







The following is the established format for referencing this article:
Analla, M., P. Fernández-Rodríguez, N. Martínez-Medina, and C. Azorit. 2022. Sexing Eurasian Eagle Owls by external body and skeletal measurements. Journal of Field Ornithology 93(4):1.ABSTRACT
The aim of this study was to obtain an effective tool for sex determination in Eurasian Eagle Owls (Bubo bubo), a species without marked sexual dimorphism, by taking external body and bone measurements. Thirty-one individuals were used for this purpose, 16 males and 15 females, and 42 measurements were obtained: 19 external and 23 bone measurements. Three discriminant analyses were carried out: the first with external measurements only, a second one with bone measurements only, and a third combining both. The validation of the classification was performed by cross-validation. The first analysis led to a discrimination function with five external measures, with a Wilk’s Lambda of 0.064 and 100% of the individuals classified correctly. The second analysis led to a discrimination function with two bone measures, with a Wilk’s Lambda of 0.282 and 90% of individuals correctly classified. The third analysis led to a discrimination function with six external and two bone measures, with a Wilk’s Lambda of 0.022 and 100% of individuals classified correctly. The study has shown that external measurements are more efficient than bone measurements, and likely sufficient for sexing. Nevertheless, bone measurements can provide complementary information, although alone they are insufficient for sexing owls, given that the length of the ulna and the nose bones were the only useful bone measurements. It can be generalized that this work provides an optimal sexing method, useful in ornithological ecology, clinical and forensic medicine, as well as in archaeozoological studies.RESUMEN
INTRODUCTION
Sex determination is central to numerous studies in avian biology, such as foraging ecology (Weimerskirch et al. 2009), dispersal ecology (Clarke et al. 1997, Végvari et al. 2018), or evolutionary ecology (Badyaev and Hill 2003). Sexual dimorphism in plumage is characteristic of many bird species, but in other species the sex of individuals is not so obvious and must be determined by corporal biometry (Muriel et al. 2010, García et al. 2013) or using blood samples (Muriel et al. 2010, Kroczak et al. 2021). Most raptors are dimorphic in size, which allows the development of sex determination methods based on morphometric data (Delgado and Penteriani 2004).
The Eurasian Eagle Owl (Bubo bubo) is one of the largest Iberian nocturnal birds of prey belonging to the order of Strigiformes. It is a common species in Asia or Africa, north-eastern Europe, as well as in the Mediterranean area of the Iberian Peninsula. It inhabits steep rocky areas with sparse forest, difficult to access for humans and predators. It has a length of –73 cm and a wingspan of 138 –170 cm (Svensson and Mullarney 2010). This raptor has a cryptic plumage of earthy ochre with darker flecks. Adults are characterized by a rounded head, large orange eyes, black bill, legs and claws feathered to the nails, false ears composed of several movable feathers, and a white throat shown when singing (Peterson and Mountfort 1995, Svensson and Mullarney 2010). It is a monomorphic species that does not show sexual dimorphism at the plumage level, making it difficult to manage and study (Branson et al. 1994). Apparently, females tend to be larger than males (Penteriani and Delgado 2008), but despite this, size differences are often not so evident, making it impossible to differentiate between males and females in the wild until hearing their calls, which are acoustically different for males and females (Penteriani 1996).
Besides knowing the biology and ecology of the species, accurate sex identification is essential for many bird studies, including brood sex ratios (Benito and Gonzalez-Solis 2007), sex differences in migration timing (Vegvari et al. 2018), sex-biased behavior (Pusey 1987), and sex differences in physiological responses (Pampori and Shapiro 1993, Benito and Gonzalez-Solis 2007). Obtaining knowledge that helps to differentiate males from females becomes almost a necessity for population management and clinical treatment of injured or sick animals. The correct administration of drugs and treatments in breeding or recovery centers, or veterinary clinics where there is an increasing demand for biomedical knowledge in falconry and/or exotic birds, brings with it the need to determine sex. When treating a particular animal, the sex of the animal must be taken into account, given that drugs and treatments differ, not only between different species (Miller et al. 2019), but also between males and females of the same species (Pampori and Shapiro 1993, Anderson 2008, Crisol-Martinez et al. 2016, Gochfeld 2017). On the other hand, another practical utility would be to have simple and reliable procedures for sex determination in the wild, during scientific ringing sessions and population census studies involving capture and marking, or radio-tracking. In addition, there is currently no reference for the identification of the species using skeletal remains on the Iberian Peninsula (Penteriani and Delgado 2008), which would also be covered by this research.
The use of traditional biometrics has been considered a rapid and effective tool for sex determination in other birds, such as Rosy Starlings (Pastor roseus; Zenatello and Botond 2005), Black-headed Gull (Chroicocephalus ridibundus; Palomares et al. 1997), Zenaid Turtle Dove (Zenaida aurita; Dechaume-Moncharmont et al. 2011), penguins or sandpipers (Palestis et al. 2012, Polito et al. 2012, Gates et al. 2013), Iberian raptors in general (Martínez Climent et al. 2002, García et al. 2013), or nocturnal raptors (García Matarranz 2013). Therefore, it can be very useful for many fields of study on species without marked sexual dimorphism such as the Eagle Owl (Delgado and Penteriani 2004). Biometry makes it possible to identify and quantify differences in size and morphology that are not detectable to the naked eye. On the other hand, discriminant function analyses have been widely used for sex determination in bird species with monomorphic plumage (Scolaro et al. 1983, Maron and Myers 1984, Hanners and Patton 1985, Malacalaza and Hall 1988). However, some authors draw attention to the need to develop predictive equations that include a validation that quantifies the reliability and accuracy of these equations using procedures such as Jackknife cross-validation (Dechaume-Moncharmont et al. 2011). The aim of this work is to develop reliable discriminant equations for sex determination from biometric measurements using external/skeletal bone parameters.
METHODS
Origin of the specimens
The material under study consisted of 31 Eagle Owl specimens from wildlife recovery centers in Andalusia. Birds were brought to the centers for various reasons, usually accidents or collisions, but died during treatment. Their carcasses were immediately frozen pending further processing. Necropsies and measurements were performed by a single investigator in a blind process. Sex was determined by gonadal identification during necropsy of the carcasses, of which 16 were sexed as males and 15 as females. A total of 42 external body and bone measurements were taken. The bone material is deposited in the Vertebrate Laboratory of the University of Jaén, Spain.
External measurements
Nineteen external measurements were taken from the complete carcasses. Prior to the necropsies, a list of measurements and necropsy data was prepared in order to avoid errors and to establish a standardization of the different tasks in each necropsy session. Biometric and bone measurements were taken with the help of two calipers of different sizes (±0.1 mm; Bortolotti 1984). All measurements have been recorded and defined in Table 1 and represented in Figure 1.
Bone measurements
The bones were recovered and prepared according to the hot maceration method (Rodríguez-Palomo and Ramírez Zamora 2009), and 23 bone measurements were taken with the two calipers (±0.1 mm). These measurements are described in Table 2 and represented in Figures 2 and 3.
Data analysis
Discriminant analyses were performed with the external measurements alone, with the bone measurements alone, and then with both types of measurements. In this way, three discriminant functions for the distinction between the two sexes were obtained. The program excluded those individuals that had a missing value in at least one variable, so the number of individuals is not always the same. Statistical analysis was performed by group (males and females). Wilks’ method was used and equal a priori probabilities were assumed. The input F-value was 3.86 and the output value was 2.71. These are the default values of IBM’s Statistical Package for Social Sciences (SPSS; see https://www.ibm.com/products/spss-statistics).
The stepwise technique was used to monitor how and why the measures used in the different analyses were chosen. The standardized coefficient matrix allowed the contribution of each variable in the function to be assessed, whereas the structure matrix revealed the correlation coefficients between each measure and the discriminant function. Analyses were then performed using Student’s T test to highlight the measures that were selected for the discriminant functions; and the degree of correlation between them was determined. SPSS software was used for all statistical analyses.
RESULTS
Table 3 recapitulates the variables chosen to form the discriminant equations using only external measurements, only bone measurements, or both. Table 4 summarizes the cross-validation of the use of these equations. Table 5 summarizes the result of the Student’s T-test, applied to the variables retained for the discriminant functions in each of the three analyses. Table 6 reports the correlations between the variables retained in the three analyses. In all three analyses, the discrimination function was highly correlated with the chosen variables: 0.976 for external measurements, 0.848 for bone measurements, and 0.989 when using both. The Wilks’ Lambda value, indicating the percentage of variability not attributed to sex, was low or very low in all three analyses: 6.4%, 28.2%, and 2.2% for external measurements, bone measurements, and both, respectively. In the three analyses there was a very little overlap between males and females. The difference between the centroids of the two groups (male vs. female) was highly significant (p < 0.001) in all three cases (-3.654 vs. 3.654, -1.433 vs. 1.653, and -6.696 vs. 6.138, for males vs. females, in the external, bone, and both analyses respectively).
Equations from external measurements
From the analysis of the external measurements, a function with five variables was obtained (see Table 3), which correctly classified 96.3% of the individuals at hand, assigning erroneously one male as a female. On the other hand, cross-validation indicated 100% correct classification (Table 4). The variables with the highest contribution were the forearm (ABR) followed by the L1 claw (GL1): 1.477 and 1.155, respectively. The variable with the lowest contribution was the tail (CL), with 0.569, although it had the highest correlation with the discriminant function (0.49). The only variable with a negative contribution was eye-ring length (OL), with a value of -0.652 and the lowest correlation with the discriminant function: -0.012 (Table 3).
Equations from bone measurements
In the analysis of bone measurements, a function based on two variables was obtained (Table 3), which classified correctly 93.5% of the available individuals, assigning incorrectly two females as males. Meanwhile, cross-validation indicated 90.3% correct classification, where 87.5% of males and 93.3% of females were classified correctly (Table 4). The two variables chosen were nostril (NAH), with a contribution of 0.546 and a correlation of 0.365 with the discriminant function, and ulna bone length (CBH), with a contribution of 0.949 and a correlation of 0.844 with the discriminant function.
Equations from both measures
From the analysis of the external and bone measurements, a final function was obtained that pooled eight of the measurements (Table 3) and correctly classified 96.0% of the available individuals, identifying incorrectly one female as male. Meanwhile, cross-validation indicated 100% correct classification (Table 4). However, the variables OL (eye-ring length) and TB (tibio-tarsus musculus length) chosen when using external measurements alone were now replaced by the measurements beak to nose length (PL1), skull length (CR), and folded wing (APL). Their contribution was 0.588, -1.203, and 0.823, and their correlation with the discriminant function was 0.195, 0.056, and 0.189, respectively. The bone measurements chosen during the second analysis were not included. This time the measurements chosen were the width of the jugal bone (YAH) and the width of the sternum, contributing with -0.956 and 1.272, and with correlations of 0.065 and 0.121, respectively. The variable with the highest contribution was the forearm with 1.405 and a correlation with the discriminant function of 0.192.
When comparing between the two sets of analysis, in the analysis from external measurements, the first variable included was the tail (CL). In the second step, the length of the beak to the nose (PL1) was included. But when the forearm (ABR) was entered in the fourth step, the F of PL1 ran outside the output limit (F = 2.71) and was then removed. This is most likely due to the strong and highly significant correlation between ABR and PL1, with a value of 0.71 (Table 6). The correlation implies redundancy of information, which leads to the elimination of the redundant variable. Thereafter, the forearm (TB) was included in the fifth step; and finally, the eye-ring length (OL) was added. In the analysis from bone measurements, the length of the ulna bone (CBH) was included in the first step, where F-value was the highest at 47.274. It was followed by the nostril (NAH) with an F-value equal to 6.492. In the third step all remaining measurements showed an F-value lower than the cut-off value of 3.84, and then no further variables were entered. In the analysis with both types of measurements, the tail (CL), with the highest F (86.436), was entered first. Then sternal width (EAH) was entered, as it showed the highest F during the second step (14.516). CBH and NAH, which were the two measures included in the discriminant function bone only, were not entered because they showed lower F values: 11.949 and 0.541, respectively. In fact, CBH shows a high correlation (0.688) with EAH and a moderate correlation (0.504) with CL, both highly significant (Table 6), resulting in redundant information. Thereafter, APL, GL1, CR, and ABR were entered, in this order. The YAH measure was entered at the seventh step and PL1 last, at the eighth step. The eye-ring length (OL) was not entered because the highest F-value it obtained was 2.663 in the seventh step. In fact, this variable is not correlated with any of the others; but it does not seem to be affected by sex, because the t-test was not significant (Table 5). This is probably the reason for not including it in the discriminant function.
DISCUSSION
According to the results of cross-validation and bearing in mind the canonical correlation as well as the reduction of variability, the inclusion of bone measurements together with the external ones did not notably improve sex discrimination. The canonical correlation increased from 0.976 to 0.989, and the reduction of unexplained variability went from 6.4% to 2.2%, which does not seem to compensate for the tiresome work necessary to obtain bone measurements. In fact, the canonical correlation was only 0.848 and the reduction in unexplained variability was relatively high (28.2%) when using bone measurements alone. Considering the laboriousness of bone preparation, it may not be worthwhile to include their measurements in sex discrimination when external measures are available. In this sense, measurements that proved to be sex-determining are easier to obtain in the field and could in turn be used to identify the sex of other Strigiformes (e.g., Calvo and Bolton 1997, Renner and Davis 1999, Leader 2000). According to the centroid values for the three discriminant analyses, those belonging to males always showed smaller values than females, which corroborate the initial idea that females are larger than males (Delgado and Penteriani 2004). These authors also found that the second claw, forearm, length of the exposed culmen, and beak depth were the most dimorphic variables.
The study has shown that external measurements are effective tools for distinguishing between sexes for Eagle Owls. External variables used in this study are easy to measure in the field and have been shown to be good predictors of sex in several bird species (Delgado and Penteriani 2004, Martínez-Abrain et al. 2006). However, some of the variables selected in this study, e.g., length of tail or wing measurement as folded wing or length of the third primary feather, are influenced by molting and by specimen or feather condition, and may lead to inconclusive results. The length of the forearm has been used successfully for sex determination in several species, such as Eurasian Eagle-Owls (Delgado and Penteriani 2004), Spanish Imperial Eagles (Aquila adalbeti; Ferrer and De Le Court 1992), and Bonelli’s Eagles (Aquila fasciata; García et al. 2013).
In contrast, bone measurements did not add relevant information when included with the external measurements in the same function, and they were also less effective than these latter when used alone for discrimination between sexes. However, such measurements should not be definitively discarded, as they can always help improve the identification of carcasses of unknown sex when the other effective measurements are lacking. In fact, many carcasses collected in the field are incomplete. In such cases, retaining only a few measurements for discrimination and discarding the others can complicate the identification of the sex of an incomplete carcass, because some of the necessary measurements could be lacking. The most practical approach should be to have this database ready to perform and recalculate a new discrimination function with the available measurements taken on the carcass or part of the carcass, at the time of collection. This work provides an optimal sexing method, useful for clinical and forensic medicine, as well as for archaeological studies.
An effective tool for sex determination in Eurasian Eagle Owls was obtained by taking measurements of external body parameters and bones. Two discriminant functions were able to determine the sex of Eagle Owls with 100% of individuals correctly classified, one of these functions only by measuring external body parameters. This function is very appropriate in contexts in which live animals are studied for clinical treatment, such as those admitted to wildlife recovery centers, or those captured for ringing or studies on the ecology of the species, as this is a species with little dimorphic variability, and where weight of the animal can create a lot of uncertainty and is not sufficient to determine the sex of the specimen. Bone measurements provide complementary information, with ulna and nose bone length being the most useful bone measurements for sexing Eagle Owls. In any case, the discriminant function that uses only bone measurements also achieves acceptable results with 90% of individuals correctly classified, so it could be used in archaeological or forensic contexts where only the skeleton of the animal is preserved. This work provides a practical and optimal sexing method, useful in zoology, ornithological ecology, and clinical and forensic medicine, as well as in archaeozoological studies.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.AUTHOR CONTRIBUTIONS
MA: writing, data curation, methodology, statistical analyses, discussing results. NMM: specimen preparation (necropsies and bone-skeleton conditioning), specimen study, graphic design, hypotheses, writing and editing, discussing results. PFR: writing and editing, hypotheses, discussing results. CA: conceptualizing the study, funding acquisition, project administration, specimen study, discussing results, writing, and corresponding author. All authors drafted the manuscript and revised it for final approval.
ACKNOWLEDGMENTS
We thank the local wildlife management authorities for their help in collecting and providing specimens, and especially Mrs. Elena Zarco from CREA Quiebrajano Jaén, Spain. This research has been funded by the University of Jaén (Programa Operativo FEDER Andalucía 2014–2020) and the Research Group RNM175.
DATA AVAILABILITY
The data/code that support the findings of this study are available on request from the corresponding author, CA. Ethical approval for this research study was granted by the bioethics committee of the University of Jaén and this study has been possible thanks to the authorization and transfer of owl carcasses from the wildlife recovery centre of the Consejería de medio ambiente y ordenación del territorio, Junta de Andalucía, Spain, with no:B 227/2/2015 SGMN/Dpto GEO y BIO/ASG/RASL.
LITERATURE CITED
Anderson, G. D. 2008. Gender differences in pharmacological response. International Review of Neurobiology 83:1-10. https://doi.org/10.1016/S0074-7742(08)00001-9
Badyaev, A. V., and G. E. Hill. 2003. Avian sexual dichromatism in relation to phylogeny and ecology. Annual Review of Ecology, Evolution, and Systematics 34(1):27-49. https://doi.org/10.1146/annurev.ecolsys.34.011802.132441
Benito, M. M., and J. González-Solís. 2007. Sex ratio, sex-specific chick mortality and sexual size dimorphism in birds. Journal of Evolutionary Biology 20(4):1522-1530. https://doi.org/10.1111/j.1420-9101.2007.01327.x
Bortolotti, G. R. 1984. Criteria for determining age and sex of nestling Bald Eagles. Journal of Field Ornithology 55(4):467-481. https://sora.unm.edu/sites/default/files/journals/jfo/v055n04/p0467-p0481.pdf
Branson, W. R., G. J. Harrison, and L. R. Harrison. 1994. Avian medicine: principles and application. Wingers, Lake Worth, Florida, USA.
Calvo, B., and M. Bolton. 1997. Sexing shags Phalacrocorax aristotelis from external measurements using discriminant analysis. Ringing and Migration 18(1):50-56. https://doi.org/10.1080/03078698.1997.9674140
Clarke, A., B. Sæther, and E. Røskaft. 1997. Sex biases in avian dispersal: a reappraisal. Oikos 79(3):429. https://doi.org/10.2307/3546885
Crisol-Martínez, E., L. T. Moreno-Moyano, N. Wilkinson, et al. 2016. A low dose of an organophosphate insecticide causes dysbiosis and sex-dependent responses in the intestinal microbiota of the Japanese quail (Coturnix japonica). PeerJ 4:e2002. https://doi.org/10.7717/peerj.2002
Dechaume-Moncharmont, F. X., K. Monceau, and F. Cezilly. 2011. Sexing birds using discriminant function analysis: a critical appraisal. Auk 128(1):78-86. https://doi.org/10.1525/auk.2011.10129
Delgado, M., and V. Penteriani. 2004. Gender determination of Eurasian Eagle-owls (Bubo bubo) by morphology. Journal of Raptor Research 38(4):375-377. https://sora.unm.edu/sites/default/files/journals/jrr/v038n04/p00375-p00377.pdf
Ferrer, M. and C. de le Court. 1992. Sex determination in the Spanish Imperial Eagle. Journal of Field Ornithology 63(3):359-364. https://www.researchgate.net/publication/285843808
García, V., R. Moreno-opo, and A. Tintó. 2013 Sex differentiation of Bonelli’s Eagle Aquila fasciata in western Europe using morphometrics and plumage colour patterns. Ardeola 60(2):261-277. https://doi.org/10.13157/arla.60.2.2013.261
García Matarranz, V. 2013. Guía de identificación de rapaces ibéricas por restos óseos (1ª parte, grandes rapaces). Ministerio de Agricultura, Alimentación y Medio Ambiente, Madrid, Spain. https://www.miteco.gob.es/es/biodiversidad/temas/conservacion-de-especies/guiarapacesrestososeos1parte_tcm30-198046.pdf
Gates, H. R., S. Yezerinac, A. N. Powell, P. S. Tomkovich, O. P. Valchuk, and R. B Lanctot. 2013. Differentiation of subspecies and sexes of Beringian Dunlins using morphometric measures. Journal of Field Ornithology 84(4):389-402. https://doi.org/10.1111/jofo.12038
Gochfeld, M. 2017. Sex differences in human and animal toxicology: toxicokinetics. Toxicologic Pathology 45(1):172-189. https://doi.org/10.1177/0192623316677327
Kroczak, A., M. Wołoszyńska, H. Wierzbicki, M. Kurkowski, K. A. Grabowski, T. Piasecki, L. Galosi, and A. D. Urantówka. 2021. New bird sexing strategy developed in the order Psittaciformes involves multiple markers to avoid sex misidentification: debunked myth of the universal DNA marker. Genes 12(6):878. https://doi.org/10.3390/genes12060878
Hanners, L. A., and S. R. Patton. 1985. Sexing Laughing Gulls using external measurements and discriminant analysis. Journal of Field Ornithology 56:158-164. https://sora.unm.edu/sites/default/files/journals/jfo/v056n02/p0158-p0164.pdf
Leader, N. 2000. Predicting the sex of Blackstarts (Cercomelam elanura) by discriminant analysis. Israel Journal of Zoology 46(2):149-154. https://www.tandfonline.com/doi/abs/10.1560/8HFR-831W-0URC-7WEV
Malacalaza, V. E., and M. A. Hall. 1988. Sexing adult King Cormorants (Phalacrocorax albiventer) by discriminant analysis. Colonial Waterbirds 11(1):32-37. https://doi.org/10.2307/1521167
Maron, J. L., and J. P. Myers. 1984. A discrimination and evaluation of two techniques for sexing Wintering Sanderlings. Journal of Field Ornithology 55(3):336-342. https://www.jstor.org/stable/4512916
Martínez-Abraín, A., D. Oro, A. Velando, M. Genovart, C. Gerique, M. A. Bartolomé, B. Sarzo, and E. Villuendas. 2006. Morphometric similarities between central and peripheral populations of the European Shag Phalacrocorax aristotelis. Marine Ornithology 34(1):21–24. https://www.researchgate.net/publication/249998711
Martínez Climent, J. A., I. Zuberogoitia, and R. Alonso. 2002. Rapaces nocturnas: guía para la determinación de la edad y el sexo en las estrigiformes ibéricas. Montícola, Madrid, Spain. https://brinzal.org/en/rapaces-nocturnas-guia-para-la-determinacion-de-la-edad-y-el-sexo-en-las-estrigiformes-ibericas/
Miller, E., N. Lambetski, and P. Calle. 2019. Fowler’s zoo and wild animal medicine current therapy, volume 10. Elsevier, St Louis, Missouri, USA.
Muriel, R., E. Casado, D. Schmidt, C. Calabuig, and M. Ferrer. 2010. Morphometric sex determination of young Ospreys Pandion haliaetus using discriminant analysis. Bird Study 57(3):336-343. https://doi.org/10.1080/00063651003674953
Palestis, B. G., I. C. T. Nisbet, J. J. Hatch, P. Szczys, and J. A. Spendelow. 2012. Morphometric sexing of Northwest Atlantic Roseate Terns. Waterbirds 35(3):479-484. https://doi.org/10.1675/063.035.0312
Palomares, L. E., B. E. Arroyo, J. Marchamalo, J. J. Sainz, and B. Voslamber. 1997. Sex- and age-related biometric variation of Black-headed Gulls Chroicocephalus ridibundus in Western European populations. Bird Study 44(3):310-317. https://doi.org/10.1080/00063659709461066
Pampori, N. A., and B. H. Shapiro. 1993. Sexual dimorphism in avian hepatic monooxygenases. Biochemical Pharmacology 46(5):885-890. https://doi.org/10.1016/0006-2952(93)90498-L
Penteriani, V. 1996. The Eagle Owl. Calderini, Bologna, Italy.
Penteriani, V., and M. M. Delgado. 2008. Owls may use faeces and prey feathers to signal current reproduction. PLoS ONE 3(8):e3014. https://doi.org/10.1371/journal.pone.0003014
Peterson, R. T., and G. Mountfort. 1995. Guía de campo de las aves de España y de Europa. Fifth edition. Omega, Barcelona, Spain.
Polito, M. J., G. V. Clucas, T. Hart, and W. Z. Trivelpiece. 2012. A simplified method of determining the sex of pygoscelis penguins using bill measurements. Marine Ornitology 40:89-94. http://marineornithology.org/PDF/40_2/40_2_89-94.pdf
Pusey, A. E. 1987. Sex-biased dispersal and inbreeding avoidance in birds and mammals. Trends in Ecology and Evolution 2(10):295-299. https://doi.org/10.1016/0169-5347(87)90081-4
Renner, M., and L. S. Davis. 1999. Sexing Little Penguins Eudyptula minor from Cook Strait, New Zealand using discriminant function analysis. Emu 99(1):74-79. https://doi.org/10.1071/MU99009C
Rodríguez Palomo, D., and J. Ramírez Zamora. 2009. Técnica de conservación de huesos en peróxido de hidrógeno. Medicina Legal de Costa Rica 26(2):117-123. https://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S1409-00152009000200006
Scolaro, J. A., M. A. Hall, and I. M. Ximenez. 1983. The Magellanic Penguin (Spheniscus magellanicus): sexing adults by discriminant analysis of morphometric characters. Auk 100(1):221-224. https://doi.org/10.1093/auk/100.1.221
Svensson, L., and K. Mullarney. 2010. Guía de aves: España, Europa, región Mediterránea. Omega, Barcelona, Spain.
Végvári, Z., G. Katona, B. Vági, R. P. Freckleton, J. M. Gaillard, T. Székely, and A. Liker. 2018. Sex-biased breeding dispersal is predicted by social environment in birds. Ecology and Evolution 8(13):6483-6491. https://doi.org/10.1002/ece3.4095
Weimerskirch, H., M. Le Corre, H. Gadenne, D. Pinaud, A. Kato, Y. Ropert-Coudert, and C.-A. Bost. 2009. Relationship between reversed sexual dimorphism, breeding investment and foraging ecology in a pelagic seabird, the masked booby. Oecologia 161:637–649. https://doi.org/10.1007/s00442-009-1397-7
Zenatello, M., and J. K. Botond. 2005. Biometrics and sex identification of the Rose Coloured Starling Sturnus roseus. Ringing and Migration 22:163-166. https://doi.org/10.1080/03078698.2005.9674326

Fig. 1
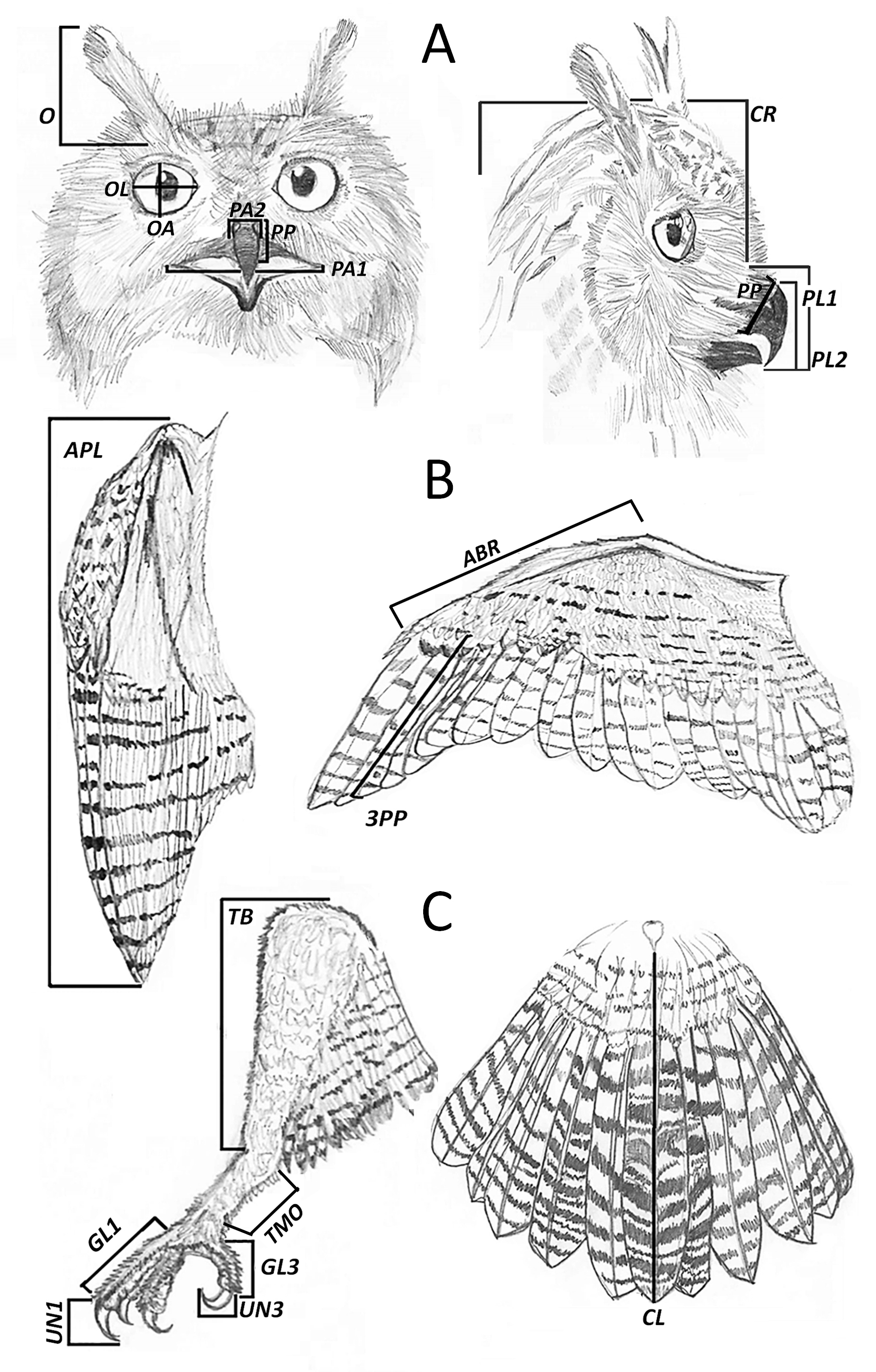
Fig. 1. (A) External measurements of the head in frontal position and in lateral position: beak width from commissure to commissure (PA1), beak width at wax (PA2), beak depth (PP), eye width and length (OL and OA), length of false ear (O), length of skull (CR), length of beak (PL1), and length of beak to skull (PL2). (B) Wing measurements: folded wing (APL), forearm (ABR), length of the third primary feather, corresponding to P8 (3PP). (C) External measurements of the leg and tail: length of tibia (TB), length of tarsus (TMO), length of toes 1 and 3 (GL1 and GL3), length of nails 1 and 3 (UN1 and UN3), and length of tail (CL).
Fig. 2
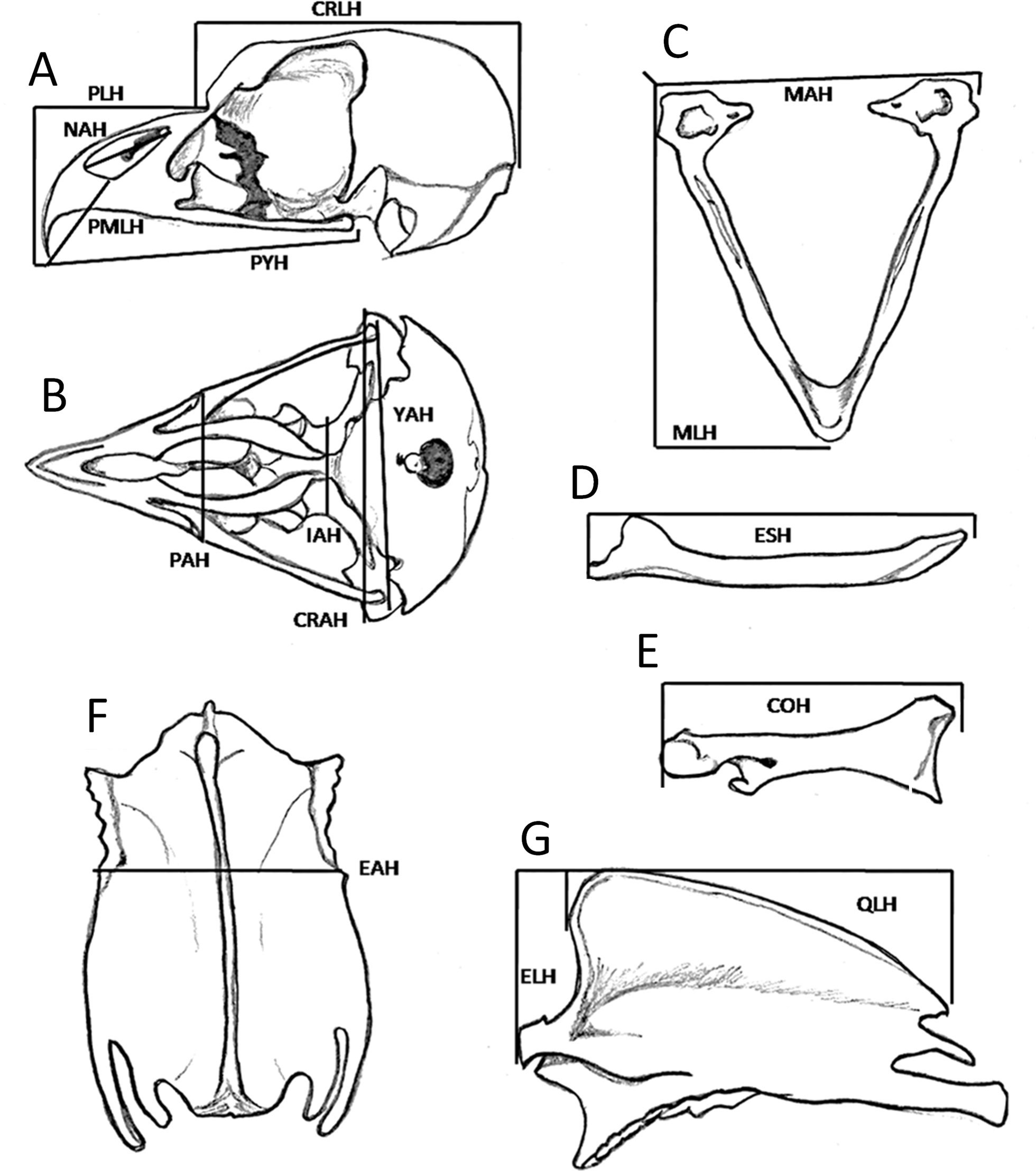
Fig. 2. Cranial measurements, lower jaw, scapula, coraoid, and sternum: (A) skull in lateral position (CRLH, PLH, PMLH, NAH, PYH); (B) skull in ventral position (CRH,YAH, IAH, PAH); (C) lower jaw (MAH, MLH); (D) scapula (ESH); (E) coracoid (COH); (F) ventral sternum (EAH); (G) lateral (ELH, QLH).
Fig. 3
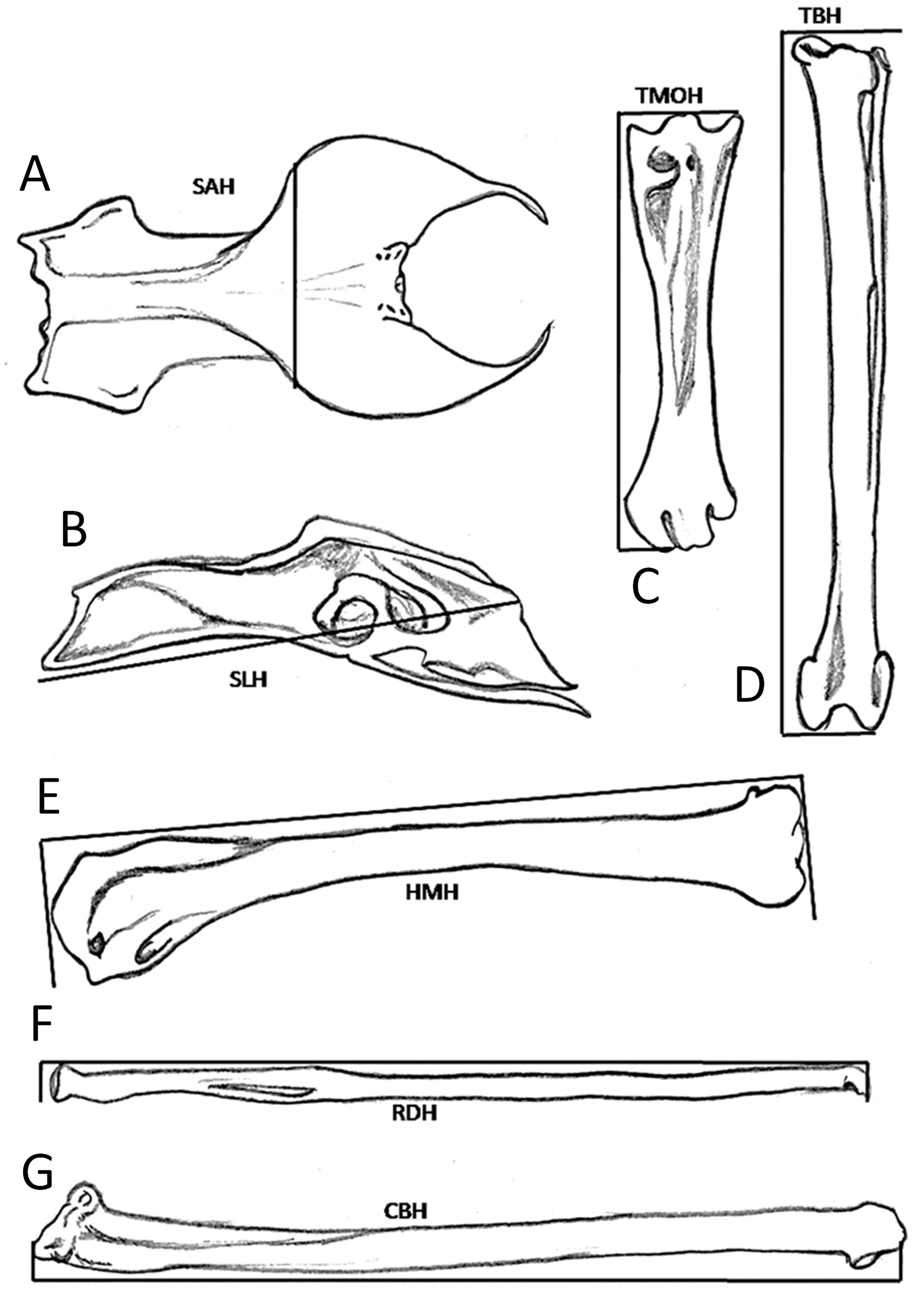
Fig. 3. Measurements of the (A) ventral synsacrum (SAH); (B) lateral synsacrum(SLH); (C) tarsus-metatarsus (TMOH); (D) tibia-peroneus (TBH); (E) humerus (HMH); (F) radius (RDH); and (G) ulna (CBH).
Table 1
Table 1. Abbreviations and definitions of external measures.
| Definition | |
| O | Length of feathers resembling false “ears” |
| OL | Length of eye-ring |
| OA | Width of eye-ring |
| PA1 | Beak width from commissure to commissure corresponding to the full opening |
| PA2 | Width of beak at wax height |
| PP | Beak depth |
| PL1 | Length of beak to the beak wax around the nostrils |
| PL2 | Length from beak to its articulation to skull |
| CR | Length of skull measured from the articulation of the beak to the base of the skull |
| APL | Length of folded wing |
| ABR | Length of the forearm corresponding to the ulnar-radius bones |
| 3PP | Third primary feather which according to wing nomenclature corresponds to the P8 |
| TB | Length corresponding to the tibio-tarsus bone and tibio-tarsus musculus length |
| TMO | Tarsus-metatarsus length to claw joint |
| GL1 | Length of claw L1 from toe insertion to start of claw 1 |
| GL3 | Length of claw L3 from toe insertion to start of nail 3 |
| UN1 | Length of claw L1 to toenail Insertion 1 |
| UN3 | Length of claw L3 to insertion of digit 3 |
| CL | Length of tail from uropygial gland to the end of the central remiges feathers |
Table 2
Table 2. Abbreviations and definitions of bone measures.
| Definition | |
| CRLH | Length from beak joint to basioccipital crests of the skull |
| PLH | Length of beak |
| NAH | Length of nostrils |
| PMLH | Length of premaxilla |
| PYH | Length from beak to jugal square |
| YAH | Width between the two jugal squares |
| IAH | Interorbital width |
| PAH | Width of beak at maxillary level |
| CRAH | Width of the skull at the level of the ear pinnae |
| MAH | Width of the lower jaw at the level of the angulars |
| MLH | Length of lower jaw from dental to angular |
| ESH | Length of scapula |
| COH | Length of coracoid |
| EAH | Width of the sternum at the level of the second last rib |
| QLH | Length from caudal notch to keel |
| ELH | Length from caudal notch to sternum |
| SAH | Width of the synsacrum at the level of the ischium |
| SLH | Length of synsacrum from the extreme ileum to the ischium |
| TMOH | Tarsus-metatarsus length |
| TBH | Tibio-tarsus length |
| HMH | Humerus length |
| RDH | Radius length |
| CBH | Ulna bone length ulna bone length |
Table 3
Table 3. Variables picked up by the discriminant analysis, with their standardized coefficients and their correlation with the discriminant function, for each set of variables.
| Analysis | Variables† | Definition | Coefficients | Correlation |
| Body variables | CL GL1 ABR TB OL |
Tail Claw L1 Forearm Tibio-tarsus muscle Length of eye-ring |
0.569 1.155 1.477 0.632 -0.652 |
0.490 0.147 0.321 0.130 -0.012 |
| Bones variables | CBH NAH |
Ulna bone Length of nostril |
0.949 0.546 |
0.844 0.365 |
| Both | CL EAH APL GL1 CR ABR YAH PL1 |
Tail Sternum width Folded wing Claw L1 Length of skull Forearm Width of jugal bone Beak to nose length |
1.133 1.272 0.823 1.253 -1.203 1.405 -0.956 0.588 |
0.302 0.121 0.187 0.076 0.056 0.192 0.065 0.195 |
| † Body measurements are in bold and bone measurements in italic. Measurements are listed according to their order of entry in the discriminant function. | ||||
Table 4
Table 4. Validation of the discrimination between sexes, using each set of variables.
| Analysis | Predicted† | Total | |||
| Male | Female | ||||
| Body variables | Original | Male | 14 | 0 | 14 |
| Female | 1 | 12 | 13 | ||
| Cross validated | Male | 14 | 0 | 14 | |
| Female | 0 | 13 | 13 | ||
| Bone variables | Original | Male | 14 | 2 | 16 |
| Female | 0 | 15 | 15 | ||
| Cross validated | Male | 14 | 2 | 16 | |
| Female | 1 | 14 | 15 | ||
| Both | Original | Male | 12 | 1 | 13 |
| Female | 0 | 12 | 12 | ||
| Cross validated | Male | 13 | 0 | 13 | |
| Female | 0 | 12 | 12 | ||
| †Numbers of individuals misclassified are italicized | |||||
Table 5
Table 5. Mean, standard deviation, and number of observations of the variables chosen by the discriminant analysis: Beak to nose length (PL1), skull length (CR), eye-ring length (OL), folded wing (APL), forearm (ABR), tibia-peroneus length (TB), claw L1 (GL1), tail (CL), width of the jugal bone (YAH), nostril (NAH), sternal width (EAH), and ulna bone length (CBH).
| Variable (mm)† | Males | Females | t-value | p-value |
| PL1 | 48.257 ± 1.832 (16) |
51.879 ± 2.406 (15) |
-4.789 | <0.001 |
| CR | 57.723 ± 7.940 (16) |
61.470 ± 2.846 (15) |
-1.778 | 0.086 |
| OL | 27.458 ± 1.109 (16) |
26.940 ± 1.147 (15) |
1.298 | 0.204 |
| APL | 434.060 ± 16.62 (16) |
455.500 ± 16.02 (15) |
-3.716 | <0.001 |
| ABR | 187.270 ± 4.29 (16) |
199.460 ± 9.11 (15) |
-4.840 | <0.001 |
| TB | 142.780 ± 13.76 (16) |
154.880 ± 15.57 (15) |
-2.330 | 0.027 |
| GL1 | 44.260 ± 3.381 (14) |
48.564 ± 5.518 (13) |
-2.397 | 0.024 |
| CL | 240.820 ± 13.03 (16) |
254.560 ± 11.48 (15) |
-3.164 | 0,004 |
| YAH | 59.627 ± 3.265 (16) |
62.445 ± 2.025 (13) |
-2.789 | 0.009 |
| NAH | 17.008±1.370 (16) |
18.386 ± 1.008 (15) |
-3.240 | 0.003 |
| EAH | 42.914 ± 1.860 (15) |
45.296 ± 2.031 (15) |
-3.397 | 0.002 |
| CBH | 177.070 ± 5.11 (16) |
190.030 ± 5.68 (15) |
-6.781 | <0.001 |
| † Body measurements are in bold, bone measurements in italic, and specimen number in brackets. | ||||
Table 6
Table 6. Correlations and number of observations above diagonal and probability of significance below diagonal, obtained among variables chosen by the discriminant analysis: beak to nose length (PL1), skull length (CR), eye-ring length (OL), folded wing (APL), forearm (ABR), tibia-peroneus length (TB), claw L1 (GL1), tail (CL), width of the jugal bone (YAH), nostril (NAH), sternal width (EAH), and ulna bone length (CBH).
| ‡ | PL1 | CR | OL | APL | ABR | TB | GL1 | CL | YAH | NAH | EAH | CBH |
| PL1 | 0.307 31 |
-0.118 31 |
0.578** 31 |
0.710*** 31 |
0.325 31 |
0.276 31 |
0.613*** 31 |
0.433* 29 |
0.163 31 |
0.632*** 30 |
0.773*** 31 |
|
| CR | 0.093 | -0.208 31 |
-0.088 31 |
0.187 31 |
-0.146 31 |
0.289 27 |
0.014 31 |
0.248 29 |
0.213 31 |
0.193 30 |
0.220 31 |
|
| OL | 0.528 | 0.263 | 0.120 31 |
0.123 31 |
0.037 31 |
-0.007 27 |
-0.072 31 |
-0.196 29 |
-0.282 31 |
0.106 30 |
0.070 31 |
|
| APL | 0.001 | 0.0637 | 0.520 | 0.690*** 31 |
0.426* 31 |
0.221 27 |
0.688*** 31 |
0.286 29 |
0.377* 31 |
0.329 30 |
0.670*** 31 |
|
| ABR | 0.000 | 0.313 | 0.509 | 0.000 | 0.238 31 |
0.059 27 |
0.523** 31 |
0.515** 31 |
0.256 31 |
0.621*** 30 |
0.825*** 31 |
|
| TB | 0.074 | 0.434 | 0.842 | 0.017 | 0.196 | 0.100 27 |
0.382* 31 |
-0.089 29 |
0.166 31 |
0.403* 30 |
0.267 31 |
|
| GL1 | 0.163 | 0.144 | 0.974 | 0.269 | 0.770 | 0.621 | 0.436* 27 |
0.266 26 |
0.324 27 |
0.081 26 |
0.151 27 |
|
| CL | 0.000 | 0.940 | 0.702 | 0.000 | 0.003 | 0.034 | 0.023 | 0.262 29 |
0.334 31 |
0.461* 30 |
0.504** 31 |
|
| YAH | 0.019 | 0.195 | 0.308 | 0.132 | 0.004 | 0.646 | 0.189 | 0.170 | 0.243 29 |
0.257 28 |
0.433* 29 |
|
| NAH | 0.382 | 0.257 | 0.124 | 0.036 | 0.165 | 0.373 | 0.099 | 0.066 | 0.203 | 0.071 30 |
0.379* 30 |
|
| EAH | 0.000 | 0.306 | 0.578 | 0.076 | 0.000 | 0.027 | 0.692 | 0.010 | 0.187 | 0.709 | 0.688*** 30 |
|
| CBH | 0.000 | 0.234 | 0.709 | 0.000 | 0.000 | 0.147 | 0.452 | 0.004 | 0.019 | 0.035 | 0.000 | |
| ‡ Body measurements are in bold and bone measurements in italic. * Significant (p < 0.05). ** Highly significant (p < 0.01). *** Very highly significant (p < 0.001). | ||||||||||||