







The following is the established format for referencing this article:
Contina, A., A. K. Pierce, S. W. Yanco, E. S. Bridge, J. F. Kelly, and M. B. Wunder. 2022. Application of stable isotopes to study movement ecology and diet variation in a migratory songbird. Journal of Field Ornithology 93(4):6.ABSTRACT
Whether and how migratory organisms exhibit interindividual behavioral and/or physiological variation across movement strategies remains an open question. The Dark-eyed Junco (Junco hyemalis) is a migratory songbird known for its intraspecies variation displayed in relation to morphology, song repertoires, and migration. Thus, studies focusing on juncos can reveal how migratory strategy may covary with other individual-scale factors and, therefore, identify the selective forces driving intraspecies variations throughout its distribution. We used Dark-eyed Junco hydrogen stable isotope feather values (δ²H) and implemented a Bayesian framework to infer the breeding and molting origin of migratory juncos captured on their winter grounds in Oklahoma, USA. We modeled the distribution of feather hydrogen stable isotope values as a function of five morphological variables including body mass and fat deposition measured in Oklahoma during the winter. We then investigated the trade-off between longer and more energetically costly migration strategies, in relation to diet preferences through carbon (δ13C) and nitrogen (δ15N) stable isotope analysis, from feather values. Dark-eyed Juncos wintering in south central U.S. likely originate from multiple breeding populations in northern U.S. and Canada. Body condition at the wintering ground (e.g., mass) had no effect on feather hydrogen stable isotope abundance. However, we found a positive correlation between nitrogen and hydrogen stable isotopes, suggesting that a trophic level shift toward insect consumption might occur in individuals migrating from southern latitudes. Increased insect-derived protein consumption might be explained by reduced fatty-acid reserves necessary to complete a shorter migratory journey.RESUME_
INTRODUCTION
Animal migration is a multifaceted life-history trait that evolved across taxa through complex behavioral and physiological adaptations (Alerstam and Bäckman 2018). Migratory species maintain energetic balance between fuel consumption (e.g., fatty acid catabolism) and replenishing resources to effectively perform a long-distance migration (Guglielmo 2018). However, research into how migrants sustain themselves during migration remains particularly challenging because of the small body size of many migratory songbirds and the fact that they travel, in some cases thousands of kilometers per day (DeLuca et al. 2015, Rani et al. 2017). Species have developed unique behavioral strategies associated with seasonal movements (Horton et al. 2016) that depend on resource availability and seasonal environmental stochasticity. Thus, an understanding of these strategies requires examination of fluctuations in body mass and fat deposition in conjunction with behavior across the spatial extent of the species’ distribution. This is relevant because variation in migration and foraging behavior, at the intraspecies level, can place different evolutionary pressures on individual physiology and act as selective forces across populations. That is, a trade-off may exist between longer and more energetically costly migratory routes, potentially resulting in decreased body condition, and alternative strategies involving diet changes or morphological adaptations (e.g., longer or pointed wings) to accommodate the increased energetic demands of their migratory journey (Kaboli et al. 2007, Reif et al. 2016).
Many species of songbirds are too small to be fitted with tracking devices (e.g., satellite transmitters) and information on their migratory patterns remain unknown. However, indirect molecular approaches can be implemented to study large-scale movements without the use of extrinsic tracking devices. (Rundel et al. 2013). In particular, movement ecology studies have exploited the latitudinal gradient in stable hydrogen isotopes occurring across North America to address questions related to avian migratory patterns and connectivity between breeding and wintering grounds (Rubenstein and Hobson 2004, Vander Zanden et al. 2018, Besozzi et al. 2021). Stable isotope ratios of hydrogen obtained from inert animal tissues (e.g., claws, hair, and feathers) provide insight on the environmental conditions and latitudinal gradient where the keratin-based tissues were grown and offer a valid molecular tool to infer large-scale animal movements in species too small for direct movement observations (Hobson 2005). Importantly, most songbirds replace their flight feathers at the breeding ground before starting fall migration to minimize the energetic costs of the journey (Barta et al. 2008). By replacing old wing feathers with new ones, presumably in better conditions, migratory birds gain aerodynamic efficiency (Bowlin and Wikelski 2008). Thus, feather samples collected at the wintering grounds, sometimes thousands of kilometers away from the breeding locations, can be used to infer where the feathers were grown based on stable isotope similarities between environmental and tissue hydrogen values (Wunder 2010).
A suitable study system for disentangling different migratory strategies from other key evolutionary adaptations is offered by the Dark-eyed Junco (Junco hyemalis), a migratory songbird comprising numerous subspecies broadly distributed across North America. It is found breeding and molting in the northern regions of Canada and Alaska and in the mountainous regions of the United States (U.S.) and wintering in the lower latitudes of Canada and the majority of the continental U.S. and northern Mexico (Nolan et al. 2002). This species has shown negative population trends of over 40% in some areas in the last decade (Conservation Assessment Database 2017, Rockwell et al. 2017) and its physiology and migratory behaviors have been the focus of numerous studies (Nolan and Ketterson 1983, Rogers et al. 1994, Fudickar et al. 2016, Liebgold et al. 2019).
Bridge et al. (2010) analyzed hydrogen isotope ratios from secondary feathers in a population of Dark-eyed Juncos, all from the same hyemalis subspecies group (slate-colored), sampled during the winter of 2009 in central Oklahoma (U.S.) and showed isotopic variation ranging from -175.6‰ to -118.6‰ from Vienna Standard Mean Ocean Water (VSMOW). This large range of variation (~60‰) in hydrogen stable isotope values prompted the authors to speculate that individuals from a wide geographic range of breeding and consequently molting locations wintered in Oklahoma. However, the study did not include explicit probabilistic geographic assignments of wintering individuals, nor did it consider their variation in trophic level.
Although the subspecies of Dark-eyed Junco’s are primarily differentiated by morphology and/or geographic distribution, considerable behavioral diversity between and even within subspecies presents a unique opportunity to address large-scale spatial and ecological questions. We used hydrogen stable isotope data to estimate breeding location origins of a population of Dark-eyed Juncos wintering in Oklahoma, and we collected a suite of morphological measurements after the birds completed fall migration. Moreover, we expect that Oklahoma wintering birds originating from northern latitudes would have a higher carbohydrate intake, but lower protein consumption compared to wintering birds from southern latitudes because of their need to fuel a longer migratory journey. To test this hypothesis, we assessed premigratory diets by examining carbon and nitrogen stable isotope ratios in flight feathers. In addition to quantifying geographic connectivity in the Dark-eyed Junco population wintering in Oklahoma, we offer new insights into how this species negotiates trade-offs between migration distance and diet. Importantly, we present a framework to identify possible confounding effects in explanatory models for different migration strategies across populations (Yanco et al. 2020).
METHODS
Study area
We studied Dark-eyed Juncos wintering in central Oklahoma where the species forages in open woodland habitats and feeds on grass sprouts, Taraxacum, species and various species of insects (Bridge et al. 2010). The weather conditions during our study (January and February 2009), as recorded by the Oklahoma climatological survey and the Oklahoma mesonet (www.mesonet.org) at the Norman (NRMN) weather station, indicated monthly average temperatures ranging from 2.7 to 8.9°C, monthly average precipitation ranging from 22.8 to 24.6 mm, and monthly average wind speeds ranging from 15.6 to 18.1 kph.
Sample collection and stable isotope analysis
We used published stable hydrogen isotope data from 80 Dark-eyed Juncos sampled in central Oklahoma, USA (35°11′0.6″ N, -97°26′44.88″ W) during winter 2009 (Bridge et al. 2010). After removing individuals with missing capture information, our data set comprised stable hydrogen isotope values of secondary feathers from 76 migrants that we used to infer geographic breeding and molting origin. We also presented novel carbon and nitrogen stable isotope data obtained from a subset (N = 74) of those migrants sampled in Oklahoma that we used to study habitat use and trophic level. However, after filtering out individuals with missing morphometric measurements, we had data from 62 migrants (39 adult males and 23 adult females) that we used for morphological models. All samples were collected between January and February of the same year. Feathers were prepared for analysis of stable isotope ratios of hydrogen (δ²H), carbon (δ13C), and nitrogen (δ15N) by following standard washing protocol in a 2:1 solution of chloroform-methanol to remove debris and oil contaminants (Paritte and Kelly 2009, Chew et al. 2019). Samples for hydrogen stable isotope ratios analysis were processed through a Thermo-Finnigan Delta V isotope ratio mass spectrometer interfaced with a high-temperature pyrolysis elemental analyzer (TC/EA, Thermo-Finnigan, Bremen, Germany). Feather δ²H values (δ²Hf) are reported as mean ± SD in delta notation of parts per mil (‰) from the standards (δDsample = [(Rsample/Rstandard) − 1]) compared to the VSMOW. We used chicken feather standard (CFS; -147.4‰), cow hooves (CHS; -187‰), and bowhead whale baleen (BWB; -108‰) as keratin that are widely adopted for sample comparative equilibration (Wassenaar and Hobson 2003). The details of hydrogen sample measurement are in Kelly et al. 2009. We conducted carbon and nitrogen stable isotope ratios analysis on a Thermo-Finnigan DeltaPlus isotope ratio mass spectrometer connected to a Carlo Erba elemental analyzer. We normalized the δ13C and δ15N values to the Vienna Pee Dee Belemnite (VPDB) and AIR scales, respectively, and used Brown-headed Cowbird (BHCO; Molothrus ater) feather standards. The values of δ13C and δ15N standards were -15.7 ± 0.1‰ (δ13C BHCO), and 7.6 ± 0.1‰ (δ15N BHCO).
Feather calibration and geographical assignments
We implemented a Bayesian analytical framework, in the R package assignR, to spatially reconstruct juncos’ breeding and/or molting areas (Ma et al. 2020, R Core Team 2020). First, we rescaled stable hydrogen isotope values in precipitation (δ²Hp) from a North American geostatistical model (Bowen et al. 2005) to 22 known-origin Song Sparrow (Melospiza melodia) samples provided by Hobson et al. (2012) and Hobson and Koehler (2015). To ensure a more accurate isotopic calibration (Wunder 2010), we used linear regression to model mean tissue isotope values as a function of δ²Hp using the calRaster function in assignR. We recognized that a species-specific calibration provides higher degree of accuracy. However, when species-specific known-origin tissues are not available, calibration data from closely related species can be used (Hobson et al. 2012). Our data set comprised entirely unknown-origin feather samples from Dark-eyed Juncos; therefore, we used published data of M. melodia, a species of comparable size, diet, and spatial distribution (Arcese et al. 2002, Hobson et al. 2012).
We determined the molting locations of juncos wintering in Oklahoma by computing posterior probability density maps through the pdRaster function in assignR (Wunder 2010, Ma et al. 2020). We presented normalized posterior probability maps, representing both the individual and cumulative group probabilities of origin, from each grid cell restricted to the breeding and year-round distribution ranges provided by BirdLife International 2016. To visualize the cumulative group probabilities of origin, we clustered the samples into four isotopic groups based on the reported stable isotope feather values (δ²Hf) across the junco data set arranged from the lowest values (G1) to the highest values (G4). The first group (G1) included δ²Hf values ranging from -175.6‰ to -160.2‰ (M = -167.1‰, SD = 5.02‰, N = 18); the second group (G2) included values ranging from -159.3‰ to -150.4‰ (M = -154.9‰, SD = 3.17‰, N = 23); the third group (G3) included values ranging from -149.8‰ to -140.1‰ (M = -145.4‰, SD = 2.88‰, N = 19); and the fourth group (G4) included values ranging from -139.3‰ to -118.6‰ (M = -130.4‰, SD = 6.39‰, N = 16). Even though this clustering approach is ad hoc, it still represents a useful subdivision based on δ²Hf variation across birds in our dataset. Wassenaar and Hobson (2006) estimated that ±3‰ variation in hydrogen stable isotope extracted from feathers translates approximately to one degree of latitude (or ~111 km). Therefore, the differential range of about 10‰ or more, which we used to cluster birds into four groups, corresponds to ~370 km of geographic separation (Hobson and Wassenaar 1996, Wassenaar and Hobson 2006). This distance is presumably large enough to consider our samples from different breeding populations.
Modeling morphometric parameters
We investigated the effect and relative predictive importance of five different morphometrics on mean hydrogen stable isotope ratios in Dark-eyed Junco feathers. Our explanatory variables were: wing chord (mm), tarsus length (mm), tail length (mm), mass (g), and fat depositions scored on a six-point ordinal scale (Helms and Drury 1960). We did not include body mass index, normalized by a structural measure, because this would be collinear with other included variables and additive model combinations should account for size normalized mass (Green 2001). We controlled for morphological variance across males and females by including “sex” as a random effect in all our models; and to standardize evaluation of predictive influence between the morphometric variables measured on different scales, we Z-scored values for all variables. We modeled a complete combinatorial set of linear models in the R package MuMIn (Bartón and Bartón 2019) and computed the relative importance of individual morphometric variables through a variable importance analysis using Akaike Information Criterion corrected for small sample sizes (AICc) and summed model weights for each model containing a given variable. We then compared the sum of all the AICc weights and odds ratios to evaluate the relative importance of each variable.
Trophic level analysis
We analyzed stable carbon (δ13C) and nitrogen (δ15N) isotopes in feather samples to explore trophic level partition among birds migrating from northern and southern latitudes as revealed by hydrogen (δ2H) data. We exploited the occurrence of distinct plant photosynthetic pathways, knowing that trees and shrubs in temperate climates (e.g., C3 plants) are depleted in carbon compared to plants inhabiting arid environments and adopting C4 or CAM photosynthesis (Keeley and Rundel 2003). These differences helped to assess the proportion of proteins derived from C3 and C4 plants, although higher nitrogen values indicated a shift toward a diet less dependent on seeds and richer in insects. We used the corr.test function, in the R package psych (Revelle and Revelle 2015) to perform a Pearson correlation test between carbon (δ13C) and hydrogen (δ2H), and between nitrogen (δ15N) and hydrogen (δ2H) stable isotopes. This test highlighted diet trends in relation to latitudinal origin as determined by δ2H data. We then used the levelplot function, in the R package lattice (Sarkar 2008), to create a visual representation of δ2H values from feathers collected in Oklahoma and their distribution within the sampling space.
RESULTS
Rescaling model and posterior probability density maps
The rescaling model provided a robust relationship between precipitation isotope values (δ²Hp) and known-origin samples of the species M. melodia (y = -35.08 + 0.91x; R² = 0.79, CI95% [0.74, 0.95], n = 22) and allowed us to calibrate our isoscape to determine junco’s posterior probability of origin (Fig. 1A). We obtained a rescaled environmental isoscape raster of 52,884 cells at 0.33 km resolution and a range of hydrogen stable isotope values spanning -193.3‰ and -41.3‰ (M = -122.9‰; SD = 37.2‰; Fig. 1B, C).
Once we performed isoscape tissue calibration, we inferred the spatial extent of the breeding and molting origin of 76 Dark-eyed Juncos wintering in Oklahoma through a Bayesian probabilistic framework. Our posterior probability density maps showed distinct latitudinal variation across the breeding and molting range of migratory juncos (Fig. 2). Overall, the northernmost migrants were assigned to either Alaska or western Canada, including the Yukon Territory and British Columbia, or central Northwest Territories, while southern migrants were assigned to either northern Saskatchewan or southern Manitoba and northwestern United States near the Rocky Mountains. However, even though the probability assignment surfaces for the southern migrants (group G3 and G4) were largely distributed at latitudes lower than 60°N (Fig. 2), we also found weak probabilities of origin along the western coast of Alaska because of the underlying structure of the precipitation isoscape used for rescaling (Fig. 1B, C). Collectively, these results suggest that different Dark-eyed Junco populations admix into the same wintering area in central Oklahoma.
Variable importance analysis
The analysis of the importance of variables revealed no clear predictive support for any variable considered in our models. We fit a full set of 32 linear mixed-effect models for mean δ²Hf including single variable models and all the possible combinations of the five predictors, with a random effect of sex, and ranked them using AICc. All five variables considered had relatively high summed AICc evidence weights suggesting models containing each respective variable outperformed those that did not (Table 1 and Table 2). Fat score had the highest summed AICc weight value and odd ratios, indicating that models containing the fat score variable accounted for all of the weight of AICc values for the model set (Table 1). However, the estimated effect of fat score on mean δ²Hf was indistinguishable from zero in all models weighted by AICc (Fig. 3A) suggesting that despite being the most comparatively informative variable, it had no discernible effect on δ²Hf values in our data set. Similarly, estimates for other variables we considered also lacked support for non-zero effects (Fig. 3B).
Migrant diet preferences and trophic level
The Pearson test did not identify a strong correlation between carbon (δ13C) and hydrogen (δ2H) isotope values (r = -0.17, p = 0.22), but it suggested a moderately positive correlation between nitrogen (δ15N) and hydrogen (δ2H) isotope values (r = 0.33, p = 0.01). These results suggest that a trophic level shift toward insect consumption might occur in migrants originating from lower latitudes (Fig. 4A, B). Interestingly, when samples were grouped by wing length (Fig. 4C, D), it appeared that birds with longer wings had a broader range of δ13C values, indicating variability in their habitat use. Moreover, when plotting δ13C against δ15N over δ2H feather isoscape within the sampling space, the changes in dietary niche across latitudinal origin appeared more evident (Fig. 5A-C; Appendix 1, Figs. A1, A2). Northern migrants showed lower protein consumption (δ15N: 1-4‰) and C3-shrub habitat use (δ13C: -22 − -24‰), while southern migrants were characterized by a higher trophic level (δ15N: 4-8‰) and δ13C toward C3-vegetation compatible with temperate forests (δ13C: -24 − -26‰). Analysis of diet preferences between sexes did not show marked differences, although males seem to have a slightly larger dietary niche compared to females.
DISCUSSION
Bridge et al. (2010) conducted a captive experiment in which wintering Dark-eyed Juncos were fed ad libitum to demonstrate that fat depositions allow them to initiate migration earlier in the spring compared to the control group. We extended previous work by providing a geostatistical approach illustrating the explicit spatial distribution of breeding and molting origin of fall migrants and related that distribution to morphological characteristics and trophic level differentiation. Our geographical assignments clearly suggested that migratory juncos wintering in central Oklahoma originated from different breeding and molting areas (Fig. 2). We identified northern and southern migrants, although each group also showed intracohort spatial variation ranging from Alaska and western Canada to central Northwest Territories, and northern Saskatchewan to southern Manitoba and northwestern United States, respectively. However, despite the posterior probability surfaces showing population admixture at the wintering area in Oklahoma, our modeling effort showed no association between winter body condition and breeding or molting latitude.
Stable isotope ratios of hydrogen, carbon, and nitrogen, extracted from inert animal tissues, such as claws and feathers, reflect the isotope ratios in the environment in which those tissues were grown and can provide inferences about large-scale animal movements and diet preferences in species with small body size (Rubenstein and Hobson 2004, Vander Zanden et al. 2018, Zenzal et al. 2018). However, a clear advantage of adopting a multielement stable isotope approach is that migrants can be assigned to different breeding and molting grounds of origin while their diet preferences are also revealed and clustered in trophic guilds (Herrera et al. 2003). The positive correlation between nitrogen (δ15N) and hydrogen (δ2H) values, although moderate, corroborated our hypothesis that wintering birds originating from northern latitudes have lower protein intake relative to southern migrants and prefer a diet rich in carbohydrates before migrating. That is, increased carbohydrate anabolism and lower protein consumption is necessary to quickly synthesize enough subcutaneous lipid reserves to sustain a long-distance migration (Cecere et al. 2011, Domer et al. 2018). Alternatively, higher δ15N values in southern migrants might be explained by environmental stressors. An early study in captive quails (Coturnix japonica) and geese (Anser rossii) showed that increased δ15N values can be obtained from growing tissues of individuals maintained under severe nutritional stress conditions (e.g., fasting) compared to control groups (Hobson et al. 1993). However, subsequent analysis in sparrows (Melospiza melodia) found no differences in δ15N between birds kept under a restricted diet versus birds fed ad libitum (Kempster et al. 2007) suggesting a less straightforward relationship between physiological stress conditions and δ15N values. Therefore, even though we cannot pinpoint the exact causes of increased δ15N values in growing feathers (e.g., baseline difference) and that other species have shown variation in feather δ15N values between boreal and more southern food webs based on land use (Hobson 1999), it is also possible that migratory juncos originating from southern latitudes were foraging at a higher level in the trophic system and consuming more insects compared to northern migrants. We do not exclude that such diet preference might be linked to an opportunistic foraging strategy due to an outburst of insect availability (Yang et al. 2008). However, analysis of isotopic niche shifts in avian migratory populations have shown sharp changes during different life cycle stages (Hahn et al. 2013), confirming that variations in diet compositions have a defined role in maintaining fluctuating energy needs (Marshall et al. 2016). Further, our δ13C results indicated that northern juncos might exploit food resources from a broader range of habitats compared to southern juncos, ranging from pine forest to deciduous forest (e.g., oak) and C3 shrubs. Bearhop et al. (2004) studied the Thick-billed Vireos (Vireo crassirostris) in the Bahamas, and based on mean claw δ13C values, found differences among two groups of birds occupying environments with either prevalent xerophytic shrubs or pine forest. Although our feather δ13C results showed that northern juncos’ seed consumption fell within the spectrum of plants characterized by a C3 photosynthetic pathway, and we found enough variation to highlight habitat occupancy differentiations across latitudinal origins (Figs. 4, 5), we note that a robust environmental isotopic baseline is necessary to determine dietary and habitat preferences with confidence (Post 2002).
The interactions between avian body mass fluctuations and migratory behaviors within and between seasons remain poorly understood (Brown and Sherry 2006, Tsvey et al. 2007, Marra et al. 2015). Importantly, body condition appears to both influence and be influenced by migratory behavior. For example, body condition may affect migratory decisions (Schaub and Jenni 2000); several species of migrating shorebirds underwent longer migratory flights when departing with higher body condition scores (Anderson et al. 2019). Similarly, spring arrival timing of Red Knots (Calidris canutus) was associated with better body condition at a major stopover site (Duijns et al. 2017) and conditions experienced during winter months by American Redstarts (Setophaga ruticilla) led to variation in body conditions that predicted spring departure dates (Marra et al. 1998). In our investigation, individuals that migrated minimum distances did not exhibit larger body masses or higher fat scores over the early winter period. However, our study was limited because individual migrants were not captured immediately upon arrival in Oklahoma. Although challenging, an earlier capturing effort might reveal mass as a predictor of latitude of origin because the body condition of those migrants would provide a stronger indication of the migration strategy performed. Moreover, there may be other mechanisms that could explain the absence of a relationship between body conditions and migration strategies in our study population. For example, the taxonomy of the genus Junco has been challenging for decades and is far from being resolved (Miller 1941, Nolan et al. 2002, Aleixandre et al. 2013). It has recently been proposed that the species Junco hyemalis be phylogenetically reassessed, based on extensive genomic data, to determine if it needs to be split into four new species that also account for distinct phenotypic variation (Friis et al. 2016). Thus, in a complex migratory system, it is possible that different individuals from populations characterized by a larger or smaller than average body size may come into contact at the winter ground. This admixture of individuals could represent a challenge when assessing ther energy resources stored and used during migration. Therefore, meaningful comparisons between populations must account for body size differences relative to subcutaneous fat depositions. Alternatively, long distance migrants could also improve their body conditions (e.g., increase mass at stopover sites) before arriving at their final wintering destination and reduce the difference in body mass compared to short distance migrants. However, if the stopover site is only used to refuel without growing new feathers, the stable isotope analysis will still detect the northern origin (e.g., molting ground) and correctly identify the migration latitudinal origin without revealing any geographical information on the stopover location. By contrast, if blood samples are collected soon after arrival, then a comparative stable isotope analysis between feathers and blood samples might reveal the use of stopover areas and potential shifts in diet during migration.
CONCLUSIONS
By implementing a Bayesian assignment framework, we offered a probabilistic geographic explanation of the juncos’ feather isotopic variation across the species’ breeding and molting areas in the northern hemisphere and attempted to explain latitudinal origin in relation to morphological traits (Wunder 2010, Alerstam 2011, Maggini et al. 2016). In our system, none of the variables considered had a clear effect on feather hydrogen stable isotope values (used as a proxy for latitudinal origin), although we found indication of a possible higher trophic level shift in individuals migrating from southern latitudes. These findings promote the concurrent use of different stable isotope markers as a means of studying interconnected behavioral traits. Our results also highlight the need to carefully examine confounding effects in explanatory models in relation to the species’ natural history and possible differential migratory behaviors across populations.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
AC conceptualized the project, analyzed the data, and wrote the manuscript. AKP analyzed the data, reviewed, and edited the manuscript. SWY provided key feedback on data analysis, reviewed, and edited the manuscript. ESB collected the data and reviewed the manuscript. JFK collected the data and reviewed the manuscript. MBW supervised data analysis and reviewed the manuscript.
ACKNOWLEDGMENTS
We thank the reviewers and Vojtech Brlík for their constructive comments, which helped us to improve the first draft of the manuscript.
DATA AVAILABILITY
Data and code from this study will be deposited in Zenodo repository upon acceptance.
LITERATURE CITED
Aleixandre, P., J. H. Montoya, and B. Mila. 2013. Speciation on oceanic islands: rapid adaptive divergence vs. cryptic speciation in a Guadalupe Island songbird (Aves: Junco). PLoS One 8(5):e63242. https://doi.org/10.1371/journal.pone.0063242
Alerstam, T. 2011. Optimal bird migration revisited. Journal of Ornithology 152(1):5-23. https://doi.org/10.1007/s10336-011-0694-1
Alerstam, T., and J. Bäckman. 2018. Ecology of animal migration. Current Biology 28(17):R968-R972. https://doi.org/10.1016/j.cub.2018.04.043
Anderson, A. M., S. Duijns, P. A. Smith, C. Friis, and E. Nol. 2019. Migration istance and body condition influence shorebird migration strategies and stopover decisions during southbound migration. Frontiers in Ecology and Evolution 7:251. https://doi.org/10.3389/fevo.2019.00251
Arcese, P., M. K. Sogge, A. B. Marr, and M. A. Patten. 2002. Song Sparrow (Melospiza melodia). Version 2.0. In P. G. Rodewald, editor. The birds of North America. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/tbna.704.p
Barta, Z., J. M. McNamara, A. I. Houston, T. P. Weber, A. Hedenström, and O. Fero. 2008. Optimal moult strategies in migratory birds. Philosophical Transactions of the Royal Society B: Biological Sciences 363(1490):211-229. https://doi.org/10.1098/rstb.2007.2136
Bartón, K., and M. K. Bartón. 2019. Package ‘MuMIn’. Multi-model inference. Version 1(6). R Foundation for Statistical Computing, Vienna, Austria. https://CRAN.R-project.org/package=MuMIn
Bearhop, S., G. M. Hilton, S. C. Votier, and S. Waldron. 2004. Stable isotope ratios indicate that body condition in migrating passerines is influenced by winter habitat. Proceedings of the Royal Society B: Biological Sciences 271(suppl_4):S215-S218. https://doi.org/10.1098/rsbl.2003.0129
Besozzi, E., B. Chew, D. C. Allen, and A. Contina. 2021. Stable isotope analysis of an aberrant Painted Bunting (Passerina ciris) feather suggests post-molt movements. Wilson Journal of Ornithology 133(1):151-158. https://doi.org/10.1676/20-00140
Bird Conservancy of the Rockies. 2017. Avian conservation assessment database (Partners in Flight). Bird Conservancy of the Rockies, Brighton, Oregon, USA. http://pif.birdconservancy.org
BirdLife International. 2016. Handbook of the birds of the world. Bird species distribution maps of the world. Version 6.0. Cambridge, England, U.K. http://datazone.birdlife.org/species/requestdis.
Bowen, G. J., L. I. Wassenaar, and K. A. Hobson. 2005. Global application of stable hydrogen and oxygen isotopes to wildlife forensics. Oecologia 143(3):337-348. https://doi.org/10.1007/s00442-004-1813-y
Bowlin, M. S., and M. Wikelski. 2008. Pointed wings, low wingloading and calm air reduce migratory flight costs in songbirds. PLoS One 3(5):e2154. https://doi.org/10.1371/journal.pone.0002154
Bridge, E. S., J. F. Kelly, P. E. Bjornen, C. M. Curry, P. H. Crawford, and J. M. Paritte. 2010. Effects of nutritional condition on spring migration: do migrants use resource availability to keep pace with a changing world? Journal of Experimental Biology 213(14):2424-2429. https://doi.org/10.1242/jeb.041277
Brown, D. R., and T. W. Sherry. 2006. Food supply controls the body condition of a migrant bird wintering in the tropics. Oecologia 149(1):22-32. https://doi.org/10.1007/s00442-006-0418-z
Cecere, J. G., F. Spina, S. Jenni-Eiermann, and L. Boitani. 2011. Nectar: an energy drink used by European songbirds during spring migration. Journal of Ornithology 152(4):923-931. https://doi.org/10.1007/s10336-011-0675-4
Chew, B., J. F. Kelly, A. Contina. 2019. Stable isotopes in avian research: a step by step protocol to feather sample preparation for stable isotope analysis of carbon (δ13C), nitrogen (δ15N), and hydrogen (δ2H). Version 1.1. Protocols.io. https://dx.doi.org/10.17504/protocols.io.z2uf8ew
DeLuca, W. V., B. K. Woodworth, C. C. Rimmer, P. P. Marra, P. D. Taylor, K. P. McFarland, S. A. Mackenzie, and D. R. Norris. 2015. Transoceanic migration by a 12 g songbird. Biology Letters 11(4):20141045. https://doi.org/10.1098/rsbl.2014.1045
Domer, A., O. Ovadia, and E. Shochat. 2018. Energy for the road: influence of carbohydrate and water availability on fueling processes in autumn-migrating passerines. Auk 135(3):534-546. https://doi.org/10.1642/AUK-17-228.1
Duijns, S., L. J. Niles, A. Dey, Y. Aubry, C. Friis, S. Koch, A. M. Anderson, and P. A. Smith. 2017. Body condition explains migratory performance of a long-distance migrant. Proceedings of the Royal Society B: Biological Sciences 284(1866):20171374. https://doi.org/10.1098/rspb.2017.1374
Friis, G., P. Aleixandre, R. Rodríguez‐Estrella, A. G. Navarro‐Sigüenza, and B. Milá. 2016. Rapid postglacial diversification and long‐term stasis within the songbird genus Junco: phylogeographic and phylogenomic evidence. Molecular Ecology 25(24):6175-6195. https://doi.org/10.1111/mec.13911
Fudickar, A. M., T. J. Greives, J. W. Atwell, C. A. Stricker, and E. D. Ketterson. 2016. Reproductive allochrony in seasonally sympatric populations maintained by differential response to photoperiod: implications for population divergence and response to climate change. American Naturalist 187(4):436-446. https://doi.org/10.1086/685296
Green, A. J. 2001. Mass/length residuals: measures of body condition or generators of spurious results? Ecology 82(5):1473-1483. https://doi.org/10.1890/0012-9658(2001)082[1473:MLRMOB]2.0.CO;2
Guglielmo, C. G. 2018. Obese super athletes: fat-fueled migration in birds and bats. Journal of Experimental Biology 221(Suppl_1):jeb165753. https://doi.org/10.1242/jeb.165753
Hahn, S., V. Amrhein, P. Zehtindijev, and F. Liechti. 2013. Strong migratory connectivity and seasonally shifting isotopic niches in geographically separated populations of a long-distance migrating songbird. Oecologia 173(4):1217-1225. https://doi.org/10.1007/s00442-013-2726-4
Helms, C. W., and W. H. Drury. 1960. Winter and migratory weight and fat field studies on some North American buntings. Bird-banding 31(1):1-40. https://doi.org/10.2307/4510793
Herrera, L. G., K. A. Hobson, M. Rodríguez, and P. Hernandez. 2003. Trophic partitioning in tropical rain forest birds: insights from stable isotope analysis. Oecologia 136(3):439-444. https://doi.org/10.1007/s00442-003-1293-5
Hobson, K. A. 1999. Stable-carbon and nitrogen isotope ratios of songbird feathers grown in two terrestrial biomes: implications for evaluating trophic relationships and breeding origins. Condor 101(4):799-805. https://doi.org/10.2307/1370067
Hobson, K. A. 2005. Using stable isotopes to trace long-distance dispersal in birds and other taxa. Diversity and Distributions 11(2):157-164. https://doi.org/10.1111/j.1366-9516.2005.00149.x
Hobson, K. A., R. T. Alisauskas, and R. G. Clark. 1993. Stable-nitrogen isotope enrichment in avian tissues due to fasting and nutritional stress: implications for isotopic analyses of diet. Condor 95(2):388-394. https://doi.org/10.2307/1369361
Hobson, K. A., and G. Koehler. 2015. On the use of stable oxygen isotope (δ18O) measurements for tracking avian movements in North America. Ecology and Evolution 5(3):799-806. https://doi.org/10.1002/ece3.1383
Hobson, K. A., S. L. Van Wilgenburg, L. I. Wassenaar, and K. Larson. 2012. Linking hydrogen (δ2H) isotopes in feathers and precipitation: sources of variance and consequences for assignment to isoscapes. PLoS One 7(4):e35137. https://doi.org/10.1371/journal.pone.0035137
Hobson, K. A., and L. I. Wassenaar. 1996. Linking breeding and wintering grounds of neotropical migrant songbirds using stable hydrogen isotopic analysis of feathers. Oecologia 109(1):142-148. https://doi.org/10.1007/s004420050068
Horton, K. G., B. M. Van Doren, P. M. Stepanian, A. Farnsworth, and J. F. Kelly. 2016. Seasonal differences in landbird migration strategies. Auk 133(4):761-769. https://doi.org/10.1642/AUK-16-105.1
Kaboli, M., M. Aliabadian, A. Guillaumet, C. S. Roselaar, and R. Prodon. 2007. Ecomorphology of the wheatears (genus Oenanthe). Ibis 149(4):792-805. https://doi.org/10.1111/j.1474-919X.2007.00714.x
Keeley, J. E., and P. W. Rundel. 2003. Evolution of CAM and C4 carbon-concentrating mechanisms. International Journal of Plant Sciences 164(S3):S55-S77. https://doi.org/10.1086/374192
Kelly, J. F., E. S. Bridge, A. M. Fudickar, and L. I. Wassenaar. 2009. A test of comparative equilibration for determining non-exchangeable stable hydrogen isotope values in complex organic materials. Rapid Communications in Mass Spectrometry 23(15):2316-2320. https://doi.org/10.1002/rcm.4150
Kempster, B., L. Zanette, F. J. Longstaffe, S. A. MacDougall-Shackleton, J. C. Wingfield, and M. Clinchy. 2007. Do stable isotopes reflect nutritional stress? Results from a laboratory experiment on song sparrows. Oecologia 151(3):365-371. https://doi.org/10.1007/s00442-006-0597-7
Liebgold, E. B., N. M. Gerlach, and E. D. Ketterson. 2019. Density-dependent fitness, not dispersal movements, drives temporal variation in spatial genetic structure in Dark-eyed Juncos (Junco hyemalis). Molecular Ecology 28(5):968-979. https://doi.org/10.1111/mec.15040
Ma, C., H. B. Vander Zanden, M. B. Wunder, and G. J. Bowen. 2020. assignR: an R package for isotope-based geographic assignment. Methods in Ecology and Evolution 11(8):996-1001. https://doi.org/10.1111/2041-210X.13426
Maggini, I., B. Metzger, M. Voss, C. C. Voigt, and F. Bairlein. 2016. Morphometrics and stable isotopes differentiate wintering populations of a migratory bird. Movement Ecology 4:20. https://doi.org/10.1186/s40462-016-0085-6
Marra, P. P., E. B. Cohen, S. R. Loss, J. E. Rutter, and C. M. Tonra. 2015. A call for full annual cycle research in animal ecology. Biology Letters 11(8):20150552. https://doi.org/10.1098/rsbl.2015.0552
Marra, P. P., K. A. Hobson, and R. T. Holmes. 1998. Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science 282(5395):1884-1886. https://doi.org/10.1126/science.282.5395.1884
Marshall, T. J., M. F. Dick, and C. G. Guglielmo. 2016. Seasonal dietary shifting in Yellow-rumped Warblers is unrelated to macronutrient targets. Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology 192:57-63. https://doi.org/10.1016/j.cbpa.2015.11.014
Miller, A. H. 1941. Speciation in the avian genus Junco. University of California Publications in Zoology. Biology 44:173–434. https://doi.org/10.2307/4509722
Nolan, V., Jr., and E. D. Ketterson. 1983. An analysis of body mass, wing length, and visible fat deposits of Dark-eyed Juncos wintering at different latitudes. Wilson Bulletin 95(4):603-620. https://www.semanticscholar.org/paper/AN-ANALYSIS-OF-BODY-MASS%2C-WING-LENGTH%2C-AND-VISIBLE-Ketterson/494491e68d80af9d16ac37279f2cf60a0f8118e8
Nolan, V., Jr., E. D. Ketterson, D. A. Cristol, C. M. Rogers, E. D. Clotfelter, R. C. Titus, S. J. Schoech, and E. Snajdr. 2002. Dark-eyed Junco (Junco hyemalis). Version 2.0. In A. F. Poole and F. B. Gill, editors. The birds of North America. Cornell Lab of Ornithology, Ithaca, New York, USA. https://doi.org/10.2173/bna.716
Paritte, J. M., and J. F. Kelly. 2009. Effect of cleaning regime on stable-isotope ratios of feathers in Japanese Quail (Coturnix japonica). Auk 126(1):165-174. https://doi.org/10.1525/auk.2009.07187
Post, D. M., 2002. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83(3):703-718. https://doi.org/10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2
R Core Team. 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.r-project.org.
Rani, S., S. Singh, S. Malik, and V. Kumar. 2017. Insights into the regulation of spring migration in songbirds. Pages 625-642 in V. Kumar, editor. Biological timekeeping: clocks, rhythms and behaviour. Springer, New Delhi, Delhi, India. https://doi.org/10.1007/978-81-322-3688-7_30
Reif, J., D. Hořák, A. Krištín, L. Kopsová, and V. Devictor. 2016. Linking habitat specialization with species’ traits in European birds. Oikos 125(3):405-413. https://doi.org/10.1111/oik.02276
Revelle, W., and M. W. Revelle. 2015. Package ‘psych’. The comprehensive R archive network. R Foundation for Statistical Computing, Vienna, Austria. https://cran.r-project.org/web/packages/psych/psych
Rockwell, S. M., J. D. Alexander, J. L. Stephens, R. I. Frey, and C. J. Ralph. 2017. Spatial variation in songbird demographic trends from a regional network of banding stations in the Pacific Northwest. Condor 119(4):732-744. https://doi.org/10.1650/CONDOR-17-44.1
Rogers, C. M., V. Nolan, and E. D. Ketterson. 1994. Winter fattening in the Dark-eyed Junco: plasticity and possible interaction with migration trade-offs. Oecologia 97(4):526-532. https://doi.org/10.1007/BF00325892
Rubenstein, D. R., and K. A. Hobson. 2004. From birds to butterflies: animal movement patterns and stable isotopes. Trends in Ecology and Evolution 19(5):256-263. https://doi.org/10.1016/j.tree.2004.03.017
Rundel, C. W., M. B. Wunder, A. H. Alvarado, K. C. Ruegg, R. Harrigan, A Schuh, J. F. Kelly, R. B. Siegel, D. F. DeSante, T. B. Smith, and J. Novembre. 2013. Novel statistical methods for integrating genetic and stable isotope data to infer individual-level migratory connectivity. Molecular Ecology 22(16):4163-4176. https://doi.org/10.1111/mec.12393
Sarkar, D. 2008. The lattice package: lattice graphics. R package, version 0.17-15. R Foundation for Statistical Computing, Vienna, Austria. https://CRAN.R-project.org/package=lattice
Schaub, M., and L. Jenni. 2000. Body mass of six long-distance migrant passerine species along the autumn migration route. Journal Für Ornithologie 141:441-460. https://doi.org/10.1007/bf01651574
Tsvey, A., V. N. Bulyuk, and V. Kosarev. 2007. Influence of body condition and weather on departures of first-year European Robins, Erithacus rubecula, from an autumn migratory stopover site. Behavioral Ecology and Sociobiology 61(11):1665-1674. https://doi.org/10.1007/s00265-007-0397-z
Vander Zanden, H. B., D. M. Nelson, M. B. Wunder, T. J. Conkling, and T. Katzner. 2018. Application of isoscapes to determine geographic origin of terrestrial wildlife for conservation and management. Biological Conservation 228:268-280. https://doi.org/10.1016/j.biocon.2018.10.019
Wassenaar, L. I., and K. A. Hobson. 2003. Comparative equilibration and online technique for determination of non-exchangeable hydrogen of keratins for use in animal migration studies. Isotopes in Environmental and Health Studies 39(3):211-217. https://doi.org/10.1080/1025601031000096781
Wassenaar, L. I., and K. A. Hobson. 2006. Stable-hydrogen isotope heterogeneity in keratinous materials: mass spectrometry and migratory wildlife tissue subsampling strategies. Rapid Communications in Mass Spectrometry 20(16):2505-2510. https://doi.org/10.1002/rcm.2626
Wunder, M. B. 2010. Using isoscapes to model probability surfaces for determining geographic origins. Pages 251-270 in J. West, G. Bowen, T. Dawson, and K. Tu, editors. Isoscapes. Springer, Dordrecht, The Netherlands. https://doi.org/10.1007/978-90-481-3354-3_12
Yanco, S. W., A. McDevitt, C. N. Trueman, L. Hartley, and M. B. Wunder. 2020. A modern method of multiple working hypotheses to improve inference in ecology. Royal Society Open Science 7(6):200231. https://doi.org/10.1098/rsos.200231
Yang, L. H., J. L. Bastow, K. O. Spence, and A. N. Wright. 2008. What can we learn from resource pulses. Ecology 89(3):621-634. https://doi.org/10.1890/07-0175.1
Zenzal, T. J., A. Contina, J. F. Kelly, and F. R. Moore. 2018. Temporal migration patterns between natal locations of Ruby-throated Hummingbirds (Archilochus colubris) and their Gulf Coast stopover site. Movement Ecology 6(1):1-7. https://doi.org/10.1186/s40462-017-0120-2

Fig. 1
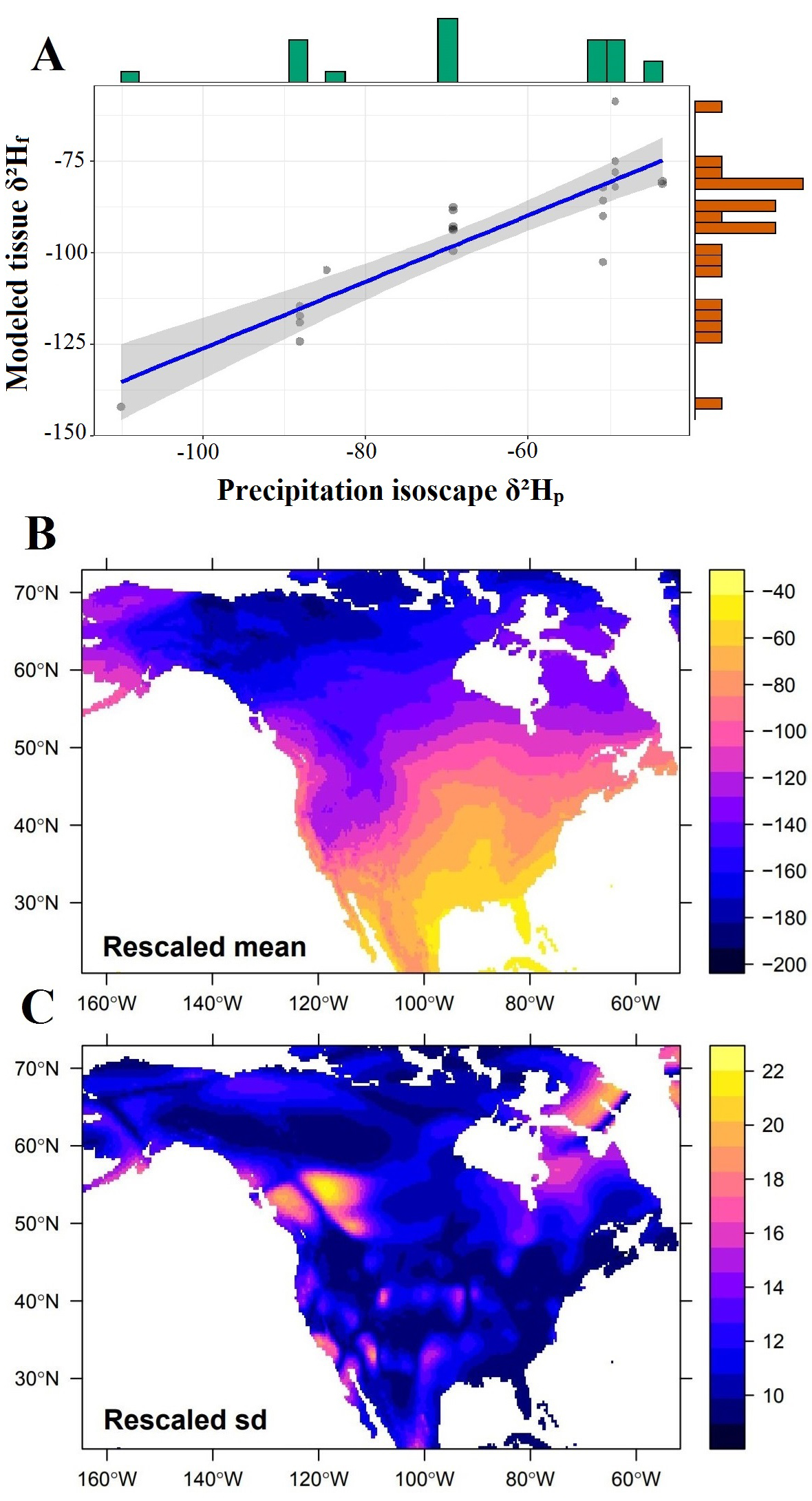
Fig. 1. Linear regression model between environmental precipitation isotope values (δ²Hp) and 22 known-origin feather samples of Melospiza melodia samples (A) used to rescale the North America hydrogen isoscape (B and C) for assigning migratory juncos wintering in Oklahoma to their molting locations at the breeding grounds across northern United States and Canada. The green and orange bars along the horizontal and vertical axes represent the distributions of the sampling data across environmental and tissue isotope values, respectively (A). The calibrated hydrogen isoscape mean values and standard deviations are presented in B and C, respectively.
Fig. 2
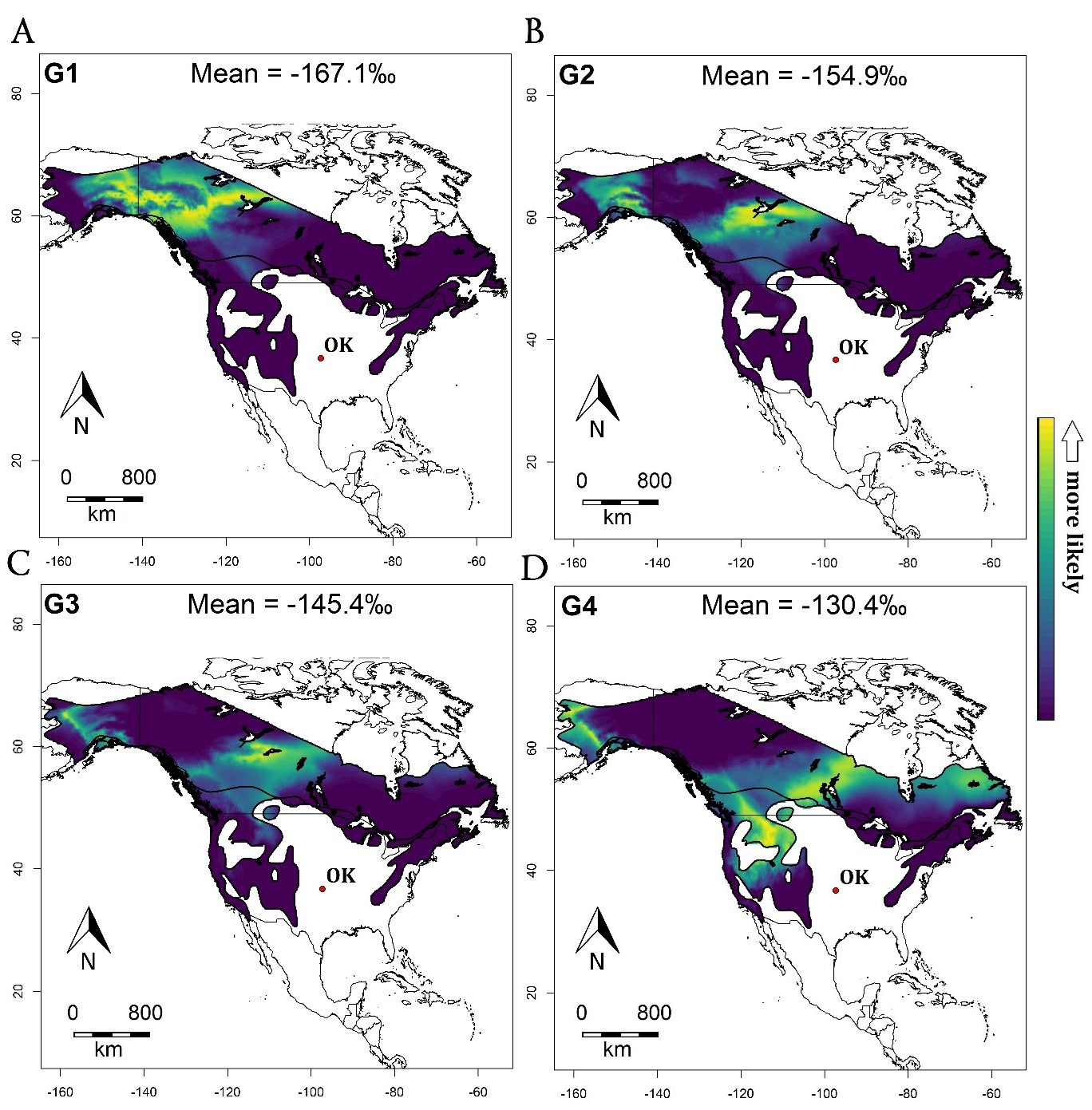
Fig. 2. Cumulative assignment posterior probability density maps across 4 isotopic groups of 76 Dark-eyed Juncos wintering in Oklahoma (red dot) and breeding at northern latitudes in the United States and Canada. The bright yellow color represents high probabilities of molting origin. The northern migrants (groups G1 and G2; A and B) were assigned to either Alaska or western Canada, including Yukon and British Columbia, or central Northwest Territories. The southern migrants (groups G3 and G4; C and D) were prevalently assigned to either northern Saskatchewan or southern Manitoba and northern western United States. The hydrogen stable isotope approach suggests that Dark-eyed Junco (Junco hyemalis) migrants from different breeding populations converge into the same location in Oklahoma over the winter.
Fig. 3
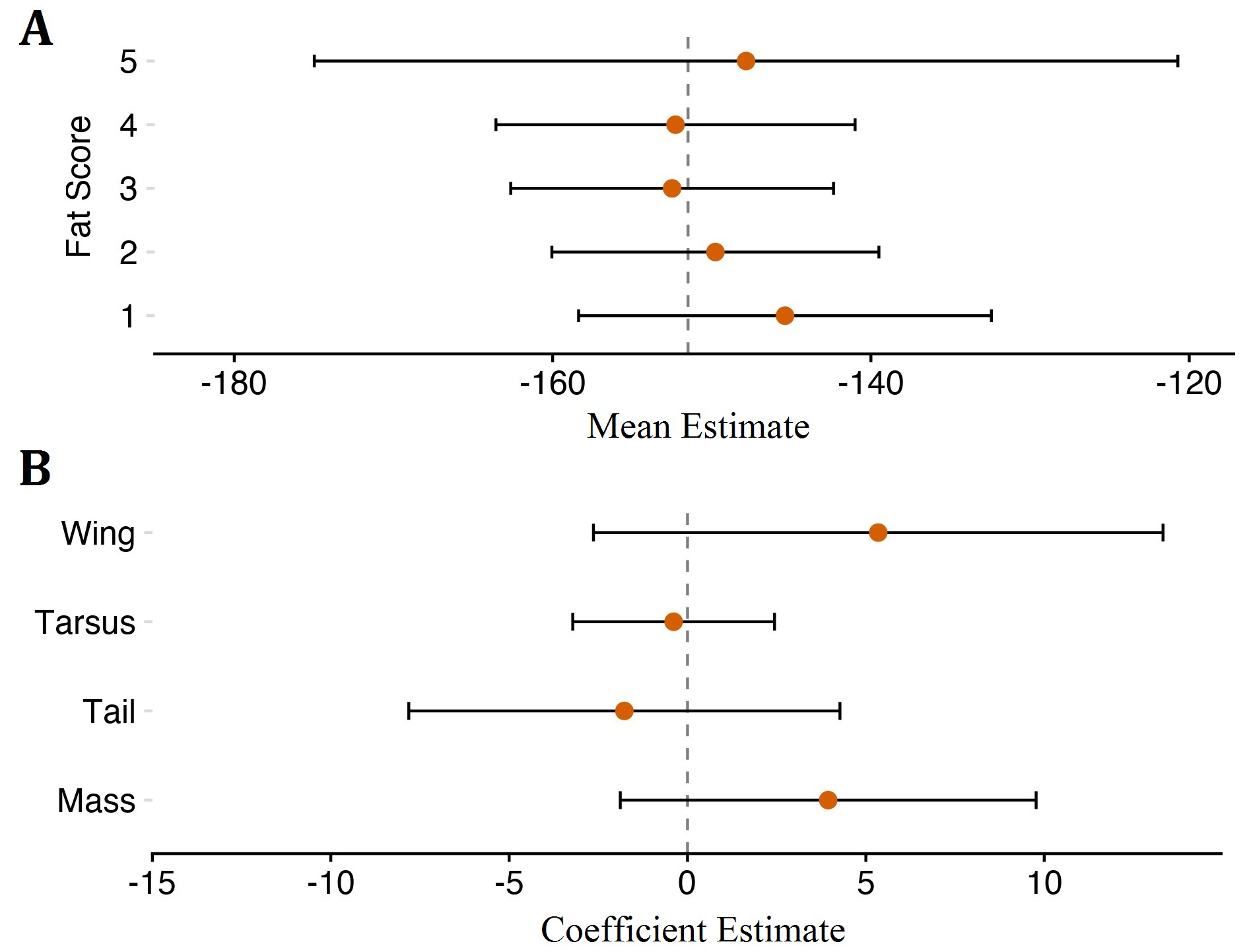
Fig. 3. Model averaged estimates of mean δ²Hf values with 95% confidence interval (CI) for levels of fat scores with centered and scaled morphometric metric measures held at 0; their mean value (A) and model averaged coefficient estimates of centered and scaled morphometric parameters with 95% CI with level 1 of fat score as the reference for the intercept (B). Both plots indicate effects for fat scores and morphometric variables on mean δ²Hf are insignificant for our data set as evidenced by CI overlap between fat score levels (A) and zero (B).
Fig. 4
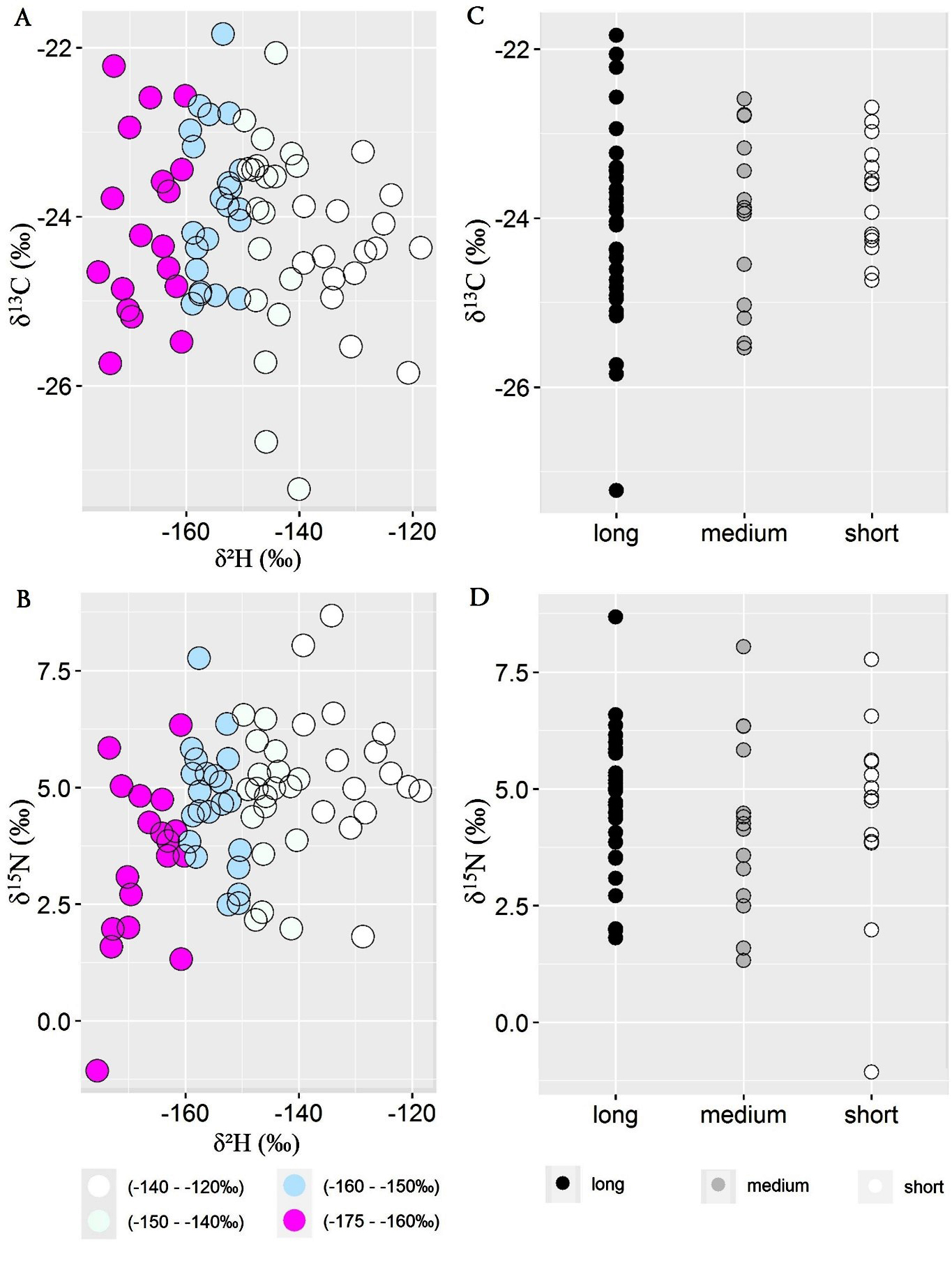
Fig. 4. Correlation between δ13C and hydrogen δ2H (A) and nitrogen δ15N and hydrogen δ2H values (B). Migrants exhibiting longer wings show a broader range of δ13C values compared to medium- and short-length winged birds suggesting more flexibility in their habitat use (C). We did not find noticeable trends between δ15N and wing length (D).
Fig. 5
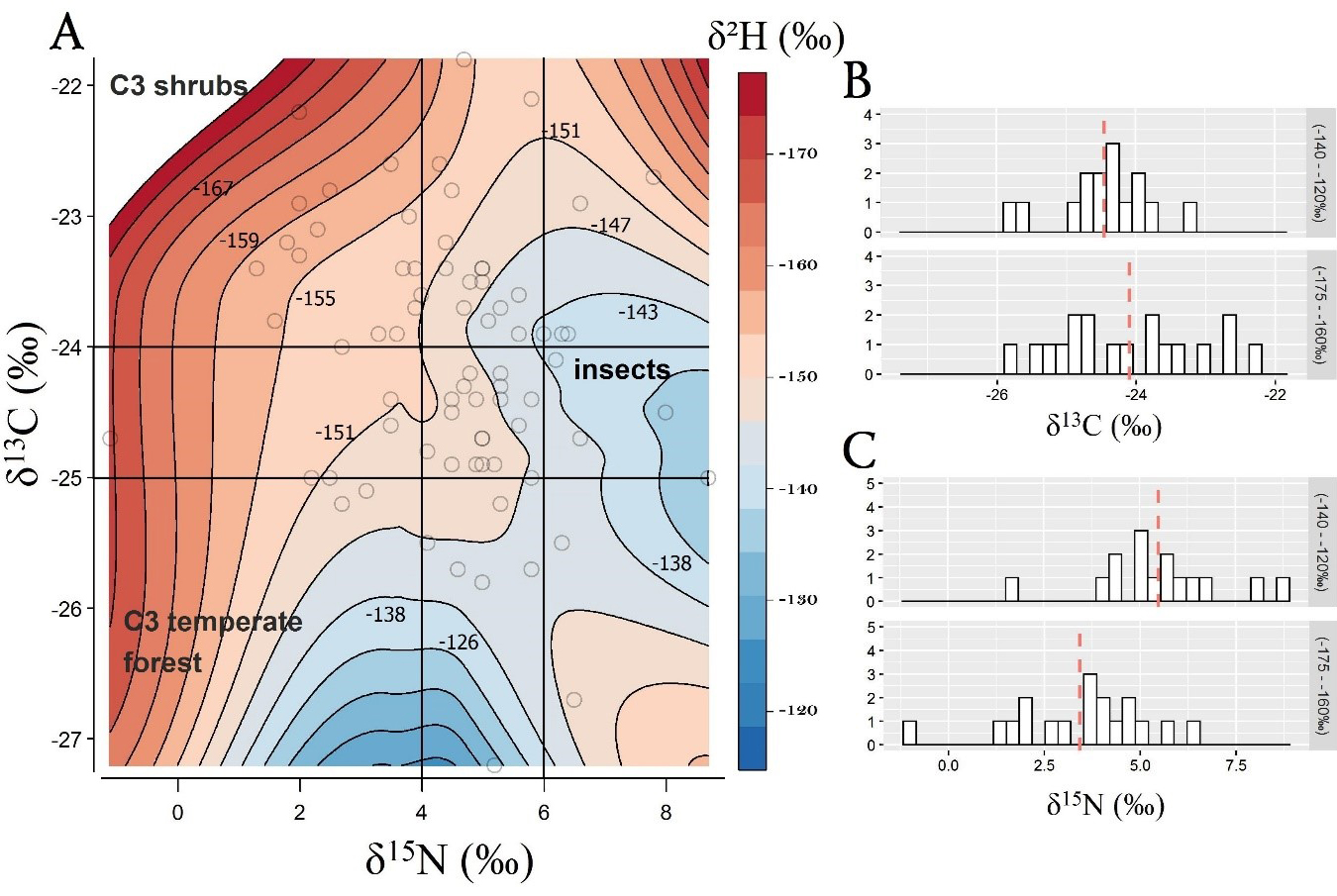
Fig. 5. Isotopic diet composition of δ13C and δ15N plotted over δ2H spatial data distribution (A). Blue areas represent lower latitudes of origin (δ2H = from -110‰ to -145‰) while red areas represent higher latitudes of origin (δ2H = from -145‰ to -180‰). Each open circle represents a single individual. Northern migrants show a moderate trend toward lower protein consumption (δ15N: 1-4‰) and C3-plant habitat use (δ13C: -22 − -24‰). Southern migrants show higher trophic level (δ15N: 4-8‰) and temperate forest habitat use (δ13C: -24 − -26‰). B and C show the distributions of δ13C and δ15N values across migrants originating from extreme northern and extreme southern latitudes. Dashed red lines represent isotope mean values for each group.
Table 1
Table 1. Variable importance results. Although the variable body fat emerged as the comparatively best predictor of hydrogen variation in feathers, the effect for this and all other variables could not be differentiated from zero. Note: AICc = Akaike Information Criterion corrected for small sample sizes; inf = infinite.
| Variable | Sum of AICc model weights |
Odds ratio |
| Fat | 1 | inf |
| Mass | 0.87 | 6.69 |
| Wing | 0.86 | 6.14 |
| Tail | 0.72 | 2.57 |
| Tarsus | 0.54 | 1.17 |
Table 2
Table 2. Akaike Information Criterion corrected for small sample size (AICc) model selection results. The most parsimonious model was the full model including all five morphological variables but was equally competitive with a model omitting tarsus based on < 2 unit difference in ΔAICc values. Simpler single term or intercept only models were generally outperformed by more complex models.
| Model ID | Model variables | LogLik | AICc | Delta | Weight |
| 32 | h ~ Fat + Mass + Tail + Tarsus + Wing + (1|sex) | -222.1 | 471.48 | 0 | 0.3 |
| 24 | h ~ Fat + Mass + Tail + Wing + (1|sex) | -223.74 | 471.8 | 0.33 | 0.25 |
| 28 | h ~ Fat + Mass + Tarsus + Wing + (1|sex) | -224.61 | 473.54 | 2.06 | 0.11 |
| 20 | h ~ Fat + Mass + Wing + (1|sex) | -226.25 | 473.95 | 2.47 | 0.09 |
| 30 | h ~ Fat + Tail + Tarsus + Wing + (1|sex) | -225.43 | 475.16 | 3.69 | 0.05 |
| 22 | h ~ Fat + Tail + Wing + (1|sex) | -226.94 | 475.33 | 3.86 | 0.04 |
| 16 | h ~ Fat + Mass + Tail + Tarsus + (1|sex) | -225.65 | 475.61 | 4.13 | 0.04 |
| 8 | h ~ Fat + Mass + Tail + (1|sex) | -227.18 | 475.83 | 4.35 | 0.03 |
| 12 | h ~ Fat + Mass + Tarsus + (1|sex) | -227.46 | 476.38 | 4.91 | 0.03 |
| 4 | h ~ Fat + Mass + (1|sex) | -228.98 | 476.68 | 5.21 | 0.02 |
| 26 | h ~ Fat + Tarsus + Wing + (1|sex) | -227.69 | 476.85 | 5.37 | 0.02 |
| 18 | h ~ Fat + Wing + (1|sex) | -229.2 | 477.11 | 5.63 | 0.02 |
| 14 | h ~ Fat + Tail + Tarsus + (1|sex) | -229.25 | 479.96 | 8.48 | 0 |
| 6 | h ~ Fat + Tail + (1|sex) | -230.84 | 480.39 | 8.91 | 0 |
| 10 | h ~ Fat + Tarsus + (1|sex) | -232.01 | 482.74 | 11.26 | 0 |
| 31 | h ~ Mass + Tail + Tarsus + Wing + (1|sex) | -233.91 | 483.9 | 12.42 | 0 |
| 2 | h ~ Fat + (1|sex) | -233.92 | 483.91 | 12.44 | 0 |
| 23 | h ~ Mass + Tail + Wing + (1|sex) | -235.4 | 484.32 | 12.85 | 0 |
| 27 | h ~ Mass + Tarsus + Wing + (1|sex) | -236.3 | 486.13 | 14.66 | 0 |
| 19 | h ~ Mass + Wing + (1|sex) | -237.79 | 486.64 | 15.16 | 0 |
| 15 | h ~ Mass + Tail + Tarsus + (1|sex) | -237.14 | 487.81 | 16.33 | 0 |
| 29 | h ~ Tail + Tarsus + Wing + (1|sex) | -237.26 | 488.04 | 16.56 | 0 |
| 7 | h ~ Mass + Tail + (1|sex) | -238.62 | 488.32 | 16.84 | 0 |
| 21 | h ~ Tail + Wing + (1|sex) | -238.76 | 488.6 | 17.12 | 0 |
| 11 | h ~ Mass + Tarsus + (1|sex) | -238.93 | 488.93 | 17.45 | 0 |
| 3 | h ~ Mass + (1|sex) | -240.4 | 489.51 | 18.03 | 0 |
| 25 | h ~ Tarsus + Wing + (1|sex) | -239.41 | 489.89 | 18.41 | 0 |
| 17 | h ~ Wing + (1|sex) | -240.9 | 490.51 | 19.03 | 0 |
| 13 | h ~ Tail + Tarsus + (1|sex) | -240.75 | 492.58 | 21.1 | 0 |
| 5 | h ~ Tail + (1|sex) | -242.43 | 493.56 | 22.09 | 0 |
| 9 | h ~ Tarsus + (1|sex) | -243.45 | 495.6 | 24.13 | 0 |
| 1 | h ~ (1|sex) | -245.58 | 497.58 | 26.1 | 0 |