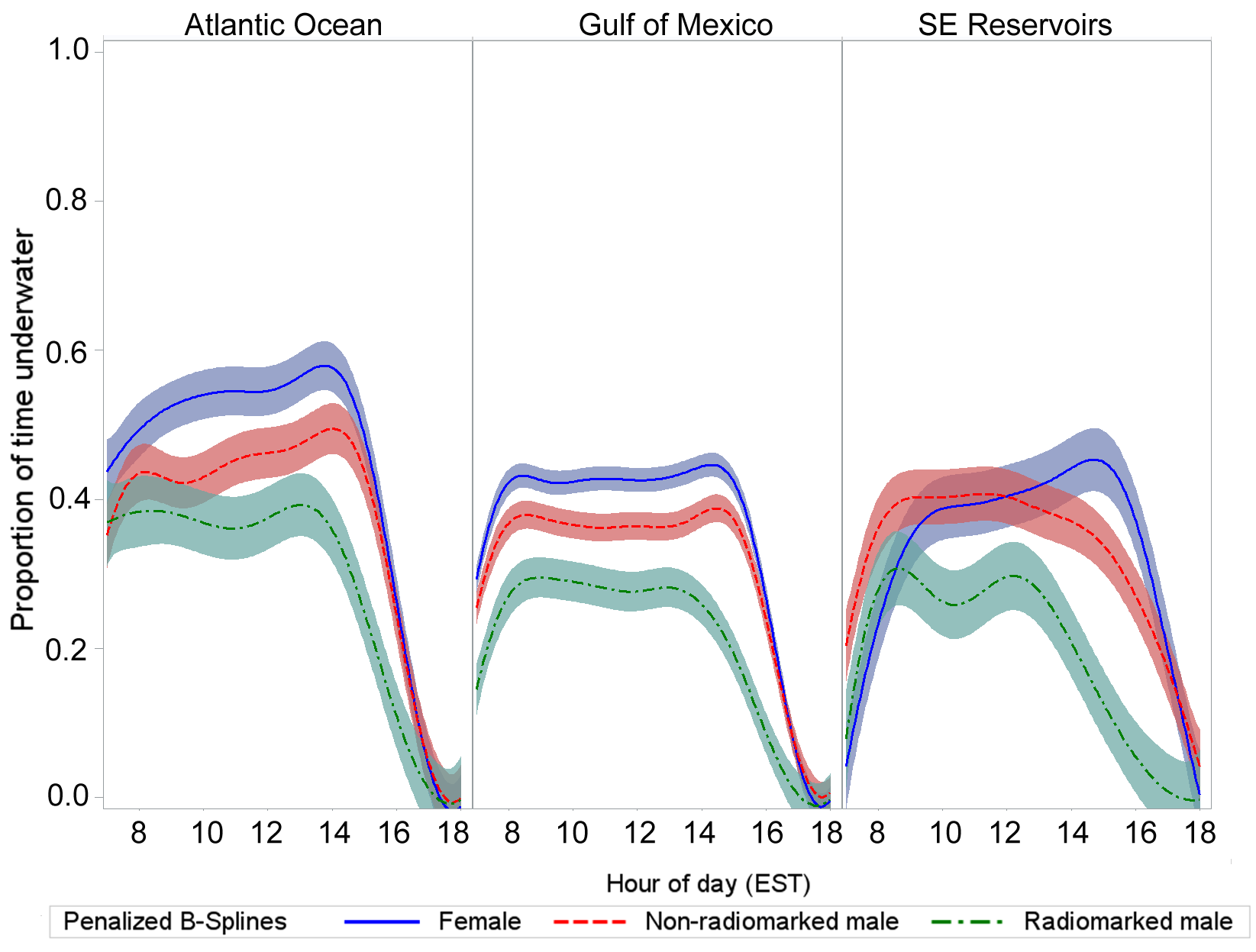
Fig. 1. Proportion of time underwater by location and time of day illustrated using dive data from geotagged Common Loons (Gavia immer; Table 2 for sample sizes) while using wintering areas. Penalized B-spline curves with 95% confidence bands are depicted for radio-marked male (green), non-radio-marked male (red), and female (blue) loons.
Fig. 2
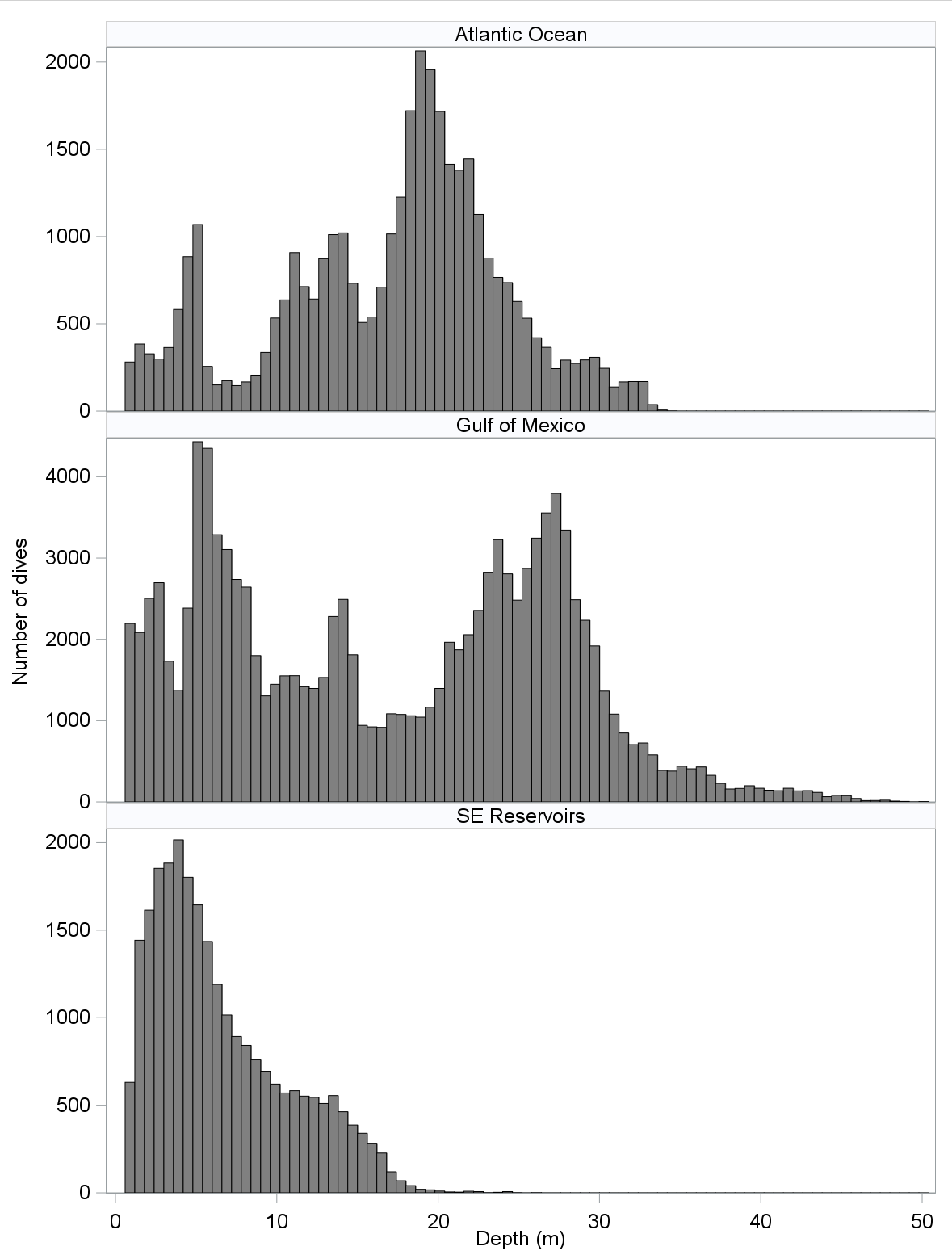
Fig. 2. Distribution of dive depths by wintering area determined using dive data from 61 geotagged Common Loons (Gavia immer; Table 2 for sample sizes).
Fig. 3
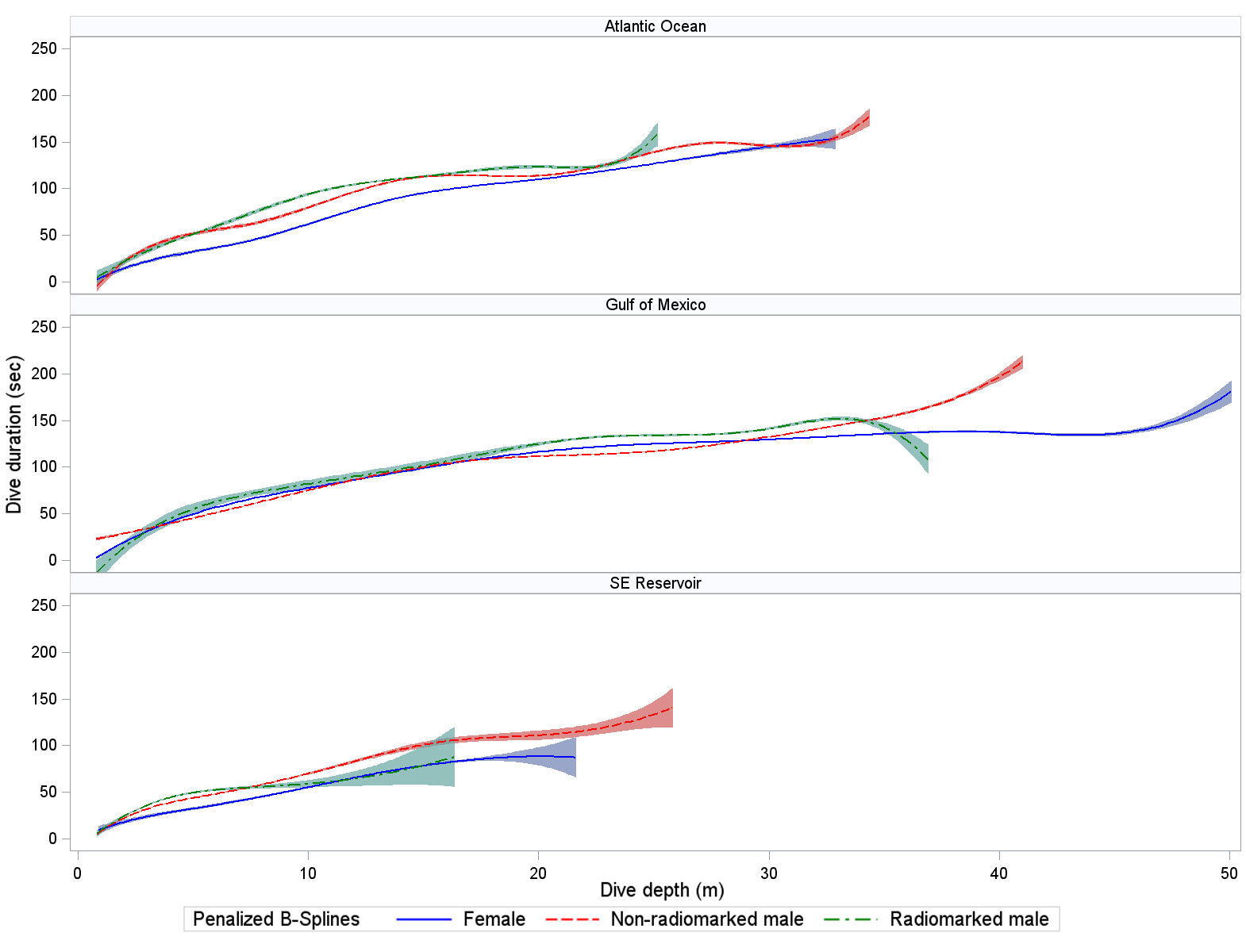
Fig. 3. Relationship between Common Loon (Gavia immer) dive duration (sec) and depth of dive (m) while using wintering areas (Table 2 for sample sizes). Penalized B-spline curves with 95% confidence bands are depicted for radio-marked male (green), non-radio-marked male (red), and female (blue) loons. Confidence bands do not adjust for correlation of measurements within birds.
Fig. 4
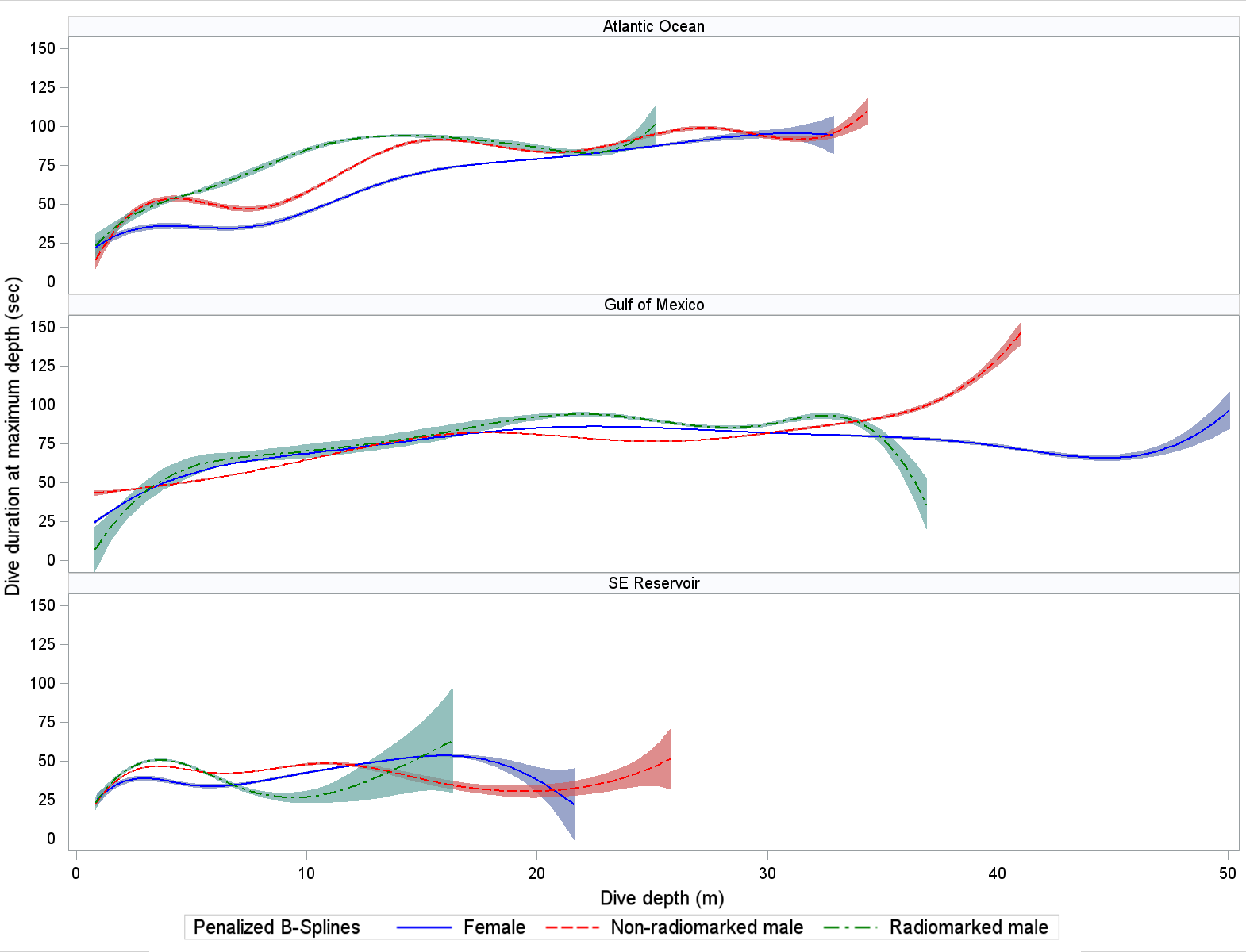
Fig. 4. Relationship between Common Loon (Gavia immer) duration of dive within 2 m of maximum depth (sec) and depth of dive (m) while using wintering areas (Table 2 for sample sizes). Penalized B-spline curves with 95% confidence bands are depicted for radio-marked male (green), non-radio-marked male (red), and female (blue) loons. Confidence bands do not adjust for correlation of measurements within birds.
Fig. 5
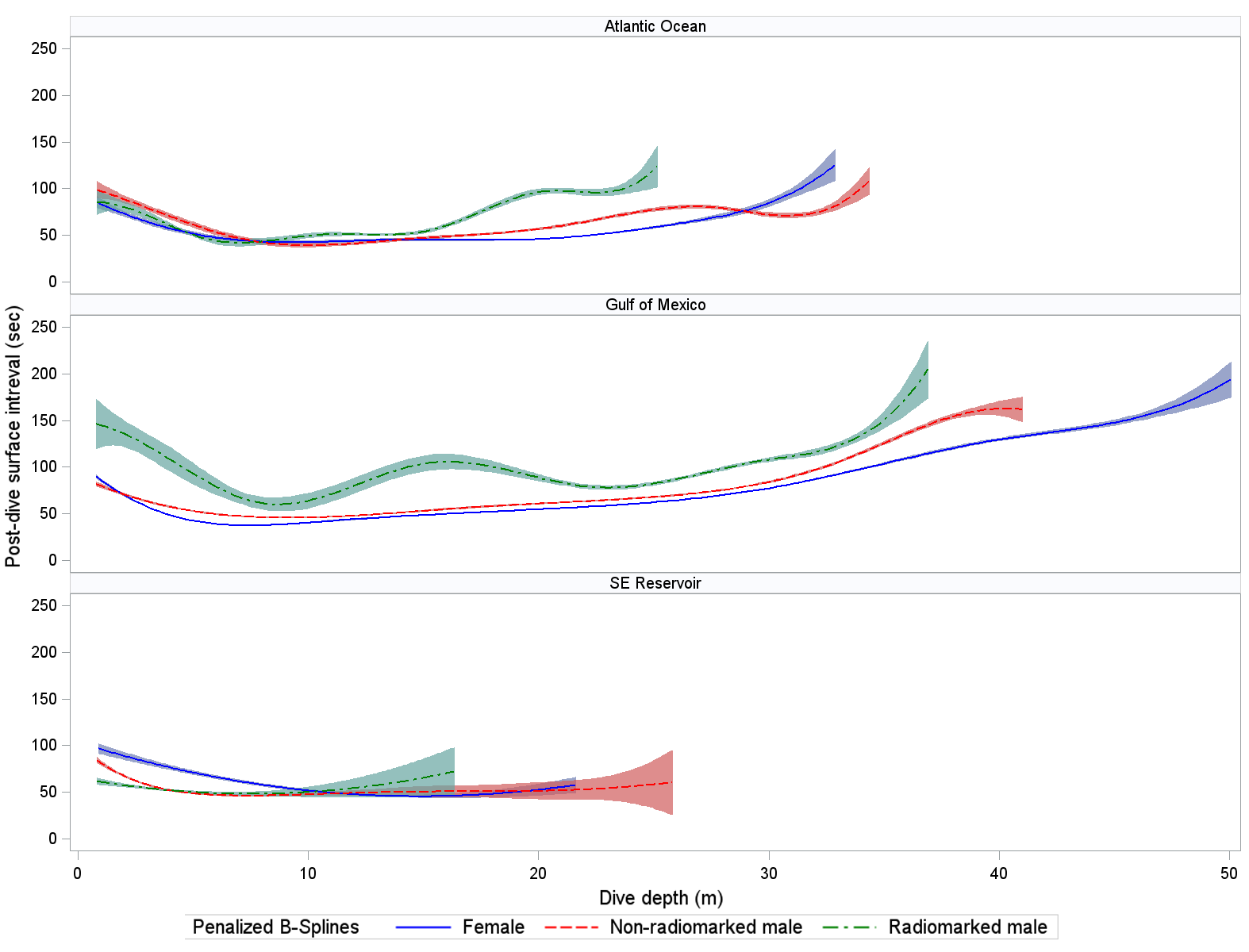
Fig. 5. Relation between Common Loon (Gavia immer) post-dive surface interval (sec) and depth of dive (m) while using wintering areas (Table 2 for sample sizes). Penalized B-spline curves with 95% confidence bands are depicted for radio-marked male (green), non-radio-marked male (red), and female (blue) loons. Confidence bands do not adjust for correlation of measurements within birds.
Fig. 6
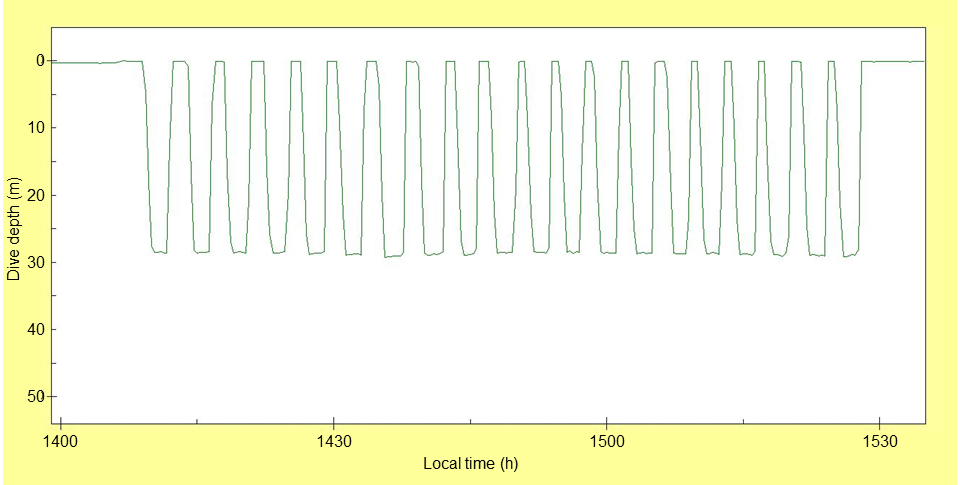
Fig. 6. Dive profile of a geotagged Common Loon (Gavia immer) (ID 1038-94241) collected with an archival geolocator tag while wintering in the Gulf of Mexico. This dive bout consisted of 20 repeated dives during the afternoon of 09 December 2010. Bathymetric data indicates water depth of 28 m for the approximate location of this loon (latitude = 28.583 degrees, longitude = -83.766 degrees) during the dive bout illustrating benthic foraging in the figure.
Fig. 7
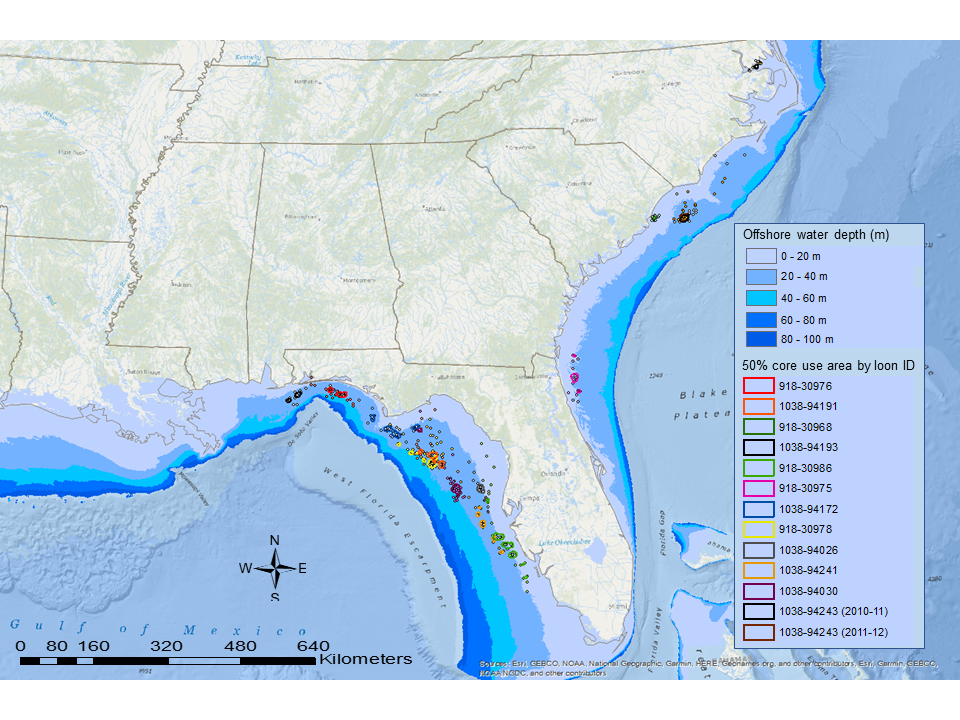
Fig. 7. Oceanic wintering locations and 50% core use areas of 12 adult male Common Loons (Gavia immer) radio-marked on their breeding territories in Minnesota, Wisconsin, and the Upper Peninsula of Michigan, 2010–2011. Location data were collected throughout the wintering period (i.e., November/December through March/April). Loon IDs are federal leg-band numbers. Loons were captured and radio-marked on breeding lakes in Minnesota (918-30976, 918-30986, 918-30975, 1038-94241), Wisconsin (1038-94172, 918-30978, 1038-94026, 1038-94030, 1038-94243), and the Upper Peninsula of Michigan (1038-94191, 918-30968, 1038-94193). (Base layer source: “North America.” ~1:72k. “World Ocean Base.” Updated February 22, 2022. http://goto.arcgisonline.com/maps/Ocean/World_Ocean_Base)
Table 1
Table 1. Number of adult Common Loons (Gavia immer), by sex, radiomarked with satellite transmitters and geotagged with archival geolocator tags in Minnesota, Wisconsin, and Michigan during 2009-2016 and subsequent geolocator tag recoveries.
| Number of adult loons | Recaptures-recoveries of geolocator-tagged loons/geolocator tags recovered | Number of geolocator records used in adult wintering area dive analyses | |||||
| Year | Radio-marked | Geotagged | |||||
| Male | Male | Female | ND† | Male | Female | ||
| 2009 | 0 | 9 | 9 | 0 | 14/13 | 0 | 0 |
| 2010 | 10 | 42 | 37 | 0 | 52/46 | 18 | 17 |
| 2011 | 21 | 24 | 13 | 0 | 23/16 | 10 | 0 |
| 2012 | 0 | 21 | 18 | 3 | 25/19 | 7 | 9 |
| 2014 | 0 | 1 | 0 | 0 | 0/0 | 0 | 0 |
| 2015 | 0 | 13 | 11 | 0 | 17/15 | 0 | 0 |
| 2016 | 0 | 1 | 2 | 0 | 3/2 | 0 | 0 |
| Total | 31 | 111 | 90 | 3 | 134/111 | 35 | 26 |
| † ND = not determined. | |||||||
Table 2
Table 2. Common Loon (Gavia immer) archival geolocator dive data sample sizes (number of individual loons; number of dives) by location and sex-radio categories. Dive analyses were based on dive activity of 61 geotagged loons using the Atlantic Ocean and Gulf of Mexico.
| Location | Sex–radio category | Total | |||||
| Females | Non-radio-marked males |
Radio-marked males | |||||
| SE Reservoirs† | 1; 6949 | 1; 15,376 | 1; 3343 | 3; 25,668 | |||
| Atlantic Ocean | 6; 18,059 | 4; 11,646 | 3; 6478 | 13; 36,183 | |||
| Gulf of Mexico | 19; 62,608 | 17; 45,011 | 9; 6631 | 45; 114,250 | |||
| Total | 26; 86,098 | 22; 73,551 | 13; 16,452 | 61; 176,101 | |||
| † SE Reservoirs included Lake Cumberland, Russell County, Kentucky, an unknown reservoir in Alabama, and an unknown reservoir in Georgia. | |||||||
Table 3
Table 3. Estimated mean dive depth, dive duration, duration at maximum depth, and post-dive surface interval of geotagged Common Loons (Gavia immer) among wintering areas, sex-radio category and interactions, 2009–2012. Analyses were based on data from 19 female, 17 non-radiomarked male, and 9 radiomarked male loons in the Gulf of Mexico, and 6 female, 4 non-radiomarked male, and 3 radiomarked male loons in the Atlantic Ocean.
| Variable | Dive depth (m; 95% CI) |
Dive duration (sec; 95% CI) |
Duration at maximum depth (sec; 95% CI) | Duration of post-dive interval (sec; 95% CI) |
| Atlantic Ocean | 16.9 (14.3, 19.5) | 104.0 (90.7, 117.3) | 79.9 (71.9, 87.9) | 53.2 (44.0, 64.3) |
| Gulf of Mexico | 21.0 (18.7, 23.2) | 111.1 (103.0, 119.2) | 78.8 (74.4, 83.1) | 64.7 (58.3, 71.7) |
| Female | 19.5 (17.0, 21.9) | 104.6 (93.5, 115.7) | 76.5 (70.0, 83.0) | 53.2 (45.6, 62.1) |
| Non-radio-marked male | 19.1 (16.2, 21.9) | 107.3 (94.2, 120.3) | 78.6 (70.9, 86.3) | 55.4 (46.1, 66.4) |
| Radio-marked male | 18.2 (14.7, 21.7) | 110.8 (95.0, 126.6) | 82.9 (73.7, 92.1) | 68.5 (55.0, 85.2) |
| Atlantic Ocean × Female | 19.4 (15.8, 23.1) | 108.5 (89.7, 127.2) | 79.3 (68.0, 90.6) | 50.8 (38.9, 66.5) |
| Atlantic Ocean × Non-radio-marked male | 18.9 (14.4, 23.3) | 113.3 (90.3, 136.3) | 84.1 (70.3, 97.9) | 52.5 (37.8, 72.9) |
| Atlantic Ocean × Radio-marked male | 12.4 (7.3, 17.5) | 90.2 (63.7, 116.7) | 76.4 (60.4, 92.3) | 56.4 (38.6, 82.4) |
| Gulf of Mexico × Female | 19.5 (16.2, 22.8) | 100.7 (89.0, 112.5) | 73.8 (67.4, 80.1) | 55.7 (48.0, 64.8) |
| Gulf of Mexico × Non-radio-marked male | 19.3 (15.8, 22.8) | 101.3 (88.8, 113.7) | 73.1 (66.4, 79.8) | 58.4 (49.8, 68.5) |
| Gulf of Mexico × Radio-marked male | 24.1 (19.3, 28.9) | 131.4 (114.2, 148.5) | 89.4 (80.2, 98.7) | 83.1 (66.7, 103.5) |
Table 4
Table 4. Differences among mean dive depth, dive duration, and duration at maximum depth means; and ratios of geometric means of post-dive surface intervals of geotagged Common Loons (Gavia immer) among sex-radio categories among wintering areas, 2009-2012. “*” denotes differences that are significantly different at p < 0.05 (adjusted for multiple comparisons within, but not across, dive characteristics). Analyses were based on data from 19 female, 17 non-radiomarked male, and 9 radiomarked male loons in the Gulf of Mexico, and 6 female, 4 non-radiomarked male, and 3 radiomarked male loons in the Atlantic Ocean.
| Differences among means | Ratios of geometric means |
||||
| Variable | Dive depth (m; 95% CI) |
Dive duration (sec; 95% CI) |
Duration at maximum depth (sec; 95% CI) | Duration of post-dive interval (ratio; 95% CI) | |
| Atlantic Ocean versus Gulf of Mexico | -4.1 (-7.5, -0.6)* | -7.1 (-22.7, 8.4) | 1.1 (-8.0. 10.2) | 0.82 (0.66, 1.02) | |
| Female versus non-radio-marked male | 0.4 (-4.3, 5.1) | -2.7 (-24.2, 18.8) | -2.1 (-14.7, 10.6) | 0.96 (0.71, 1.30) | |
| Female versus radio-marked male | 1.3 (-4.1, 6.6) | -6.2 (-30.4, 18.0) | -6.4 (-20.5, 7.8) | 0.78 (0.56, 1.09) | |
| Non-radio-marked male versus radio-marked male |
0.8 (-4.8, 6.5) | -3.5 (-29.2, 22.2) | -4.3 (-19.4, 10.8) | 0.81 (0.57, 1.16) | |
| Female | |||||
| Atlantic Ocean versus Gulf of Mexico |
7.8 (-8.6, 8.3) | 7.8 (-30.4, 46.0) | 5.5 (-16.8, 27.8) | 0.91 (0.54, 1.55) | |
| Non-radio-marked male | |||||
| Atlantic Ocean versus Gulf of Mexico |
-0.4 (-10.2, 9.3) | 12.0 (-33.1, 57.0) | 11.0 (-15.5, 37.5) | 0.90 (0.48, 1.69) | |
| Radio-marked male | |||||
| Atlantic Ocean versus Gulf of Mexico | -11.7 (-23.8, 0.4) | -41.2 (-95.6, 13.3) | -13.1 (-44.9, 18.8) | 0.68 (0.32, 1.44) | |