







The following is the established format for referencing this article:
Zarri, E. C., and T. E. Martin. 2022. A comparison of the breeding biology of the Bornean Whistling-Thrush (Myophonus borneensis) and White-crowned Forktail (Enicurus leschenaulti borneensis). Journal of Field Ornithology 93(3):3.ABSTRACT
Advances in understanding geographic patterns of life history variation depend on documentation of life history traits for species in poorly studied regions. In the tropics, most species have not had their life history traits described. We compared the reproductive biology of two closely related Muscicapids, the Bornean Whistling-Thrush (Myophonus borneensis) and the Bornean subspecies of the White-crowned Forktail (Enicurus leschenaulti borneensis), coexisting in Mount Kinabalu Park in Sabah, Malaysia. Both species specialize in riparian habitats, but whistling-thrushes are more adaptable to human presence and can be found nesting on and foraging around buildings, whereas forktails are more wary of humans. We located and monitored 77 forktail nests and 130 whistling-thrush nests from 2009 to 2020. Mean clutch size was 2.00 ± 0.04 for forktails and 1.89 ± 0.04 for whistling-thrushes. Mean egg mass at lay was 4.07 ± 0.04 g for forktails and 11.65 ± 0.12 g for whistling-thrushes. Forktail incubation period (17.0 ± 0.42 d) was slightly shorter than for whistling-thrush (18.6 ± 0.19 d), but both species had similar rates of incubation attentiveness. Nestling periods were much shorter for the forktail (15.89 ± 0.39 d) than the whistling-thrush (24.00 ± 0.20 d). The shorter nestling period was associated with faster growth and higher provisioning rates for forktails than whistling-thrushes. Whistling-thrushes are larger and better able to protect their nests from predation, yielding much lower daily nest predation rates (0.012 ± 0.002) than in forktails (0.042 ± 0.006). Indeed, whistling-thrushes were themselves predators of nestlings and eggs of other passerines, whereas forktails were not. The larger size and lower predation risk may explain the slower growth and development of whistling-thrushes. Both species are vulnerable to climate change and droughts because of their reliance on riparian habitats.RESUMEN
INTRODUCTION
Life history theory investigates broad patterns of demographic strategies across geographic regions (Lack 1947, Skutch 1949, MacArthur and Wilson 1967, Martin 1996). Documentation of reproductive biology is essential to understanding global patterns and testing life history theories (Martin 1996, 2015, Boyle et al. 2016). Study within the tropics is particularly important because coexisting species can exhibit extensive variation in life history traits (Blake and Loiselle 2008, Martin 2008, 2015, Martin et al. 2017). This variation raises questions about the extent to which tropical strategies can be generalized as different from temperate strategies (Karr et al. 1990, Blake and Loiselle 2008). Consequently, documentation of strategies in tropical species is critical to a fuller understanding of life history similarities and differences among latitudes. Nonetheless, tropical species remain significantly understudied with respect to their breeding life history traits relative to their diversity.
Within the diverse and widespread Muscicapidae family, many genera have been studied in the temperate region. For example, many species in the Ficedula, Luscinia, and Phoenicurus genera have well-described breeding biology (Morgan 1982, Fennici 1984, Porkert and Zajíc 2005, Lu et al. 2011). However, other genera in the family, especially the tropical genera, are poorly studied and much remains to be learned about their reproductive biology. Myophonus (whistling-thrushes) and Enicurus (forktails) are two genera with little published research on their breeding biology, despite the fact that they are common and widespread throughout Asia (Collar 2020a, b). The forktails and whistling-thrushes are sister taxa that form a clade within the Saxicolinae subfamily of Muscicapidae (Sangster et al. 2010, Zuccon and Ericson 2010). Understanding the breeding biology of species within these genera will increase understanding of the demographic strategies of the tropical members of the Muscicapids and add to knowledge of variation within the tropics.
This paper details the breeding biology of two co-occurring species, the Bornean Whistling-Thrush (Myophonus borneensis) and the Bornean subspecies of the White-crowned Forktail (Enicurus leschenaulti borneensis), hereafter whistling-thrush and forktail, respectively (Fig. 1a, b). These species provide a comparison of two poorly studied genera that coexist in the same habitat. They are both montane endemics on the island of Borneo and specialize in riparian habitats (Martin and Mouton 2020). Both species are considered “Least Concern,” but whistling-thrush populations are declining (BirdLife International 2016a). The Bornean subspecies of the forktail is lumped with all other subspecies, so the population status of the Bornean subspecies is unknown (BirdLife International 2016b). However, some phylogenetic analyses have suggested that the Bornean subspecies of the forktail should be considered a separate species (Moyle et al. 2005, 2017, Sheldon et al. 2009). We describe life history traits including nest site characteristics, nesting phenology, morphometrics, and nest survival of these two species. This paper details data on previously undescribed breeding traits and substantially updates data and variation of traits used in previous analyses (e.g., Martin 2015, Martin et al. 2015a, b).
METHODS
Study site
We conducted this study in Kinabalu Park in Sabah, Malaysia (6°05′N, 116°33′E). The primary study site was in montane tropical rainforest at elevations from 1350–1950 m asl. We also collected data on one nest of the whistling-thrush at a high elevation site on Mount Kinabalu at 3200 m asl. Nest searching was conducted from February to June, 2009–2020 at the primary site (Martin and Mouton 2020) and February to June, 2013–2017 at the high elevation site (Mitchell et al. 2020). The primary site was located around the park headquarters, which included a mix of undisturbed natural areas and areas with hotels, restaurants, and lodges. Many buildings were surrounded by forest. Average temperatures at the primary site were 15–18 °C and average rainfall was 2300 mm annually (Kitayama and Aiba 2002).
Nest monitoring and developmental periods
We searched for nests using parental behavior and systematic searching (Martin and Geupel 1993). Once a nest was found, we collected data on elevation and nest substrate, and we visually estimated nest height and substrate height. We used ladders to access high nests. Nest size was measured once building was complete only for nests found during building or egg laying. We measured total height, total diameter, cup depth, and cup diameter using a ruler with an accuracy of 0.1 cm. After nests were no longer active, a few nests from each species were collected, dried, and weighed to obtain nest mass. We monitored active nests every 1–4 days, but daily or twice daily around stage transitions (lay, hatch, and fledge) to obtain exact period lengths (Martin et al. 2015b). The incubation period was calculated as the time between the day the last egg was laid and the day the eggs hatched (Martin et al. 2007, 2015a). All hatching was synchronous. Nestling period was calculated as the length of time between the day of hatching and the day that the last nestling fledged (Martin et al. 2011, 2015b, Martin 2015). Nests found during incubation and nestling periods were backdated using mean incubation and nestling periods to estimate the initiation date (the day the first egg was laid). Nests were considered to have been depredated if the nest was found empty more than two days before the end of the mean nestling period for that species and no evidence of fledglings was detected.
Egg and nestling measurements
For clutch size description, we used clutch size only in nests found prior to clutch completion to ensure no partial loss of eggs as might occur if found later in incubation. Within the first three days of incubation, we weighed the eggs using a portable electronic scale (0.001 g accuracy). Nestlings were measured on days 0 (hatch day), 1, 2, and every other day until fledge at roughly the same time each day for a nest. Nestling mass was measured using a portable electronic scale (0.001 g accuracy), and the tarsus and wing chord were measured using digital calipers (0.01 mm accuracy).
Nest attentiveness and provisioning rates
We measured nest attentiveness during incubation using temperature probes inserted under the nest lining and connected to a data logger that recorded the temperature every 12 seconds for the entire day. Data from the temperature loggers were analyzed using Raven (K. Lisa Yang Center for Conservation Bioacoustics 2016) to calculate on and off bouts and record bout length. We quantified brooding attentiveness and nestling provisioning rates through video recording of nests. Nests were filmed throughout the nestling period using HD video cameras (Canon Vixia HF R800) with 32x zoom set up at least 5 m away from the nest and camouflaged to reduce impacts on parental behavior. Nests were filmed starting around sunrise for 6–8 hours. Videos were stored on hard-drives and transcribed back at the University of Montana after the field season. Incubation attentiveness was calculated as the amount of time spent on the nest divided by the number of daylight hours. Brooding attentiveness was calculated as the amount of time the adult spent brooding divided by the total time filmed that day. Provisioning rate was calculated as the number of feeding trips to the nest per hour. Data on food delivered to nestlings was recorded only when the food item was identifiable.
Adult capture
We conducted standard-effort mist netting, as well as target-netting at nests, to capture and color-band adults throughout the primary field site (Martin et al. 2015a, 2017). When adults were caught, they were fitted with an aluminum band and a unique combination of colored plastic bands for future identification. Standard morphometrics were also collected, including mass, tarsus length, and wing chord. We looked for evidence of breeding condition, such as brood patch or cloacal protuberance to determine sex when possible.
Statistical analyses
Incubation and nestling periods were only calculated for nests where the transition (clutch completion, hatching, fledging) was known within 24 h on one end and 24–48 h on the other end of the stage. Nest survival was calculated using only nests that were confirmed to be active by visual confirmation of adults at the nest and eggs or nestlings inside. All data were analyzed using program R, version 4.0.3 (R Core Team 2020). Daily nest survival was calculated using a logistic exposure model (Shaffer 2004). Daily nest predation was calculated based on nests that failed due to predation relative to exposure time of all nests.
Incubation attentiveness was assessed by fitting a polynomial mixed effects model with species as a fixed effect and individual nest ID as a random factor. For nestling growth data, we used measurements of nestling mass, tarsus, and wing chord to fit logistic growth curves and calculate the growth rate constant, K (Remeš and Martin 2002). Models were fit using package nlme (Pinheiro et al. 2022). Nest provisioning rates were assessed using a linear mixed model with species, nestling age, and brood size as fixed effects and individual nest ID as a random effect. Nest attentiveness during the nestling stage was assessed by fitting backward logistic curves to attentiveness data. We used a self-starting logistic model from the stats package (R Core Team 2020). All means are reported ± 1 standard error.
RESULTS
We found and monitored a total of 77 forktail nests and 130 whistling-thrush nests. Forktail nests were found at elevations ranging from 1406 to 1653 m asl, while most whistling-thrush nests were found from 1414 to 1699 m asl. Whistling-thrush also nested at higher elevations, and an additional nest was found at 3206 m asl. The two species overlapped extensively in timing of reproduction. The earliest initiation dates found by backdating were 10 January for forktails and 2 January for whistling-thrushes (Fig. 2). Median initiation dates were 22 March for forktails and 5 April for whistling-thrushes (Fig. 2). We observed both forktails and whistling-thrush re-nesting within the season. On six occasions, forktails were observed renesting after a successful first nest (i.e., double-brooding), four occasions after a failed nest, and one instance of two re-nests after failed attempts. For whistling-thrushes, we observed four occurrences of double-brooding and three re-nests after a failed nest.
Nest description
Both the whistling-thrush and forktail built large, open cup nests and were found primarily in riparian (creek/river) habitats (Fig. 1c, d). Whistling-thrush nests were much larger than forktail nests, which is unsurprising given the larger size of adult whistling-thrushes (Table 1).
Forktail nests were made of moss and mud and lined with leaf skeletons (Fig. 1c, e). Most of the nests were on banks (n = 39) or rocks (n = 21), while the rest were in trees (n = 9), logs (n = 3), snags (n = 2), or palms (n = 1, the remainder of nests had no substrate recorded). Forktails were not observed re-using nests in the same year, but they were observed building new nests on top of nests from previous years. Adults typically started nest building by bringing clumps of mud to the nest site. Next, they used moss to build the base and sides of the cup, and finally, leaf skeletons were used to line the cup. The longest building period we observed was 20 days, but very few nests were found in the earliest stages of building, so the building period could be even longer in some cases.
Whistling-thrush nests were large structures made of mud, sticks, dried leaves, and moss and lined with rootlets (Fig. 1d, e). Whistling-thrushes were more flexible in their nesting sites than forktails and the vast majority of nests were on rocks (n = 69) and human-made structures (n = 46), while a few nests were found on trees (n = 4) and banks (n = 3, the remainder of nests had no substrate recorded). Whistling-thrushes were frequently observed re-using nests from previous years. The nest building period was typically long, with many days of no activity between building bouts, even in nests that were re-used from previous years. Adults would rebuild the often-collapsed walls of the old nest and re-line the nest with rootlets (Fig. 1f).
Eggs and clutch size
Forktail eggs were light tan in color with brown spots somewhat concentrated on the larger end (Fig. 1e). Forktail eggs were much smaller than those of whistling-thrushes (Table 1) and averaged 11.0% of female mass. Whistling-thrush eggs were elliptical in shape and off-white with light brown spots concentrated near the larger end (Fig. 1f) and averaged 9.6% of female mass. Clutch size was very similar between the species (Table 1).
Incubation period
Only the female incubated in both species. Forktail incubation length was slightly shorter than in whistling-thrush (Table 1). Forktail attentiveness rose sharply for the first five days of the incubation period, then plateaued around 65% of daylight hours (n = 13 nests, 1728 h of nest probe data) with a slight drop near the end (Fig. 3). Similarly, whistling-thrush attentiveness increased for the first half of the incubation period, plateaued around day 7 at about 70% of daylight hours (Fig. 3, n = 11 nests, 2335 h of probe data), and then dropped off near the end of incubation. Incubation attentiveness did not differ significantly between the species (Fig. 3, P = 0.2; species term in the mixed effects model).
Nestling period
Forktail nestling period was significantly shorter than that of the whistling-thrush (Table 1). The eighth primary broke through the sheath on day 8 (n = 7), 9 (n = 12), or 10 (n = 3) for forktails and on day 8 (n = 1), 9 (n = 15), 10 (n = 26), or 11 (n = 1) for whistling-thrush. Forktail nestlings fledged at 89.3% of average adult mass and 67.8% of adult wing length, while whistling-thrush nestlings fledged at 85.8% of adult mass and 71.3% of adult wing length (Fig. 4, Table 1). Growth rate of forktail nestlings was faster than for whistling-thrushes, based on the growth rate constant, K, for mass, (t1289 = 5.88, P < 0.001, Fig. 4a), tarsus (t1289 = 6.43, P < 0.001, Fig. 4b), and wing chord (t1289 = 9.58, P < 0.001, Fig. 4c).
Males and females fed the nestlings in both species. Forktail adults fed nestlings 46% insects, 12% larva, 14% frogs, 4% lizards, and 24% worms. For the whistling-thrush, food items delivered included 1% seeds, 46% insects, 12% larva, 6% frogs, 3% snakes, 16% lizard, and 17% worms, along with eggs and nestlings of other songbirds. Forktails fed at higher rates than whistling-thrush (Fig. 5a; P < 0.001; species term in the mixed effects model) and feeding rates were more variable (range = 2.45–16.75 trips/h; n = 29 nests, 533 h of video) than for whistling-thrush (range = 0.47–8.07 trips/h; n = 58 nests, 1442 h of video). Feeding rate declined slightly with nestling age (P < 0.001; age term in the mixed effects model) and increased with brood size (P < 0.001; brood size term in mixed effects model).
Only females brooded the nestlings in both species. The percent of time spent on the nest decreased as nestlings aged for both forktails (Fig. 5b; n = 29 nests, 533 h of video) and whistling-thrush (Fig. 5b, n = 59 nests, 1466 h). Brooding rates decreased more gradually for whistling-thrush than for forktails (t277 = 3.10, P = 0.002), reaching zero after day 20 in many cases, while doing so around day 8 in forktails (Fig. 5b).
Nest survival and predation
Twenty-one of the 77 forktail nests monitored were successful, while 42 were depredated and 7 were still active when we left the study site. The remaining nests (n = 7) failed because of abandonment, external circumstances (such as weather and poor nest construction), and unknown causes. When split up by stage of the nesting cycle, daily predation was 0.049 ± 0.009 during incubation, and 0.034 ± 0.010 during the nestling stage and no predation was observed during lay. Predators observed on video included treeshrews (Scandentia) and the whistling-thrush. Other potential predators included Bornean Green-Magpies (Cissa jefferyi), Hair-crested Drongos (Dicrurus hottentottus), Malayan weasels (Mustela nudipes), and macaques (Macaca spp.).
Of the 130 whistling-thrush nests monitored, 58 were successful and 35 were predated. Seventeen were still active when we left the field site and the 20 remaining nests failed because of abandonment or other reasons. Whistling-thrush daily nest success was higher (Table 1, t205 = -4.24, P < 0.001) and daily nest predation was lower than for forktails (Table 1, t205 = -4.75, P < 0.001). Split up by nesting stage, whistling-thrush daily nest predation was 0.015 ± 0.003 during incubation, and 0.010 ± 0.003 during the nestling stage and no predation was observed during lay. No predators were observed on video, but the potential predators are similar to those of forktail nests. Whistling-thrush nest predation was higher on natural substrates (0.017 ± 0.003, n = 46) than on human-made structures (0.005 ± 0.002, n = 76, t120 = 3.34, P = 0.001).
Whistling-thrush are nest predators at this site and were observed through video monitoring to depredate nests of 10 different species, including the forktail. Other species that were depredated by the whistling thrush included Bornean Stubtail (Urosphena whiteheadi), Bornean Swiftlet (Collocalia dodgei), Chestnut-crested Yuhina (Staphida everetti), Mountain Wren-Babbler (Gypsophila crassa), Snowy-browed Flycatcher (Ficedula hyperythra), Temminck’s Babbler (Pellorneum pyrrogenys), White-browed Shortwing (Brachypteryx montana), White-throated Fantail (Rhipidura albicollis), and Yellow-breasted Warbler (Phylloscopus montis). Of the nests depredated by whistling-thrush (n = 16), roughly one third were depredated during incubation (n = 5) and the remainder were depredated during the nestling period.
DISCUSSION
Very little has been published on the breeding biology of the Enicurus and Myophonus genera, but the scant literature reveals interesting patterns across latitudes and between species. Both species we studied exhibited life history strategies typical of slow-paced, long-lived tropical species. However, both genera have members throughout temperate and tropical Asia, so some comparisons across latitudes are possible.
The two species we studied had very similar clutch sizes, while other subspecies of White-crowned Forktails at higher latitudes were found to have clutch sizes of 3–4 eggs (Wenkai 2013, Pierce et al. 2020), following the common pattern of increasing clutch size with latitude (Lack 1947, Jetz et al. 2008). The whistling-thrushes follow a similar pattern, where tropical species, such as the Malayan (M. robinsoni), Javan (M. glaucinus), and Bornean Whistling-Thrushes (M. borneensis) have clutch sizes of two, whereas sub-tropical and temperate species, such as the Malabar (M. horsfieldii) and Taiwan Whistling-Thrushes (M. insularis) have clutch sizes up to four (Delacour 1942, Collar 2020a).
In China, White-crowned Forktails had an egg size of 4.9 g at hatch (n = 9; Wenkai 2013), which is almost one gram larger than the mean egg size we report here. This is surprising, given that egg size can decrease with larger clutch size (Smith and Fretwell 1974, but see Martin 2008). Additional data on adult body mass, predation risk, and incubation attentiveness of forktails in more temperate locations may help explain this pattern (Martin et al. 2006, Martin 2008). No data have been published on whistling-thrush egg sizes, but based on their larger average body mass, they should have much larger eggs than the forktails (Sæther 1987, Martin et al. 2006). Differences in egg mass between species may also be influenced by demographic parameters such as clutch size, nest predation rates, and adult mortality.
At sites in Thailand (latitude ~14°N) and China (latitude ~26.6°N), other subspecies of White-crowned Forktails had incubation periods of 16 (n = 5) and 17–19 days (n = 6), respectively, and nestling periods of 15 (n = 8) and 16–17 days (n = 6), respectively (Wenkai 2013, Pierce et al. 2020). We found incubation and nestling period lengths in the same range as these studies, yielding no clear latitudinal difference. Minor differences between our results and other studies may be due to the different climates across the studies or to evolved differences between subspecies.
No data outside of our study have been published on the Bornean Whistling-Thrush, but some data is available for other species in the genus Myophonus. Malabar Whistling-Thrushes (range = 8–23°N) had incubation periods of 16–17 days, while Taiwan Whistling-Thrushes (range = 22–25°N) had incubation periods of 12–14 days (Collar 2020a). This very small set of data suggests that incubation periods may increase toward the tropics (Martin et al. 2007), as our tropical species had the longest period of the three species. All three species are roughly the same size (25–30 cm; Collar 2020a), so differences in body mass do not appear to be driving the trend. Instead, the trend may be the result of differences in adult mortality and incubation attentiveness between the species (Martin 2002, Martin et al. 2015a), but no data are available to test this hypothesis. Unfortunately, virtually nothing else has been published about the reproductive biology of any of the species in the genus Myophonus.
Forktail daily nest success in Thailand (0.975 ± 0.008, n = 22; Pierce et al. 2020), the only other study of nest success, was higher than at our site. Whistling-thrush nest success has not been reported outside of our study. However, we found that whistling-thrush nesting on human-made structures benefitted from significantly lower nest predation rates than those nesting on natural substrates. Thus, it appears that proximity to human settlements may deter some predators. Other species of whistling-thrush have also been found nesting on human-made structures (Delacour 1942, Fang and Wang 2002, Walther 2015), so predation reduction may be a widespread pattern.
While forktails and whistling-thrush had similar habitat associations, nesting seasons, and clutch sizes, they were different from each other in all other reproductive traits studied, including relative egg size, incubation attentiveness, nestling period, nestling growth rate, provisioning rates, and daily nest predation. These differences are potentially attributable to the fact that the whistling-thrush is a large, predatory species, and adults are better able to protect their nests. Unzeta et al. (2020) found that larger songbird species were able to deter a larger range of nest predators than smaller species. Whistling-thrushes are large enough to deter nest predation by small mammals and other birds, but forktails are not. As a result, whistling-thrush had lower daily nest predation rates, which were associated with longer nestling periods and slower nestling growth than forktails (Bosque and Bosque 1995, Martin 1995, Martin et al. 2011). Despite whistling-thrush’s longer nestling periods, nestlings of both species fledge with similar mass, tarsus, and wings relative to adults (Fig. 4). The more vulnerable forktails likely evolved faster nestling growth to permit nestlings to leave the nest sooner (Martin et al. 2011). However, short nestling periods and high nest predation rates are typically associated with lower provisioning rates, because of the risk of attracting the attention of predators with increased trips to the nest (Skutch 1949, Martin et al. 2000, 2011, Muchai and du Plessis 2005, Martin 2015, Matysioková and Remeš 2018). We found that forktails have high rates of provisioning, despite their high nest predation risk. Given that whistling-thrushes are generally larger and more predatory than forktails (Wang and Hung 2019, Collar 2020a, b), the dynamics seen here may be representative of broad life history differences between the two genera.
Although neither the whistling-thrush nor the forktail are classified as threatened species, climate change and habitat loss due to development are likely to cause population declines in the future. Climate change is expected to increase drought frequency (Dai 2012, Trenberth et al. 2014) and tropical riparian songbirds appear to suffer severe consequences in drought years (Martin and Mouton 2020). Both the whistling-thrush and the forktail reduced reproduction during a drought, and incurred declines in adult survival probability, indicating that frequent droughts may have long-term population consequences (Martin and Mouton 2020). Additionally, forest loss is accelerating at higher elevations in Southeast Asia (Zeng et al. 2018, Feng et al. 2021), threatening these species. Conserving intact primary forest and included riparian habitats, such as found in Kinabalu Park, is likely essential to the long-term survival of both species.
We uncovered some interesting patterns of divergences and similarities within and between the whistling-thrush and forktails despite their close taxonomic relatedness and living in the same habitat. The larger size and more predatory nature of the whistling-thrush sets it apart from most passerines and their ability to defend against nest predators has led to some unusual demographic traits. However, much remains to be learned about members of Enicurus and Myophonus. More information on the other members of these genera will provide an interesting contrast to the data presented here. Both genera are widespread across Asia and occur in temperate, sub-tropical, and tropical habitats, so latitudinal patterns may exist. Additional study will enhance our understanding of geographical patterns of life history variation.
RESPONSES TO THIS ARTICLE
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a response, follow this link. To read responses already accepted, follow this link.ACKNOWLEDGMENTS
Timothy Forrester, Holly Jackson, Adam Mitchell, and two anonymous reviewers provided helpful comments on the manuscript. We thank the many field assistants who found and monitored nests and the undergraduates who provided analysis of parental behavior data from nest videos. We greatly appreciate Sabah Parks and the Sabah Biodiversity Council for authorization and collaboration on this project. Support for this work came from the National Science Foundation (DEB-1241041, DEB-1651283, IOS-1656120, IOS-1656273), and was conducted under University of Montana IACUC #059-10TMMCWRU.
DATA AVAILABILITY
The data/code that support the findings of this study will be openly available in Dryad upon publication. Ethical approval for this research study was granted by University of Montana IACUC #059-10TMMCWRU.
LITERATURE CITED
BirdLife International. 2016a. Myophonus borneensis. The IUCN Red List of Threatened Species 2016:e.T22732973A95053418. https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22732973A95053418.en
BirdLife International. 2016b. Enicurus leschenaulti. The IUCN Red List of Threatened Species 2016:e.T22710138A94236268. https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22710138A94236268.en
Blake, J. G., and B. A. Loiselle. 2008. Estimates of apparent survival rates for forest birds in eastern Ecuador. Biotropica 40:485-493. https://doi.org/10.1111/j.1744-7429.2007.00395.x
Bosque, C., and M. T. Bosque. 1995. Nest predation as a selective factor in the evolution of developmental rates in altricial birds. American Naturalist 145:234-260. https://doi.org/10.1086/285738
Boyle, W. A., B. K. Sandercock, and K. Martin. 2016. Patterns and drivers of intraspecific variation in avian life history along elevational gradients: a meta-analysis. Biological Reviews 91:469-482. https://doi.org/10.1111/brv.12180
Collar, N. 2020a. Genus Myophonus, version 1.0. In Birds of the World, Cornell Lab of Ornithology, Ithaca, New York, USA.
Collar, N. 2020b. Genus Enicurus, version 1.0. In Birds of the World, Cornell Lab of Ornithology, Ithaca, New York, USA.
Dai, A. 2012. Increasing drought under global warming in observations and models. Nature Climate Change 3:52-58. https://doi.org/10.1038/nclimate1633
Delacour, J. 1942. The Whistling Thrushes (Genus Myiophoneus). Auk 59:246-264. https://doi.org/10.2307/4079555
Fang, J. R., and Y. Wang. 2002. Environmental characteristics of bridges selected by nesting Formosan Whistling Thrush (Myiophoneus insularis). Bioformosa 37:17-23.
Feng, Y., A. D. Ziegler, P. R. Elsen, Y. Liu, X. He, D. V. Spracklen, J. Holden, X. Jiang, C. Zheng, and Z. Zeng. 2021. Upward expansion and acceleration of forest clearance in the mountains of southeast Asia. Nature Sustainability 4:892-899. https://doi.org/10.1038/s41893-021-00738-y
Fennici, A. Z. 1984. Breeding biology of the Pied Flycatcher Ficedula hypoleuca in relation to population density. Annales Zoologici Fennici 21:187-197.
Jetz, W., C. H. Sekercioglu, and K. Böhning-Gaese. 2008. The worldwide variation in avian clutch size across species and space. PLoS Biology 6(12):e303. https://doi.org/10.1371/journal.pbio.0060303
K. Lisa Yang Center for Conservation Bioacoustics. 2016. Raven Lite: Interactive Sound Analysis Software. Cornell Lab of Ornithology, Ithaca, New York, USA. https://www.birds.cornell.edu/ccb/
Karr, J. R., J. D. Nichols, M. K. Klimkiewicz, and J. D. Brawn. 1990. Survival rates of birds of tropical and temperate forests: Will the dogma survive? American Naturalist 136:277-291. https://doi.org/10.1086/285098
Kitayama, K., and S. I. Aiba. 2002. Ecosystem structure and productivity of tropical rain forests along altitudinal gradients with contrasting soil phosphorus pools on Mount Kinabalu, Borneo. Journal of Ecology 90:37-51. https://doi.org/10.1046/j.0022-0477.2001.00634.x
Lack, D. 1947. The significance of clutch-size. Parts I and II. Ibis 89(2):302-352. https://doi.org/10.1111/j.1474-919X.1947.tb04155.x
Lu, X., D. Ke, Y. Guo, S. Tang, L. Zhang and C. Wang. 2011. Breeding ecology of the Black Redstart Phoenicurus ochruros at a Tibetan site, with special reference to cooperative breeding. Ardea 99:235-240. https://doi.org/10.5253/078.099.0215
MacArthur, R. H., and E. O. Wilson. 1967. The theory of island biogeography. Princeton University Press, Princeton, New Jersey, USA. https://doi.org/10.1515/9781400881376
Martin, T. E. 1995. Avian life history evolution in relation to nest sites, nest predation, and food. Ecological Monographs 65:101-127. https://doi.org/10.2307/2937160
Martin, T. E. 1996. Life history evolution in tropical and south temperate birds: What do we really know? Journal of Avian Biology 27:263-272. https://doi.org/10.2307/3677257
Martin, T. E. 2002. A new view of avian life-history evolution tested on an incubation paradox. Proceedings of the Royal Society B: Biological Sciences 269:309-316. https://doi.org/10.1098/rspb.2001.1879
Martin, T. E. 2008. Egg size variation among tropical and temperate songbirds: an embryonic temperature hypothesis. Proceedings of the National Academy of Sciences of the United States of America 105:9268-9271. https://doi.org/10.1073/pnas.0709366105
Martin, T. E. 2015. Age-related mortality explains life history strategies of tropical and temperate songbirds. Science 349:966-970. https://doi.org/10.1126/science.aad1173
Martin, T. E., S. K. Auer, R. D. Bassar, A. M. Niklison, and P. Lloyd. 2007. Geographic variation in avian incubation periods and parental influences on embryonic temperature. Evolution 61:2558-2569. https://doi.org/10.1111/j.1558-5646.2007.00204.x
Martin, T. E., R. D. Bassar, S. K. Bassar, J. J. Fontaine, P. Lloyd, H. A. Mathewson, A. M. Niklison, and A. Chalfoun. 2006. Life-history and ecological correlates of geographic variation in egg and clutch mass among passerine species. Evolution 60:390-398. https://doi.org/10.1111/j.0014-3820.2006.tb01115.x
Martin, T. E., and G. R. Geupel. 1993. Nest-monitoring plots: methods for locating nests and monitoring success. Journal of Field Ornithology 64:507-519.
Martin, T. E., P. Lloyd, C. Bosque, D. C. Barton, A. L. Biancucci, Y. R. Cheng, and R. Ton. 2011. Growth rate variation among passerine species in tropical and temperate sites: an antagonistic interaction between parental food provisioning and nest predation risk. Evolution 65:1607-1622. https://doi.org/10.1111/j.1558-5646.2011.01227.x
Martin, T. E., and J. C. Mouton. 2020. Longer-lived tropical songbirds reduce breeding activity as they buffer impacts of drought. Nature Climate Change 10:953-958. https://doi.org/10.1038/s41558-020-0864-3
Martin, T. E., J. C. Oteyza, A. J. Boyce, P. Lloyd, and R. Ton. 2015a. Adult mortality probability and nest predation rates explain parental effort in warming eggs with consequences for embryonic development time. American Naturalist 186:223-236. https://doi.org/10.1086/681986
Martin, T. E., J. C. Oteyza, A. E. Mitchell, A. L. Potticary, and P. Lloyd. 2015b. Postnatal growth rates covary weakly with embryonic development rates and do not explain adult mortality probability among songbirds on four continents. American Naturalist 185:380-389. https://doi.org/10.1086/679612
Martin, T. E., M. M. Riordan, R. Repin, J. C. Mouton, and W. M. Blake. 2017. Apparent annual survival estimates of tropical songbirds better reflect life history variation when based on intensive field methods. Global Ecology and Biogeography 26:1386-1397. https://doi.org/10.1111/geb.12661
Martin, T. E., J. Scott, and C. Menge. 2000. Nest predation increases with parental activity: separating nest site and parental activity effects. Proceedings of the Royal Society B: Biological Sciences 267:2287-2293. https://doi.org/10.1098/rspb.2000.1281
Matysioková, B., and V. Remeš. 2018. Evolution of parental activity at the nest is shaped by the risk of nest predation and ambient temperature across bird species. Evolution 72:2214-2224. https://doi.org/10.1111/evo.13580
Mitchell, A. E., J. Boersma, A. Anthony, K. Kitayama, and T. E. Martin. 2020. Experimental amelioration of harsh weather speeds growth and development in a tropical montane songbird. American Naturalist 196:E110-E118. https://doi.org/10.1086/710151
Morgan, R. 1982. The breeding biology of the Nightingale Luscinia megarhynchos in Britain. Bird Study 29:67-72. https://doi.org/10.1080/00063658209476739
Moyle, R. G., J. D. Manthey, P. A. Hosner, M. Rahman, M. Lakim, and F. H. Sheldon. 2017. A genome-wide assessment of stages of elevational parapatry in Bornean passerine birds reveals no introgression: implications for processes and patterns of speciation. PeerJ 5:e3335. https://doi.org/10.7717/peerj.3335
Moyle, R. G., M. Schilthuizen, M. A. Rahman, and F. H. Sheldon. 2005. Molecular phylogenetic analysis of the White-crowned Forktail Enicurus leschenaulti in Borneo. Journal of Avian Biology 36:96-101. https://doi.org/10.1111/j.0908-8857.2005.03510.x
Muchai, M., and M. A. du Plessis. 2005. Nest predation of grassland bird species increases with parental activity at the nest. Journal of Avian Biology 36:110-116. https://doi.org/10.1111/j.0908-8857.2005.03312.x
Pierce, A. J., W. Sankamethawee, L. A. Powell, and G. A. Gale. 2020. Patterns of nesting and nest success in an evergreen forest in Southeast Asia. Emu 120:46-55. https://doi.org/10.1080/01584197.2019.1686645
Pinheiro, J., D. Bates, and R Core Team. 2022. nlme: Linear and nonlinear mixed effects models.
Porkert, J., and J. Zajíc. 2005. The breeding biology of the Common Redstart, Phoenicurus phoenicurus, in the Central European pine forest. Folia Zoologica 54:111-122.
R Core Team. 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org
Remeš, V., and T. E. Martin. 2002. Environmental influences on the evolution of growth and developmental rates in passerines. Evolution 56:2505-2518. https://doi.org/10.1111/j.0014-3820.2002.tb00175.x
Sæther, B.-E. 1987. The influence of body weight on the covariation between reproductive traits in European birds. Oikos 48:79-88. https://doi.org/10.2307/3565691
Sangster, G., P. Alström, E. Forsmark, and U. Olsson. 2010. Multi-locus phylogenetic analysis of Old World chats and flycatchers reveals extensive paraphyly at family, subfamily and genus level (Aves: Muscicapidae). Molecular Phylogenetics and Evolution 57:380-392. https://doi.org/10.1016/j.ympev.2010.07.008
Shaffer, T. L. 2004. A unified approach to analyzing nest success. Auk 121:526-540. https://doi.org/10.1093/auk/121.2.526
Sheldon, F. H., H. C. Lim, J. Nais, M. Lakim, A. Tuuga, P. Malim, J. Majuakim, A. Lo, M. Schilthuizen, P. A. Hosner, and R. G. Moyle. 2009. Observations on the ecology, distribution and biogeography of forest birds in Sabah, Malaysia. Raffles Bulletin of Zoology 57:577-586.
Skutch, A. F. 1949. Do tropical birds rear as many young as they can nourish? Ibis 91:430-455. https://doi.org/10.1111/j.1474-919X.1949.tb02293.x
Smith, C. C., and S. D. Fretwell. 1974. The optimal balance between size and number of offspring. American Naturalist 108:499-506. https://doi.org/10.1086/282929
Trenberth, K. E., A. Dai, G. Van Der Schrier, P. D. Jones, J. Barichivich, K. R. Briffa, and J. Sheffield. 2014. Global warming and changes in drought. Nature Climate Change 4:17-22. https://doi.org/10.1038/nclimate2067
Unzeta, M., T. E. Martin, and D. Sol. 2020. Daily nest predation rates decrease with body size in passerine birds. American Naturalist 196:743-754. https://doi.org/10.1086/711413
Walther, B. A. 2015. Two unusual nest-sites of the Taiwan Whistling Thrush Myophonus insularis: part of a trend towards urbanisation? BirdingASIA 24:122-125.
Wang, J. S., and C. M. Hung. 2019. Barn swallow nest predation by a recent urban invader, the Taiwan whistling thrush - implications for the evolution of urban avian communities. Zoological Studies 58:e1.
Wenkai, C. 2013. Breeding ecology of Enicurus leschenaulti. Sichuan Journal of Zoology 4.
Zeng, Z., L. Estes, A. D. Ziegler, A. Chen, T. Searchinger, F. Hua, K. Guan, A. Jintrawet, and E. F. Wood . 2018. Highland cropland expansion and forest loss in Southeast Asia in the twenty-first century. Nature Geoscience 11:556-562. https://doi.org/10.1038/s41561-018-0166-9
Zuccon, D., and P. G. P. Ericson. 2010. A multi-gene phylogeny disentangles the Chat-flycatcher complex (Aves: Muscicapidae). Zoologica Scripta 39:213-224. https://doi.org/10.1111/j.1463-6409.2010.00423.x

Fig. 1

Fig. 1. Images of Bornean Forktail (Enicurus leschenaulti borneensis) and Bornean Whistling-Thrush (Myophonus borneensis) adults, nests, eggs, and nestlings. (A) Bornean Forktail adults at nest, (B) Bornean Whistling-Thrush adult with nesting material, (C) Bornean Forktail nest with nestlings in the end of a hollow log, (D) Bornean Whistling-Thrush nest with eggs on a rock ledge, (E) Bornean Forktail egg and newly hatched nestling, and (F) Bornean Whistling-Thrush egg and newly hatched nestling. Photos by Thomas Martin, Warren Whaley, Elise Zarri, and Daniel Muñoz.
Fig. 2
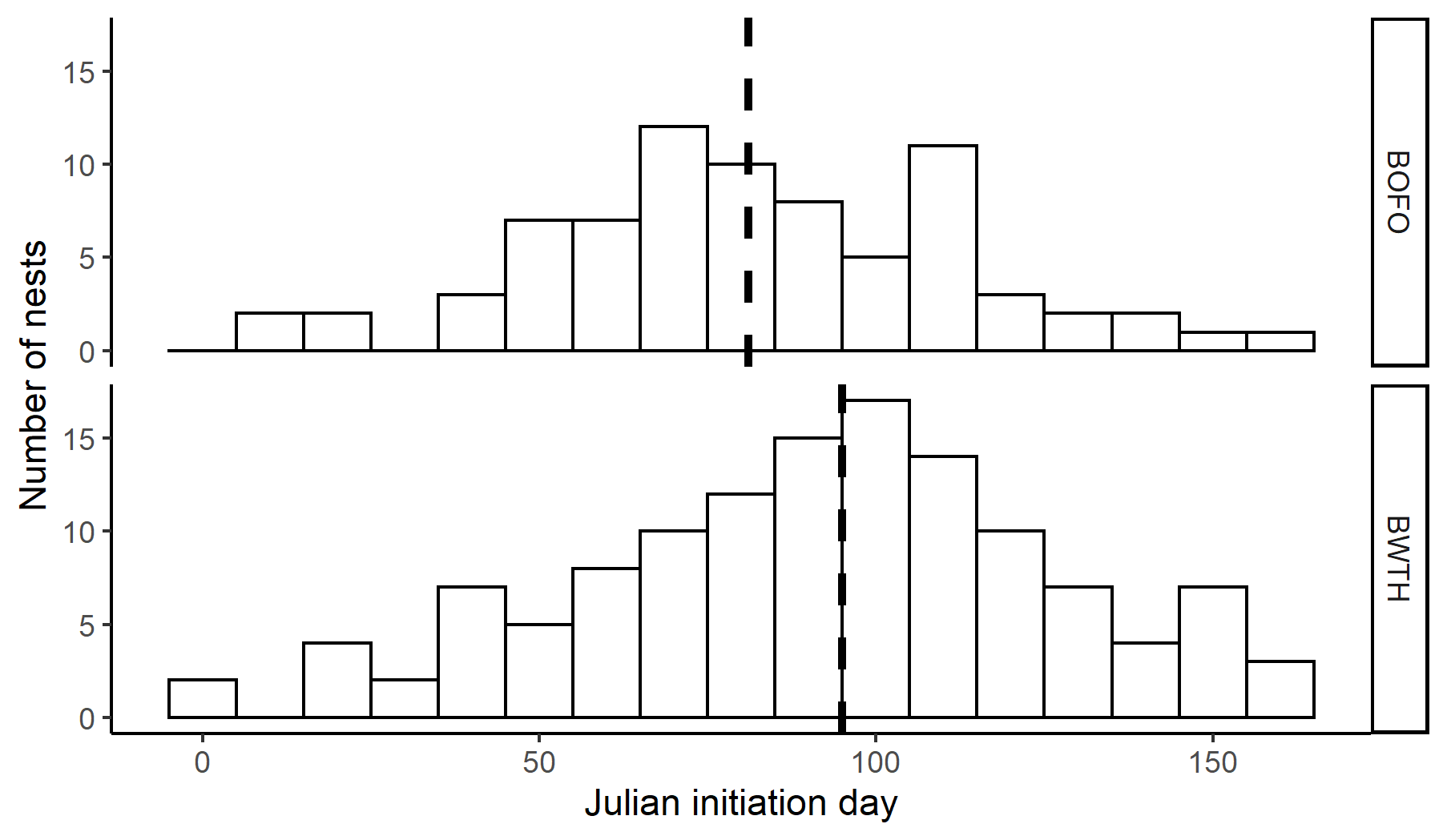
Fig. 2. Timing of nest initiation for the Bornean Forktail (Enicurus leschenaulti borneensis, BOFO, top) and the Bornean Whistling-Thrush (Myophonus borneensis, BWTH, bottom) from 2009 to 2020 in Kinabalu Park, Sabah, Malaysia. Dashed line indicates median initiation date.
Fig. 3
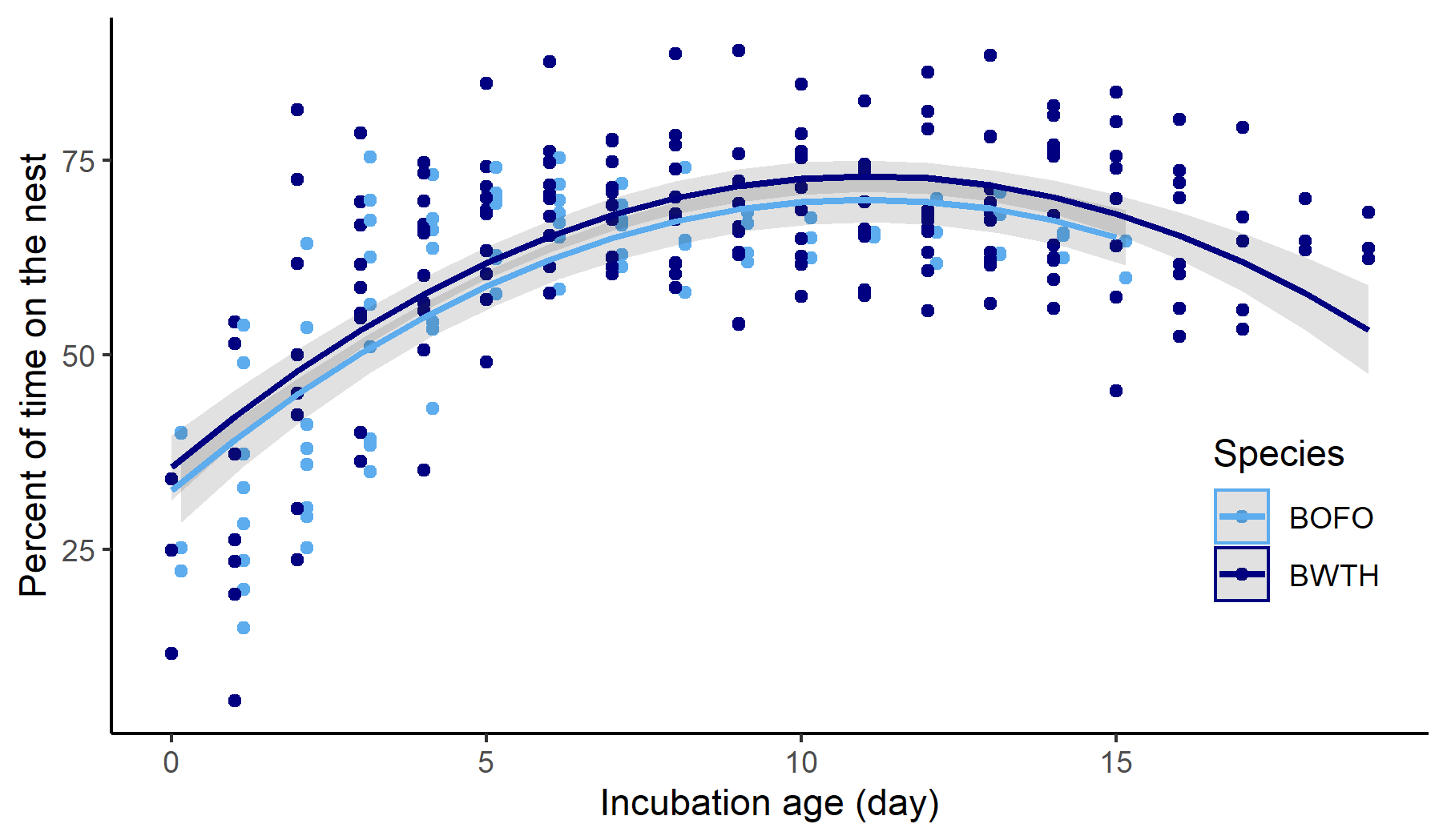
Fig. 3. Incubation attentiveness for the Bornean Forktail (Enicurus leschenaulti borneensis, BOFO, light blue) and the Bornean Whistling-Thrush (Myophonus borneensis, BWTH, dark blue). Each point represents the percent time on the nest for one nest during one day of probe data (daylight hours only). Gray bands represent the 95% confidence intervals.
Fig. 4
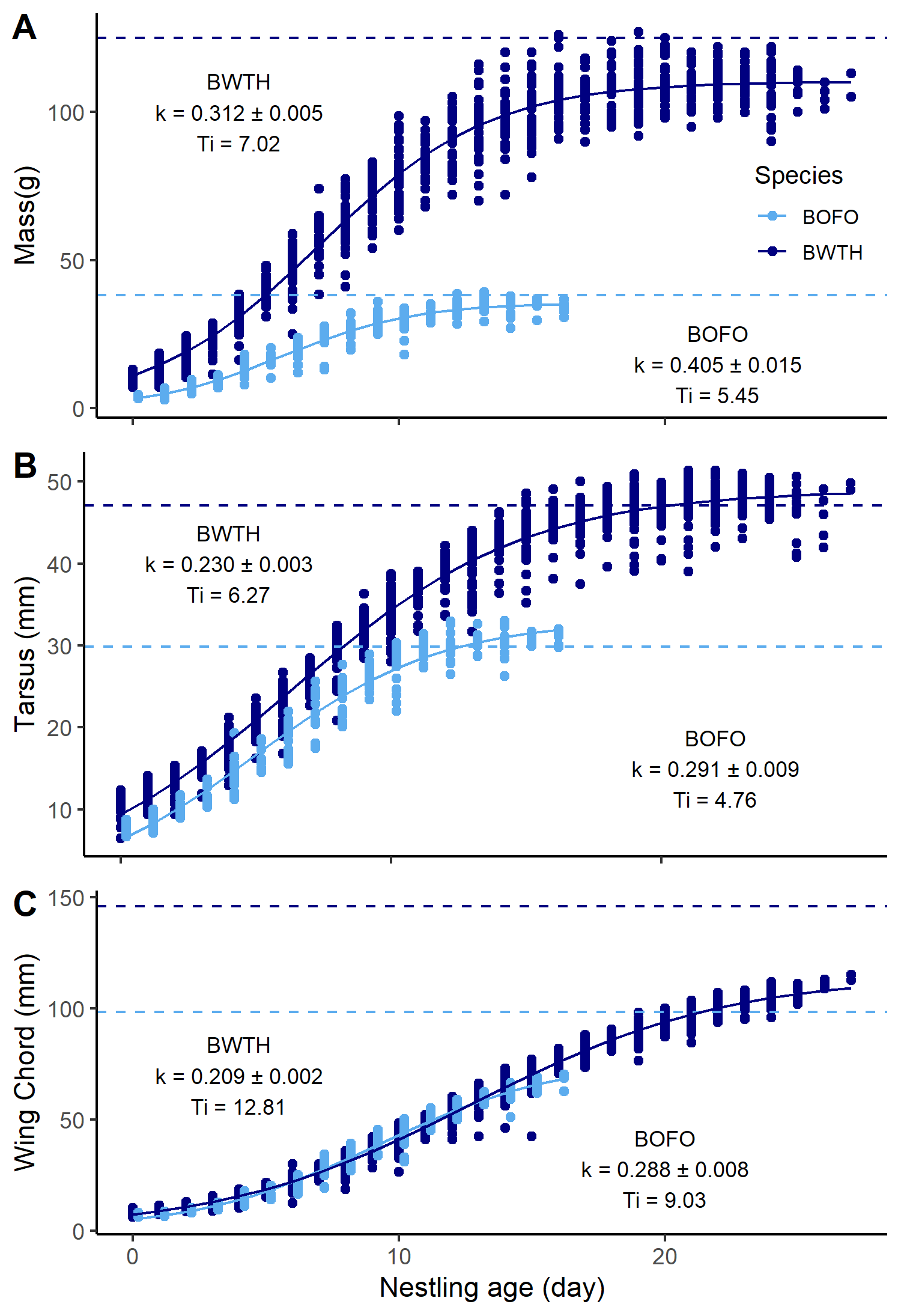
Fig. 4. Nestling growth rates for (A) mass, (B) tarsus, and (C) wing chord of Bornean Forktails (Enicurus leschenaulti borneensis, BOFO, light blue) and Bornean Whistling-Thrush (Myophonus borneensis, BWTH, dark blue). Dashed lines indicate adult mass, tarsus, and wing chords, respectively.
Fig. 5
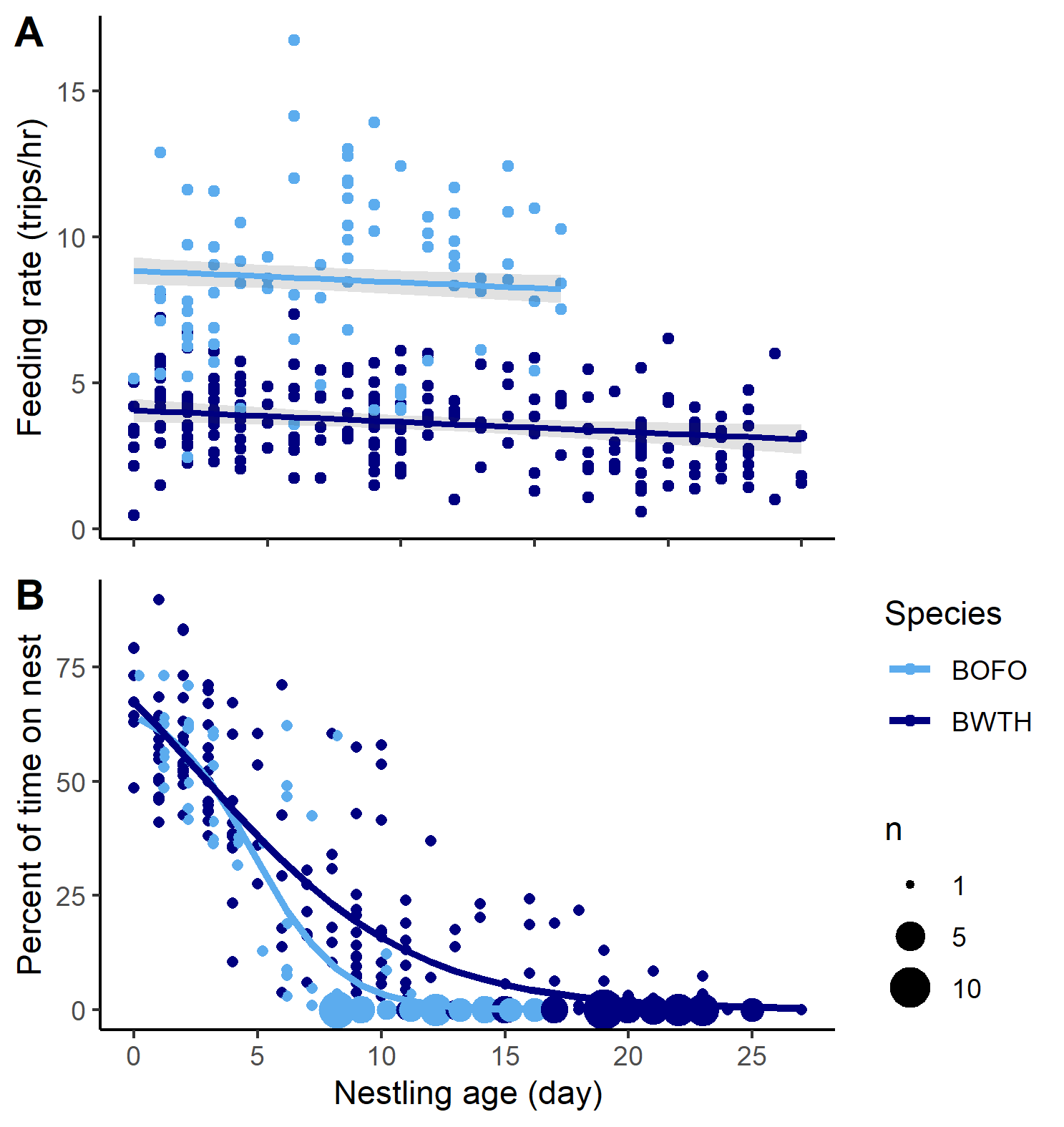
Fig. 5. (A) Nestling provisioning rate and (B) brooding attentiveness for Bornean Forktails (Enicurus leschenaulti borneensis, BOFO, light blue) and Bornean Whistling-Thrush (Myophonus borneensis, BWTH, dark blue) across the nestling period. Each point represents the feeding rate or percent time on the nest for a nest from one video recording period. Larger points on the brooding attentiveness plot represent multiple observations of 0.
Table 1
Table 1. Comparison of nest measurements and life history traits for Bornean Forktail (Enicurus leschenaulti borneensis) and Bornean Whistling-Thrush (Myophonus borneensis). Outer height of the nest refers to the distance between the lowest and highest part of the nest. Outer diameter refers to the distance between the outer edges of the nest. Cup height and diameter refer to the depth and inner diameter of the nest, respectively.
| Bornean Forktail | Bornean Whistling-Thrush | |||||
| Measurement | n | Mean ± SE | Range | n | Mean ± SE | Range |
| Dry mass | 6 | 61.9 ± 6.01 g | 46.6–80.2 g | 2 | 203.4 ± 66.00 g | 137.4–269.4 g |
| Outer height | 35 | 117.6 ± 5.08 mm | 62–230 mm | 24 | 145.1 ± 12.24 mm | 70–342 mm |
| Outer diameter | 35 | 136.1 ± 4.10 mm | 77–205 mm | 24 | 196.5 ± 12.25 mm | 120–380 mm |
| Cup height | 35 | 54.8 ± 2.17 mm | 41–112 mm | 24 | 59.3 ± 5.54 mm | 40–180 mm |
| Cup diameter | 35 | 74.7 ± 1.15 mm | 52–88 mm | 24 | 101.6 ± 2.34 mm | 80–120 mm |
| Male mass | 34 | 39.8 ± 0.51 g | 34.0–48.9 g | 34 | 127.5 ± 1.79 g | 94–140 g |
| Female mass | 28 | 36.9 ± 0.45 g | 32.5–41.8 g | 21 | 121.0 ± 2.27 g | 108–142 g |
| Clutch size | 37 | 2.0 ± 0.04 eggs | 1–3 eggs | 53 | 1.9 ± 0.04 eggs | 1–2 eggs |
| Egg mass | 78 | 4.1 ± 0.04 g | 3.7–4.6 g | 101 | 11.6 ± 0.12 g | 9.9–13.2 g |
| Incubation period | 8 | 17.0 ± 0.42 days | 16–19 days | 28 | 18.6 ± 0.19 days | 17–21 days |
| Nestling period | 9 | 15.9 ± 0.39 days | 14–18 days | 38 | 24.0 ± 0.20 days | 22–27 days |
| Nestling mass at fledge | 6 | 34.4 ± 0.85 g | 33.2–36.8 g | 29 | 107.3 ± 1.79 g | 80–130 g |
| Daily nest success | 77 | 0.927 ± 0.12 | - | 130 | 0.978 ± 0.003 | - |
| Daily nest predation | 77 | 0.042 ± 0.006 | - | 130 | 0.012 ± 0.002 | - |